
Abstract
Sirtuin 4 (SIRT4) belongs to the mitochondrial sirtuin protein family, a class of NAD+-dependent protein deacylases that remove post-translational acyl modifications from cellular substrates during the regulation of various biological pathways. SIRT4 has been shown to regulate lipid homeostasis. However, the mechanism by which the bovine SIRT4 gene is transcriptionally regulated remains unknown. To explore the molecular mechanism of SIRT4 expression, we obtained a 400-kb fragment of the 5′-regulatory region of bovine SIRT4 by molecular cloning, which contained a CpG island. Electrophoretic mobility shift assays and luciferase reporter gene assays identified the nuclear respiratory factor 1 (NRF1) and myb proto-oncogene protein (CMYB) binding sites as transcriptional repression and activation sites in the SIRT4 promoter region, respectively. We further verified that NRF1 and CMYB bind to the SIRT4 promoter using chromatin immunoprecipitation assays. In addition, from DNA methylation and reporter gene assays, results revealed that SIRT4 promoter activity was enhanced by demethylation. Further, NRF1-mediated transcriptional inhibition and CMYB-mediated transcriptional activation of SIRT4 expression were strengthened by demethylation during bovine adipocyte differentiation. Taken together, our results shed light on the mechanism underlying the promoter methylation and transcriptional regulation of SIRT4 expression in bovine adipocytes.
Introduction
S
SIRT4 is a mitochondrial ADP-ribosyltransferase (Zhao et al., 2004). SIRT4 downregulates insulin secretion in response to amino acids by inhibiting mitochondrial glutamate dehydrogenase 1 activity (Haigis et al., 2006). It has been shown that SIRT4 regulates fatty acid oxidation and mitochondrial gene expression in the liver, adipocytes, and muscle cells (Nasrin et al., 2010). SIRT4 is capable of removing several acyl-groups from lysine residues, although it was originally reported to be an ADP-ribosyltransferase located in the mitochondrial matrix. Previous studies have reported that it affects important biological pathways, especially those involved in metabolic diseases, such as diabetes and obesity (Kida and Goligorsky, 2016; Kumar and Lombard, 2017; Zaganjor et al., 2017; Tao et al., 2018). SIRT4 is highly expressed in islet β cells and can interact with adenine nucleotide translocator 2/3 and insulin-degrading enzyme (Ahuja et al., 2007). It also acts as a key factor of adipogenesis, regulating the proliferation and differentiation of preadipocytes (Ho et al., 2013; Laurent et al., 2013a, 2013b). In addition, it has been reported to decrease fatty acid oxidation and enhance lipid synthesis (Anderson et al., 2017; Carrico et al., 2018). However, the molecular mechanisms associated with bovine SIRT4 expression during adipose development are still unclear.
Transcriptional regulation is a vital physiological regulatory process in all living organisms. It is orchestrated by the coordination of transcription factors (TFs) and regulatory proteins. Therefore, the amount of RNA can be precisely tuned through various mechanisms, such as altering the transcribed RNA copy number and the temporal control and timing of gene transcription (Jacob and Monod, 1961). Transcriptional regulation allows the cell or organism to respond to a variety of intracellular and extracellular signals, and thus adapt accordingly. The promoter is a DNA fragment region, which may bind RNA polymerase, TFs, and other proteins for successful initiation of transcription directly upstream of the particular gene (Busby and Ebright, 1994). In general, the promoter is located near the transcription start site (TSS) of a gene and upstream on the DNA toward the 5′ region of the sense strand (Struhl, 1999). TFs can be divided in two main categories: activators and repressors (Wei et al., 2017b). The regulation of sequence-specific TFs, such as nuclear respiratory factor 1 (NRF1) and myb proto-oncogene protein (CMYB), is coordinated in part by the action of adipocytes.
DNA methylation is a process of adding methyl groups to a DNA molecule. The activity of the DNA segment can be changed through methylation without changing the sequence. DNA methylation of gene promoters plays a key role in the regulation of expression (Cao et al., 2018b). Remarkably, there is a link between TF binding and DNA methylation status (Wang et al., 2017a). When DNA methylation is located in a gene promoter, it can typically inhibit gene transcription by repressing TF binding to the promoter. However, the relationship between methylation and transcriptional regulation of the SIRT4 promoter activity during adipocyte differentiation still needs to be clarified.
This study aimed to explore the transcriptional regulatory mechanism of SIRT4 in bovine adipose tissue. Further, this study aimed to evaluate the effect of DNA methylation on transcriptional regulation via the 5′-regulatory region during adipocyte differentiation.
Materials and Methods
Ethics statement
All the experimental procedures with cattle used in this study had been approved by the Experimental Animal Manage Committee of Northwest A&F University (2011—31,101,684). Cattle were raised with free access to food and humanely slaughtered in the National Beef Cattle Improvement Center (Yangling, China). All the operations and experimental procedures were complied with the National Standard of Laboratory Animal-Guideline for Ethical Review of Animal Welfare (GB/T 35892-2018) (General Administration of Quality Supervision, Inspection and Quarantine of the People's Republic of China, Standardization Administration of the People's Republic of China, 2018) and Guide for the Care and Use of Laboratory Animals: Eighth Edition (National Research Council, 2011).
Quantitative PCR analysis of gene expression
Total RNA and cDNA were extracted and generated from bovine adipocytes using a Total RNA kit (Tiangen, Beijing, China) and PrimeScript™ RT Reagent kit with gDNA Eraser (Perfect Real Time; TaKaRa, Dalian, China), respectively, and the procedure was performed in accordance with the manufacturer's instructions. Quantitative PCR (qPCR) was performed using a SYBR Green PCR Master Mix kit (TaKaRa) and the 7500 System SDS V 1.4.0 (Applied Biosystems). The PCR reaction mixture was 20 μL, and the PCR conditions consisted of an initial incubation of 5 min at 95°C, followed by 34 cycles of 30 s at 95°C, 30 s at 60°C, and 30 s at 72°C. Gene-specific primers used for qPCR are listed in Table 1; β-actin was used as the endogenous control gene. The expression of genes was analyzed using the 2−ΔΔCt method (Livak and Schmittgen, 2001).
The restriction enzyme sites (Kpn I and Xho I) are indicated by the italicized letters in the forward (F) and reverse (R) primers used for promoter cloning. Underlines indicate base mutations.
EMSA, electrophoretic mobility shift assay; ChIP, chromatin immunoprecipitation; NRF1, nuclear respiratory factor 1; CMYB, myb proto-oncogene protein; SIRT4, sirtuin 4; DNMT, DNA methyltransferase.
Promoter cloning and generation of luciferase reporter vectors
Gene-specific primers (SIRT4-F/R, Table 1) were designed to amplify a 400-base pair (bp) promoter region upstream of the TSS of this gene (NCBI accession AC_000174.1 from 64873159 to 64875279). PCR amplifications were performed using genomic DNA from Qinchuan cattle blood as a template and KOD DNA Polymerase (Toyobo, Osaka, Japan). To generate the luciferase reporter vector, the 400-bp bovine SIRT4 promoter fragment was ligated into the pGL3-basic vector, which was digested with restriction enzymes KpnI and XhoI (TaKaRa) and named pGL3−402/+44. Potential TF binding sites were analyzed using the Genomatix suite (
Cell isolation, culture, and transfection
Bovine adipocytes were isolated from inguinal adipose tissue, which was collected from Qinchuan fetal cattle 3 days after birth. The inguinal adipose tissue was washed thrice in 1 × phosphate buffered saline (PBS), minced, and digested with 1 mg/mL collagenase type I (Invitrogen) at 37°C for 1.5 h, followed by filtration through a 200 μm nylon mesh. The cells were collected by centrifugation at 1500 g for 10 min and washed thrice with Dulbecco's modified Eagle medium:nutrient mixture F-12 (DMEM/F-12; Gibco). The cells were suspended in DMEM/F12 medium supplemented with 10% fetal bovine serum (FBS; PAN-Biotech, South America), 100 IU/mL penicillin, and 100 mg/mL streptomycin. Subsequently, the cells were seeded into 100 mm × 20 mm cell culture dishes with 5 × 104 cells/cm2 and cultured at 37°C in a humidified atmosphere with 5% CO2. After 12 h, cells were washed thrice with PBS to purify and cultured in fresh DMEM/F12 medium supplemented with 10% FBS, 100 IU/mL penicillin, and 100 μg/mL streptomycin. Under the microscope small lipid droplets were observed in the cells. To identify cell types, the medium of some cells was exchanged for DMEM/F12 supplemented with 10% FBS, 0.5 mM 3-isobutyl-1-methylxanthine (IBMX), 1 μM dexamethasone (DEX), and 5 μg/mL insulin to induce differentiation for 2 days. We reexamined the cells under the microscope and found that both the number and the diameter of the lipid droplets all increased, which confirmed that the isolated cells were undifferentiated bovine adipocytes.
3T3-L1 and bovine adipocytes were maintained in DMEM/F-12 supplemented with 10% FBS and antibiotics (100 IU/mL penicillin and 100 μg/mL streptomycin) at 37°C with 100% humidity and 5% CO2, and they were passaged using standard cell culture techniques. Cells were grown overnight in 24-well plates until reaching a confluency of 80–90%. The transfection mixture for each well comprised of the following: 800 ng of expression vectors, 10 ng of pRL-TK normalizing vector, 2 μL of X-tremeGENEHP DNA transfection reagent (Roche), and 100 μL of Opti-DMEM (GIBCO; Invitrogen). The pGL3-basic vector served as a negative control. Six hours after transfection, media were replaced with DMEM/F-12 containing 0.5 mM IBMX, 1 μM DEX, and 5 μg/mL insulin and incubated for 42 h to induce differentiation of 3T3-L1 and bovine adipocytes. Cell lysates were collected 48 h post-transfection and used to measure the relative transcriptional activity of each fragment using the Dual-Luciferase Reporter Assay System (Promega), according to the manufacturer's instructions. Relative luciferase activities were determined using a NanoQuant Plate™ (TECAN, infinite M200PRO). Experiments were conducted in parallel and in triplicate.
Site-directed mutagenesis and knockdown of TFs
The potential TF binding sites for NRF1 and CMYB were mutated using the corresponding primers (Table 1) and the QuickChange Site-Directed Mutagenesis Kit (Stratagene, La Jolla, CA). The procedure was run in accordance with the manufacturer's instructions. PCR conditions were in accordance with kit instructions, and products were treated with DpnI and then amplified in XL10-Gold competent cells (Stratagene). The resulting vectors were sequenced for confirmation. siRNAs targeting NRF1 and CMYB (Table 1) were designed and synthesized along with the control siRNA (GenePharma Co., Ltd., Shanghai, China); negative control (NC) siRNA served as a negative control. Bovine adipocytes cultured in 24-well plates were transiently co-transfected with 50 nM of each siRNA and the corresponding pGL3−402/+44 vector.
Electrophoretic mobility shift assays
Nuclear extracts from bovine adipocytes were prepared using the Nuclear Extract Kit (Active Motif Corp., Carlsbad, CA) according to the manufacturer's protocol (Kamanga-Sollo et al., 2010). A LightShift Chemiluminescent EMSA Kit (Thermo Fisher Corp., Waltham, MA) was used for electrophoretic mobility shift assay (EMSA) in accordance with the manufacturer's protocol with modifications. All DNA probes (listed in Table 1) were synthesized (Invitrogen) and labeled at the 5′ end with biotin. Briefly, 10 μg of nuclear protein extract was incubated with 2 μL of 10 × binding buffer and 1 μL of poly (dI.dC) in a 20-μL volume for 15 min on ice. Subsequently, 200 fmol of 5′-biotin labeled probes were added, and the reaction mixture was incubated at room temperature (26°C) for 20 min. For the competition assay, unlabeled probes or mutated probes were added to the reaction mixture 15 min before addition of the labeled probes. For the super-shift assay, 10 μg of each of the following antibodies were added to the reaction mixture and then incubated on ice for 30 min before addition of the labeled probes: anti-NRF1 (ab175932; Abcam, United Kingdom) and anti-CMYB (sc-7874; Santa Cruz). Finally, the DNA-protein complexes were separated on a 6% nondenaturing polyacrylamide gel by polyacrylamide gel electrophoresis using 0.5 × Tris-borate-ethylenediaminetetraaceticacid buffer for 1 h. Images were captured using the molecular imager ChemiDoc™ XRS+ system (Bio-Rad, Hercules, CA).
Chromatin immunoprecipitation assays
Inguinal adipose tissue from Qinchuan fetal cattle (n = 3) was isolated 3 days after birth. The chromatin immunoprecipitation (ChIP) assays were performed using the SimpleChIP® Enzymatic Chromatin IP kit (Cell Signaling Technology, Danvers, MA) according to the manufacturer's protocol. The DNA-protein complexes were cross-linked with 37% formaldehyde and neutralized with glycine. After digesting the DNA into fragments of ∼200–1000 bp in length using micrococcal nuclease, the fragmented chromatin samples were suspended in ChIP dilution buffer as the input. An equal amount of cross-linked chromatin was immunoprecipitated overnight at 4°C with 4 μg of anti-NRF1 (ab175932; Abcam) and anti-CMYB (sc-7874; Santa Cruz) antibodies and with normal rabbit immunoglobulin G (IgG). The immunoprecipitated products were isolated using protein G agarose beads, and the bound chromatin was collected by salt washing. The bound chromatin in the ChIP Elution Buffer was eluted and digested with proteinase K, followed by purification for PCR analysis. The immunoprecipitated products from the normal rabbit IgG group were used as a negative control. ChIP primers used in the standard PCR and quantitative real-time PCR experiments are listed in Table 1. The percent input was calculated as follows: % Input = 2 [−ΔCt(Ct[ChIP] − (Ct[Input] − Log2(Input dilution factor)))] (Chakrabarti et al., 2002).
Determination of DNA methylation status by bisulfite conversion
Genomic DNA was extracted using a TIANamp Genomic DNA Kit (Tiangen). Bisulfite treatment of genomic DNA was performed using the EZ DNA Methylation-Gold Kit (Zymo Research), according to the manufacturer's instructions. The bisulfite sequencing PCR primers were designed using online MethPrimer software and are shown in Table 1. For the bisulfite sequencing, PCR fragments were cloned into the pGEMT-Easy vector system (Promega) (n = 3). For each duplicate T-vector, 10 clones were sequenced to determine the methylation status.
Methylation and demethylation in vitro
Multiple recombinant luciferase reporter constructs were methylated in vitro using 4 μM CpG Methyltransferase (M.SssI; NEB, Ipswich, MA), according to the manufacturer's instructions. The cells were then incubated with 5 μM 5-azacytidine (5-AZA; Sigma, St. Louis, MO) to block methylation.
Statistical analysis
Statistical calculations were performed using the Statistical Analysis System (SAS) v8.0 (SAS Institute, Cary, NC). Statistical significance was determined using the one-way analysis of variance test for gene expression and luciferase assay data. Data are expressed as mean ± standard deviation; p < 0.05 was considered significant (* denotes p < 0.05 and ** denotes p < 0.01).
Results
Bioinformatics and protein structure analysis of bovine SIRT4
The phylogenetic tree revealed the relationship between SIRT4 in different species and the potential evolutionary process. The amino acid sequence of bovine SIRT4 exhibits high similarity to other mammalian homologs, especially those of the Bovidae family (bovine, goat, and sheep) (Fig. 1A). Previous studies have shown that the core promoter (also called the functional proximal minimal promoter) and CpG islands are usually located within −1000/−1 bp of the promoter region (Roy and Singer, 2015; Zabidi and Stark, 2016; Zhao et al., 2016; Wei et al., 2017a, 2017b; Zhang et al., 2018). Therefore, the −1000/−1 bp promoter sequence of the SIRT4 gene of these eight species was used to construct the phylogenetic tree. It was shown that the promoter of bovine SIRT4 still possessed high homology with goat and sheep, and the low homology with mouse and human (Fig. 1B). Bovine SIRT4 spans ∼17.643 kb on chromosome 17 [AC_000174.1 (64875236…64892879)] and contains nine exons and eight introns. An open reading frame of 948 bp was verified, which encoded a protein of 315 amino acids (aa). The functional domain of bovine SIRT4 protein ranges from aa 48 to 309. Furthermore, the bovine SIRT4 protein is comprised of two putative domains: the transit peptide ranges from aa 1 to 29 and mat peptide ranges from aa 30 to 315 (Fig. 1C). The resulting protein had a molecular mass of 35.5878 kDa and a molecular formula of C1591H2509N459O448S11, as analyzed by the ExPASy program. The Swiss EMBnet node server program was used to analyze the transmembrane structures of bovine SIRT4 protein, and the SIRT4 had no transmembrane structure (transmembrane structures consisting of >500 aa are considered significant transmembrane structures) (Fig. 1D). The tertiary structure, including the α-helix, β-fold, β-angle, and first-order random curl of bovine SIRT4 protein, was predicted using the online tool SWISS-MODEL software (Fig. 1E).

Bioinformatics and protein structure analysis of bovine SIRT4.
Analysis of TF binding sites and methylation sites in the SIRT4 promoter
We further analyzed TF binding sites and methylation sites in the −402 bp/+44 bp of SIRT4 promoter region using the Genomatix suite and the MethPrimer program. Analysis revealed a CpG island located in the −352/−15 bp region, which highly coincided with the promoter in the −402/+44 bp region (Fig. 2B). The promoter methylation region of −325/−214 bp was amplified by methylation-specific PCR (MSP) primers, which contains seven CpG sites: −302, −294, −288, −277, −245, −241, and −235. The methylation sites are marked in red letters (Fig. 2A). Moreover, two motifs as TF binding sites for NRF1 and CMYB were identified in this region, which contained four methylation sites (Fig. 2A, B).

Analysis of TF binding sites and methylation sites in the bovine SIRT4 promoter.
Identification of NRF1 and CMYB as transcriptional activators or repressors in the SIRT4 promoter of bovine adipocytes
To clarify the functions of TFs in the regulation of bovine SIRT4, we constructed a series of DNA plasmids with 4-bp point mutations in the TF binding motifs and transfected these into 3T3-L1 and bovine adipocytes. The mutation in the NRF1 site in the construct pGL3 − 402/+44 caused a significant increase in SIRT4 promoter activity, while the mutation in the CMYB site in the construct pGL3 − 402/+44 caused a significant decrease in SIRT4 promoter activity (Fig. 3A; p < 0.05). Subsequently, siRNAs against NRF1 and CMYB were designed, synthesized, and transfected into bovine adipocytes to detect the knockdown efficiency of the respective genes and the interaction between the gene expression of NRF1 and CMYB. The results indicated that the expression levels of NRF1 and CMYB were effectively reduced. siNRF1 had no significant effect on CMYB expression; in addition, siCMYB had no significant effect on NRF1 (Fig. 3B; p < 0.05). Further, the siRNAs were utilized to determine the effect of TF knockdowns on SIRT4 expression. The results showed that knockdown of NRF1 upregulated the SIRT4 mRNA level, while knockdown of CMYB downregulated SIRT4 mRNA expression (Fig. 3C; p < 0.05). To further validate these results, siRNAs targeting these TFs and pGL3 − 402/+44 were co-transfected, and they were shown to dramatically alter the activity of the SIRT4 promoter. Transcriptional activity of SIRT4 promoter was significantly increased with knockdown of NRF1, whereas significantly decreased with knockdown of CMYB (Fig. 3D; p < 0.05).
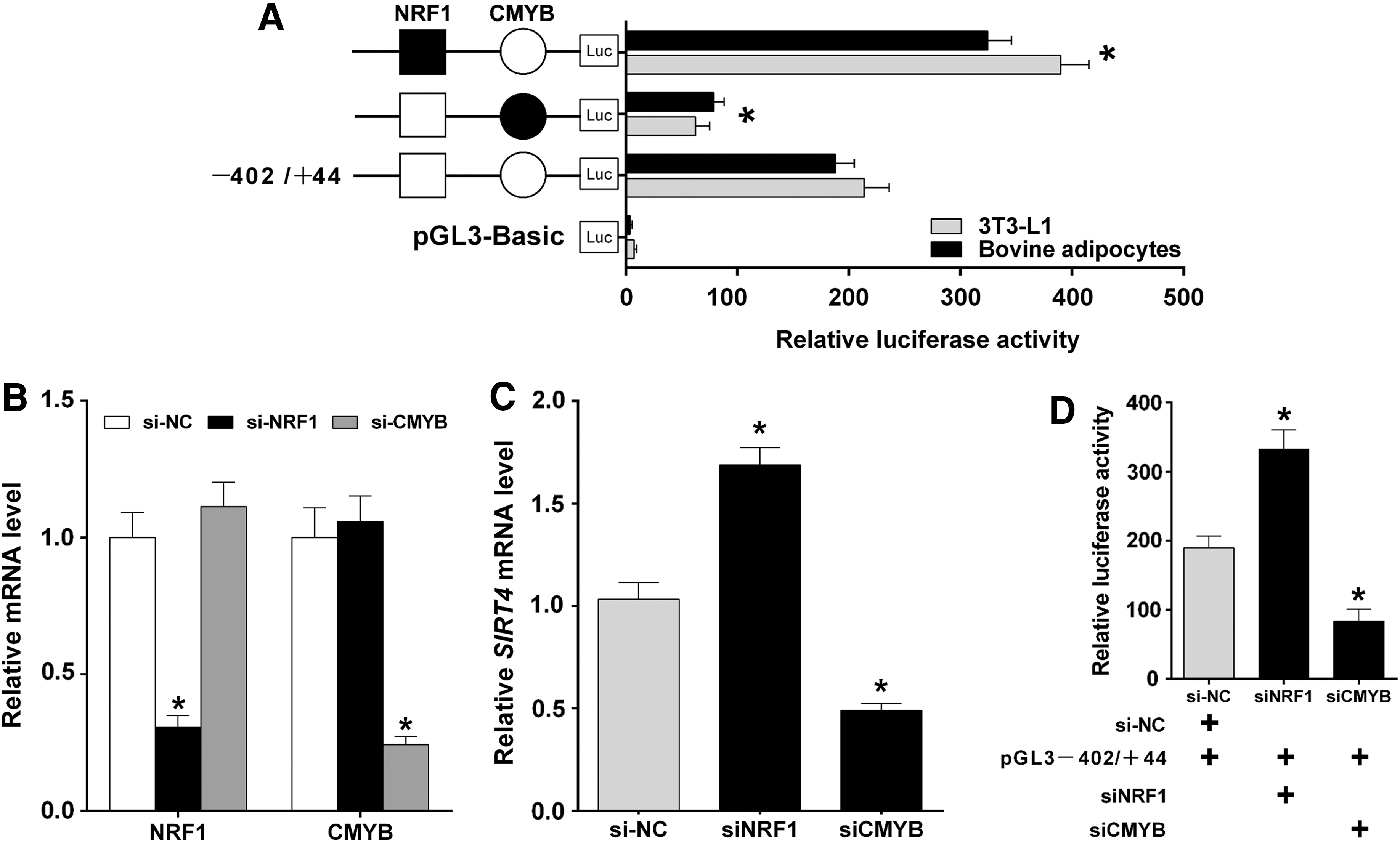
Identification of NRF1 and CMYB binding sites for transcriptional repression and activation in the SIRT4 promoter.
NRF1 and CMYB bind to the promoter of SIRT4 in vitro and in vivo
EMSAs were performed to assess binding of NRF1 and CMYB to the SIRT4 promoter. As shown in Figure 4A, the bovine adipocyte nuclear protein bound the 5′-biotin labeled NRF1 probes and formed one predominant complex (lane 2, Fig. 4A). Competition assays determined that the mutant probe had little effect on this complex (lane 3, Fig. 4A). Additionally, the specific NRF1/DNA interaction was prevented through competition of the excess unlabeled DNA (lane 4, Fig. 4A). The last lane indicates that the complex was super-shifted upon incubation with an anti-NRF1 antibody (lane 5, Fig. 4A). CMYB probes yielded similar results (Fig. 4B). We found that the upshifted bands were diminished when the antibodies were added and incubated, indicating that NRF1 and CMYB bound to the promoter of SIRT4 in vitro (Fig. 4A, B).
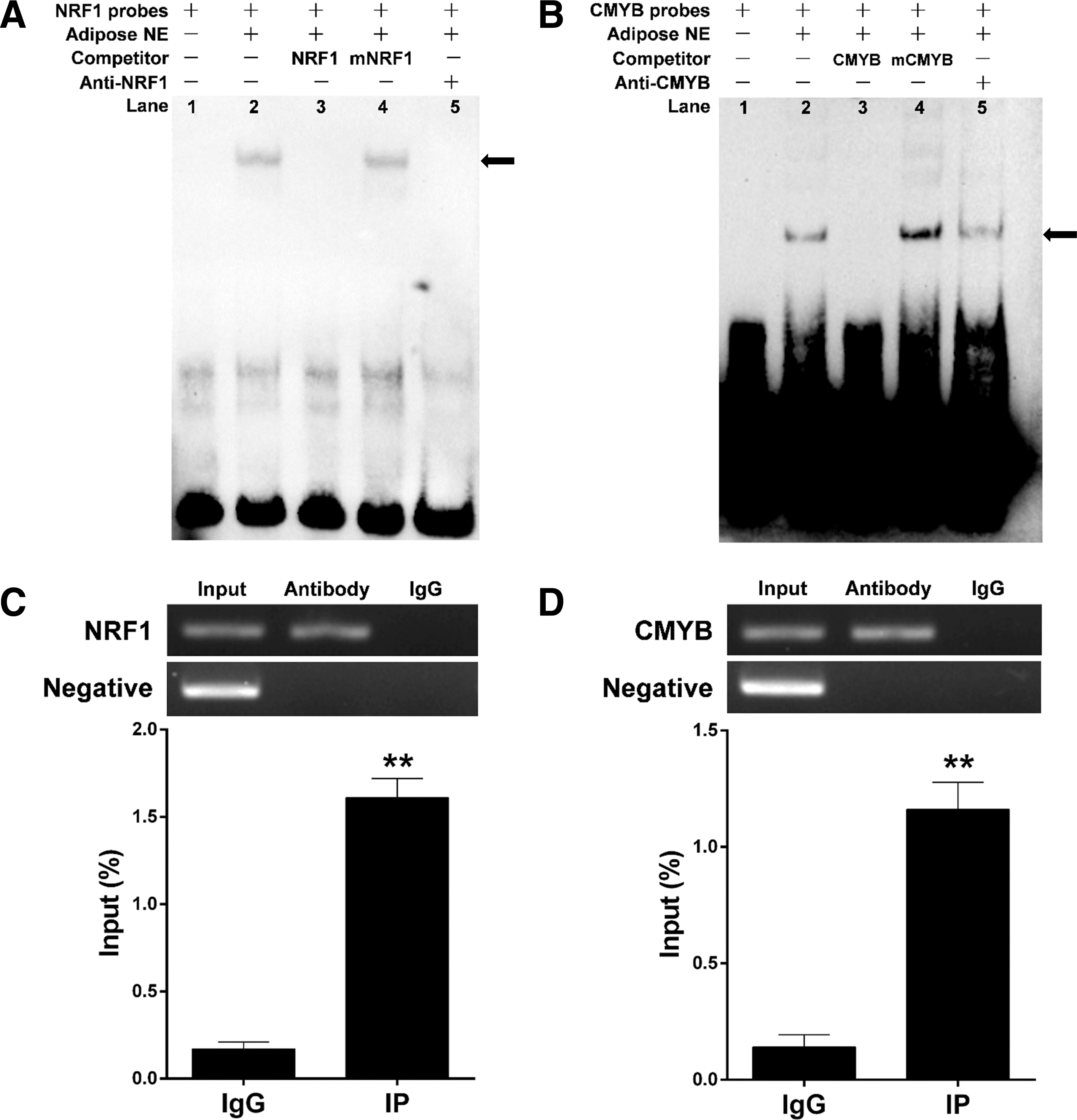
Identification of NRF1 and CMYB binding to the SIRT4 promoter based on EMSA and ChIP assays.
The results of ChIP assays showed that the TFs NRF1 and CMYB interact with their corresponding binding site. Based on three independent experiments, the relative enrichment levels were 9.47- and 8.29-fold higher than in the IgG control, respectively (Fig. 4C, D).
Methylation analysis of SIRT4 promoter at different developmental stages of bovine adipocytes
There is a CpG island located in the −352/−15 bp region of the SIRT4 promoter, and the −325/−214 bp region can be amplified by MSP primers. We found that the methylation level of seven CpG sites in the SIRT4 promoter of differentiated adipocytes was significantly higher than that in undifferentiated adipocytes (Fig. 5A; p < 0.05). However, this increased methylation level of seven CpG sites during adipocyte differentiation was reversed by 5-AZA, which is a DNA methyltransferases (DNMTs) inhibitor that reduces enzyme activity and expression levels of DNA methyltransferases (Lyko and Brown, 2005; Ding et al., 2012) (Fig. 5B; p < 0.05).
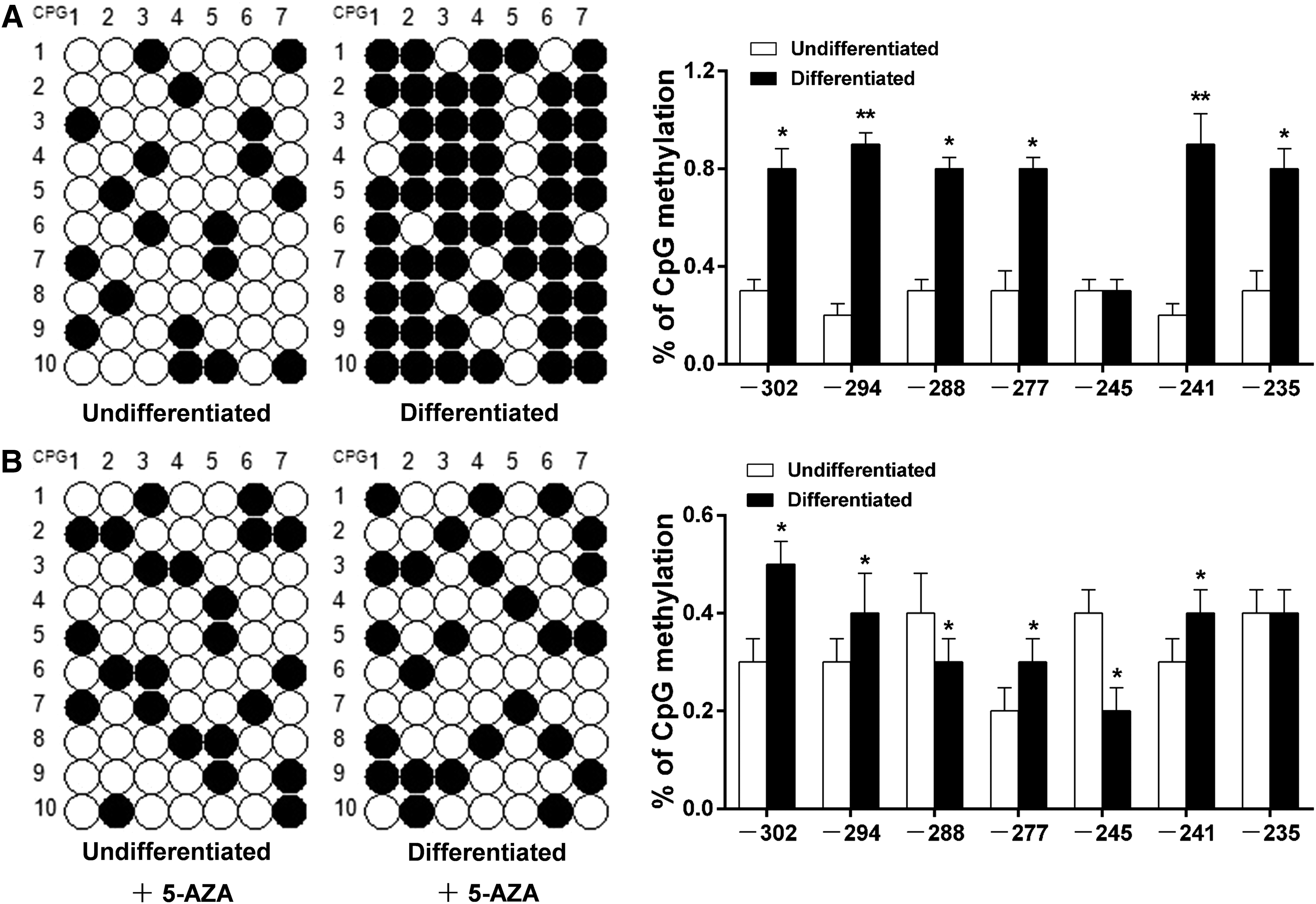
Methylation analysis of seven CpG sites in the SIRT4 promoter during different developmental stages of bovine adipocytes.
SIRT4 promoter activity is competitively regulated by NRF1, CMYB, and methylation during bovine adipocytes differentiation
Since we determined that SIRT4 promoter activity might be regulated by a combination of NRF1, CMYB, and methylation, we further explored the relationship between transcriptional regulation and methylation with respect to SIRT4 promoter activity. For this, we established a demethylation and hypermethylation model using bovine adipocytes. The promoter activity of SIRT4 in bovine adipocytes was assessed by luciferase reporter assays. As such, we found enhanced SIRT4 promoter activity after NRF1 knockdown, and it was reduced after CMYB knockdown in bovine adipocytes (Fig. 6A; p < 0.05). In addition, SIRT4 promoter activity was enhanced by demethylation, and the NRF1-mediated transcriptional inhibition by NRF1 and CMYB-mediated transcriptional activation by CMYB of SIRT4 expression were strengthened by demethylation (Fig. 6A; p < 0.05). We also measured the effect of demethylation and TFs knockdown on SIRT4 mRNA expression in both undifferentiated and differentiated adipocytes, and we first confirmed whether the differentiation degree of the adipocytes was appropriate. Further, we found that the expression level of NRF1 significantly decreased after adipocyte differentiation, while the expression level of CMYB significantly increased (Fig. 6B; p < 0.05). The mRNA expression result indicated that the SIRT4 mRNA level was upregulated by NRF1 knockdown and downregulated by CMYB knockdown, independent of 5-AZA treatment in the undifferentiated and differentiated stages (Fig. 6C, D; p < 0.05). In the differentiated adipocytes, demethylation had a more effective regulation of SIRT4 expression than TFs. This also indicated that during adipocyte differentiation, methylation gradually weakened TF regulation of SIRT4 expression (Fig. 6C, D; p < 0.05). Moreover, the mRNA levels of methylation-related genes DNMT1, DNMT3A, and DNMT3B were decreased by 5-AZA, whereas TET1, TET2, and TET3 were increased by 5-AZA, in both the undifferentiated and differentiated stages (Fig. 6C, D; p < 0.05).
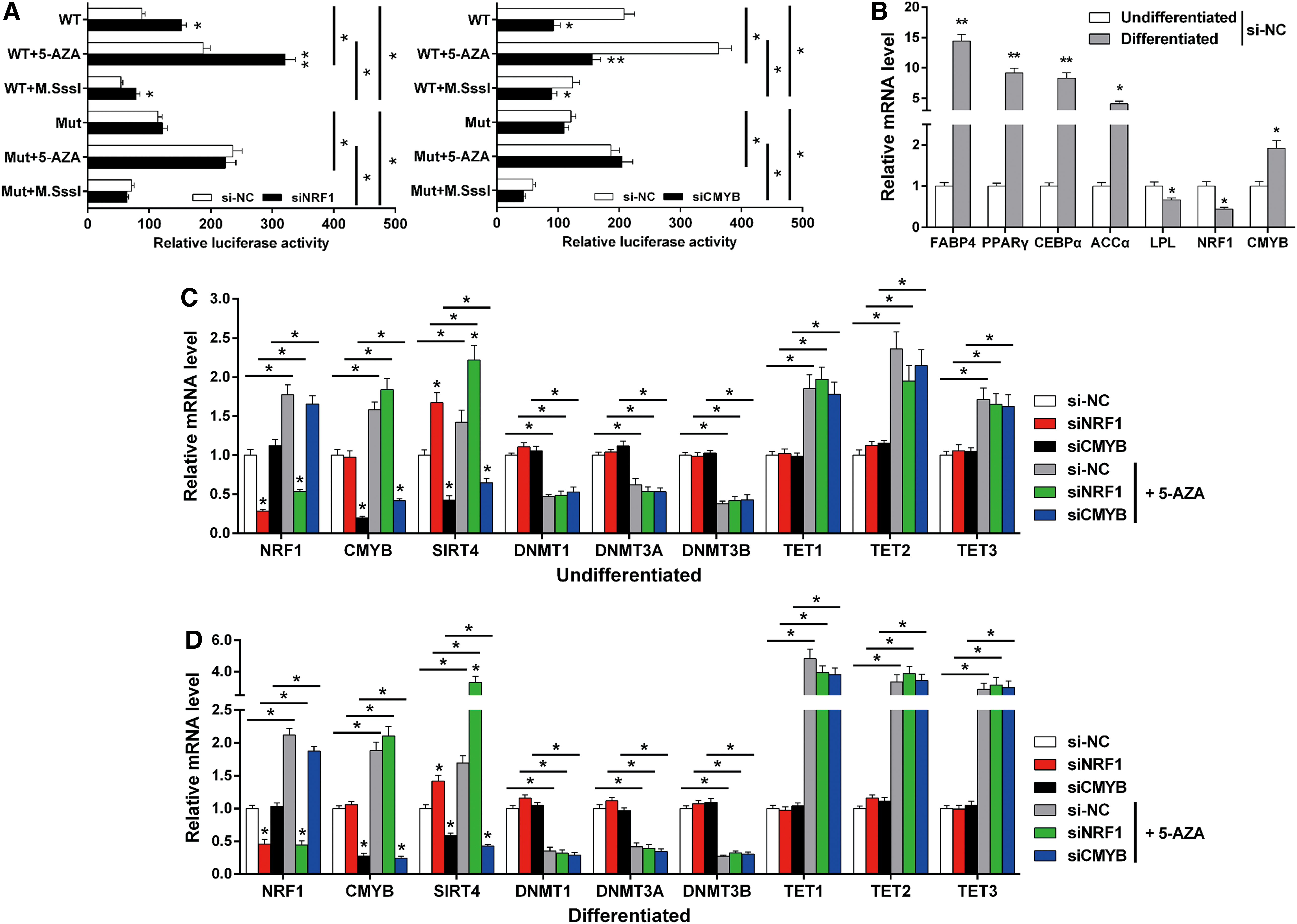
SIRT4 promoter activity is competitively regulated by NRF1, CMYB, and methylation during bovine adipocyte differentiation.
Discussion
Sirtuins have been shown to affect a variety of cellular processes, such as aging, transcription, apoptosis, inflammation, stress resistance, and energy metabolism (Schlicker et al., 2008; Park et al., 2013). To achieve metabolism regulation, sirtuins directly interact with enzymes and alter enzymatic activity in metabolism reactions (Imai and Guarente, 2016). Sirtuins interact with specific pathways and intracellular signaling molecules in various cells. These pathways and signaling molecules include adenosine monophosphate-activated protein kinase (AMPK)-angiotensin-converting enzyme 2 signaling, manganese superoxide dismutase, mammalian target of rapamycin, and others (Wyman and Atamas, 2018). Sirtuins also facilitate DNA repair, and this repair slows the aging process (Vazquez et al., 2017).
Mammalian SIRT4 is localized to the mitochondria and plays an important role in regulating mitochondrial gene expression and fatty acid oxidation in the liver, adipocytes, and muscle cells. Knockdown of SIRT4 results in increased fatty acid oxidation, cellular respiration, and pAMPK levels in hepatocytes (Nasrin et al., 2010; Laurent et al., 2013b; Wang et al., 2017b). SIRT4 inhibition increases the fat oxidative capacity in the liver and mitochondrial function in muscle, and there is also compelling evidence that SIRT4 is an important metabolic regulator via its effects on insulin secretion in beta cells (Ahuja et al., 2007). Therefore, SIRT4 might provide a potential therapeutic target for diseases associated with ectopic lipid storage, such as type 2 diabetes.
NRF1 activates expression of some key metabolic genes regulating cellular growth, and nuclear genes required for respiration, heme biosynthesis, and mitochondrial DNA transcription and replication. In addition, it connects the transcriptional modulation of key metabolic genes to cellular growth and development (Lezza et al., 2001; Adam et al., 2010; Radde et al., 2016). NRF1 also has an important function in integrating the transcription of nuclear-encoded and mitochondrial-encoded genes, which are essential for higher mitochondrial fatty acid β-oxidation capacity (Leal et al., 2012; Widenmaier et al., 2017). Studies have shown that the reduced expression of fatty acid β-oxidation enzyme-encoding genes in type 2 diabetic subjects is closely correlated with NRF1 downregulation (Mootha et al., 2003; Patti et al., 2003). Moreover, it plays an essential role in integrating nucleo-mitochondrial interactions (Li et al., 2012).
CMYB is also known as MYB proto-oncogene protein, which has an important role in regulating the balance between cell division, differentiation, and apoptosis (Chen et al., 2007). CMYB is a central factor in hematopoiesis and stem cell maintenance, and specifically it plays a key role in tumorigenesis (Sebastian et al., 2012). Studies have shown that SIRT4 upregulates E-cadherin expression and suppresses proliferation, migration, and invasion through inhibition of glutamine metabolism in colorectal cancer cells (Miyo et al., 2015). SIRT4 expression in cancer decreased with the progression of invasion and metastasis, and a low expression level of SIRT4 was correlated with a worse prognosis (Wang et al., 2015; Sun et al., 2018). Moreover, SIRT4 has been identified as a tumor suppressor, which regulates the cellular metabolic response to DNA damage by inhibiting mitochondrial glutamine metabolism (Jeong et al., 2013). In tumor cells, SIRT4 is involved in the homeostatic control of glucose metabolism. Therefore, we have reason to speculate that CMYB regulates the tumor suppressor function of SIRT4 by binding to the SIRT4 promoter.
In this study, to clarify the regulatory mechanism of bovine SIRT4 expression, we analyzed the 5′ regulatory region of bovine SIRT4 using the Genomatix suite and determined potential TF binding sites for NRF1 and CMYB at −402/+44 bp of the SIRT4 promoter. We hypothesized that these potential TF binding motifs might play major roles in regulating the transcriptional activity of SIRT4. Following, we determined that NRF1 and CMYB binding sites were transcriptional repression and activation sites in the SIRT4 promoter region, respectively. This result clarifies the novel relationship between these vital TFs and SIRT4 expression. Although studies have shown that most TFs have positive regulatory effects on the promoter activity of target genes, however, the negative regulation of TFs on target genes were also quite common (Zhao et al., 2016; Wei et al., 2017b, 2018; Cao et al., 2018a, 2018b). This may be caused by the following reasons: the binding sites of adjacent TFs usually overlap (or contain each other) and share a promoter sequence. As a result, if a TF binding site is highly overlapping with several adjacent TF binding sites, it will significantly affect the bindings of other adjacent TFs to the promoter when this TF binds to the promoter sequence of the target gene. This competitive regulation ultimately manifests itself as negative regulation of this TF on the promoter activity.
Processes that regulate gene transcription are directly under the influence of the genome organization, and the epigenome contains additional information that is not brought by the DNA sequence. DNA methylation on CpGs constitutes an epigenetic mark generally correlated with transcriptionally silent condensed chromatin (Métivier et al., 2008). It has been reported that DNA methylation might inhibit the association between TFs and specific motifs (Maurano Matthew et al., 2015). Interestingly, some TFs can bind to methylated regions to activate their demethylation (Hashimoto et al., 2017; Hwang and Zukin, 2018). Herein, we assumed that NRF1 and CMYB bindings to sites in the SIRT4 promoter were enhanced by demethylation in bovine adipocytes. Accordingly, results confirmed the hypothesis. Our finding revealed that the activity of the bovine SIRT4 promoter was competitively regulated by NRF1, CMYB, and methylation.
Conclusion
In this study, we determined that bovine SIRT4 is regulated by various TFs, and results indicated that NRF1 transcriptionally represses while CMYB transcriptionally activates SIRT4 promoter activity, as determined by luciferase assays, EMSAs, and ChIPs. Furthermore, we found that demethylation significantly enhanced binding of NRF1 and CMYB to the SIRT4 promoter. All these data provide a foundation for a better understanding of the transcriptional regulation and biological function of SIRT4 in adipocytes (Fig. 7).

A proposed schematic summary of the SIRT4 transcriptional activity regulation in bovine adipocytes by DNA methylation and TFs, including NRF1 and CMYB. Color images available online at
Footnotes
Acknowledgments
This study was supported by the National Key Research and Development Program of China (No. 2018YFD0501700); National 863 Program of China (No. 2013AA102505); National Science and Technology Support Projects (No. 2015BAD03B04); and National Modern Agricultural Industry Special Program (No. CARS-37).
Disclosure Statement
No competing financial interests exist.