
Abstract
Intervertebral disk degeneration (IDD) is a common disease that is caused by degeneration of the nucleus pulposus (NP). One goal in the treatment of IDD is delaying or reversing the degeneration of NP via the transformation of exogenous genes. This study first investigated the role of BMP9 in the extracellular matrix (ECM) of nucleus pulposus cells (NPCs) and its mechanism. We found that BMP9 promotes the expression of ECM in NPCs, and the key molecules of Notch signaling, namely, NICD-1, hes and hey, and it was significantly altered in BMP9-transfected NPCs, which suggests that BMP9 may regulate the ECM via the Notch signaling pathway. We verified the expression of Notch ligands and receptors in NPCs infected with Ad-BMP9 and demonstrated a significant decrease in DLL1 and Notch1; then, NPCs were transfected with Ad-dnNotch1, Ad-Jagged1, and Ad-DLL1, and different multiple groups were established to further identify the ligands or receptors that affected ECM expression. The results demonstrated that Ad-dnNotch1, Jagged1 and DLL1 inhibited ECM expression, and dnNotch1 promoted expression. Therefore, we demonstrated that BMP9 promoted the expression of ECM in NPCs via inhibition of Notch1 and DLL1. This study provides a possible method for IDD treatment.
Introduction
Intervertebral disk degeneration (IDD) is a degenerative disease caused by multiple factors (Wang et al., 2017a; Holguin and Silva, 2018). Morbidity increases gradually with the aging of the population and the prolongation of life expectancy, which brings great pain to patients and heavy social burden (Holguin and Silva, 2018; Li et al., 2018). Nucleus pulposus (NP) degeneration is the main pathological change of IDD in the early stage, which is manifested as the gradual change of extracellular matrix (ECM). The ECM is primarily composed of aggrecan and type II collagen (Col2a1), which are gradually reduced, and type I collagen (Col1a1) replaces Col2a1, which results in fibrosis and the loss of the elastic modulus (Wang et al., 2017a; Henry et al., 2018). A corresponding deficiency in the treatment of NP degeneration includes surgery or conservative treatment (Hiyama et al., 2012). Therefore, the introduction of exogenous genes to promote the ECM of nucleus pulposus cells (NPCs) is an ideal method.
BMP9 is a member of the TGF-β superfamily (Lamplot et al., 2013), and it is a strong growth differentiation factor that was originally isolated from the livers of growing mice (Song et al., 1995). It plays an important role in the promotion of chondrogenic differentiation of mesenchymal stem cells (Ren et al., 2016). The main components of the ECM in NP and cartilage are similar, and their essence is aggrecan and Col2a1. Therefore, we investigated whether BMP9 promoted ECM expression to maintain the physiological function of the NP and further elucidated its mechanism of action.
Materials and Methods
NP sample source and inclusion criteria
All NP tissues were obtained from patients who underwent lumbar discectomy in our department. The following inclusion criteria were used: no structural abnormalities of the spine, such as tumors; tuberculosis; suppurative infections; age <60 years; and open surgery. A total of 26 cases, with an average age of 49.8 ± 2.7 years, were included: 8 cases with lumbar spinal stenosis and 18 cases with lumbar disk herniation. Priffmann grading (Pfirrmann et al., 2001) was performed according to preoperative MRI to assess the degree of IDD and included 1 case of grade II, 15 cases of grade III, 8 cases of grade IV, and 2 cases of grade V. All donors signed a written informed consent before surgery, and the Ethics Committee of Chongqing Medical University approved the study protocol.
NPC culture
Intervertebral disk tissue was repeatedly washed with phosphate-buffered saline (PBS) in the Biological Safety Cabinet to remove blood and impurities, and the residual endplate and annulus fibrosus (AF) were separated (Fig. 1A) (Risbud et al., 2006). The NP tissue was cut into small pieces, digested by using 0.25% trypsin (Beyotime, China) for 30 min, and sequentially digested by using 0.2% type II collagenase (Sigma) for 4 h. Lysates were filtered through a 200-mesh filter, and the cell-containing liquid was collected and centrifuged. Pellets were resuspended in DME/F12 medium (Gibco) containing 17% fetal bovine serum (Corning) and placed in an incubator at 5% CO2 and 37°C. Only P1 passage cells were used for all experiments. Only cells of the same source were selected for a single experiment to reduce error.

NP isolation, cell culture, and identification.
Immunofluorescence and NPC identification
Primary NPCs were digested, resuspended in medium, seeded on sterile cell slides in a 24-well plate at 5 × 104 cells/well, and cultured for 24 h. Cells were washed with PBS, and 4% paraformaldehyde was used to fix the cells for 20 min at room temperature. Cells were washed three times with PBS, and 0.5% Triton was added for 10 min for perforation. The cells were washed with PBS, and goat serum was used to block nonspecific binding for 1 h. The serum was completely aspirated, and cells were incubated with a primary antibody (rabbit anti-Col2a1 or mouse anti-aggrecan [Abcam]) at 4°C overnight. The plate was placed at room temperature for 1 h and washed with PBS. Subsequent steps were performed while avoiding light. Secondary antibodies (goat anti-rabbit, red fluorescence; goat anti-mouse, green fluorescence) were incubated for 1 h. Cells were washed with PBS, and DAPI was used to stain the nuclei for 10 min. Slides were washed with PBS, transferred to a glass slide, and photographed by using a fluorescence microscope after sealing with glycerin. Immunofluorescence detection of the expression of ECM in NPCs was performed by using a secondary antibody with red fluorescence in subsequent experiments.
Adenovirus infecting NPCs
The adenoviruses used in this study included Ad-GFP, Ad-BMP9, Ad-RFP, Ad-DLL1, Ad-Jagged1, and Ad-dnNotch. Ad-GFP and Ad-BMP9 carry green fluorescence, and the other viruses carry red fluorescence (Fig. 1D). P0 passage NPCs were digested by using 0.25% trypsin, passaged, and returned to the incubator for 4 h. Most of the cells adhered to the plate at this time. Polybrene and different groups of adenovirus were added (MOI value 100) (Wang et al., 2017b), and the infection efficiency was maintained at 50%. The medium was replaced 6–8 h after infection, and the infection was observed under a fluorescence microscope the next morning. Subsequent experiments were performed after infection for 24 h.
Cell proliferation assay
The effect of BMP9 on the proliferation of NPCs was detected by using the CCK8 assay. Except for specific instructions, all the experiments used Ad-GFP as the control group and followed the infection methods mentioned earlier, and Ad-GFP- and Ad-BMP9-infected cells were compared. Cells were seeded in a 96-well plate at 1 × 104 cells/well, and each group used three duplicate wells. CCK8 reagent (10 μL) was added 24 h after cell infection, and plates were returned to the incubator for 2 h. Absorbance was measured at 450 nm by using an automicroplate reader (Tecan Infinite 200 PRO, Switzerland).
Flow cytometry
Flow cytometry was used to detect the effect of BMP9 on the NPC cycle. NPCs infected with Ad-GFP and Ad-BMP9 were digested by using trypsin and collected by using low-speed centrifugation. The cells were washed with precooled PBS, fixed for 12 h in precooled 70% ethanol at 4°C, and washed with precooled PBS. A propidium iodide staining solution (one sample: staining buffer 0.5 mL, propidium iodide [20 × ] 25 mL, RNase A [50 × ] 10 mL) was used according to the manufacturer's instructions (Beyotime). Staining solution (0.5 mL) was added to each sample, and the cells were slowly resuspended. Cells were reacted at 37°C for 30 min in light-proof conditions and sorted by using flow cytometry (BD FACSCanto).
RNA extraction and semi-quantitative RNA analysis
Each group of NPCs with adenovirus infection was collected for RNA extraction by using the GeneJET RNA Purification kit (Thermo Scientific). The RNA concentration was measured by using a spectrophotometer (Thermo Scientific). cDNA was produced by using reverse transcription and the RevertAid First Strand cDNA Synthesis Kit (Thermo Scientific). The following reaction conditions were used: incubation for 5 min at 25°C, followed by 60 min at 42°C. The reaction was terminated via heating at 70°C for 5 min. The cDNA was used directly for PCR application by using Dream Taq Green PCR Master Mix (2 × ) (Thermo Scientific). The following reaction conditions were used: initial denaturation at 95°C for 2 min for 1 cycle; denaturation at 95°C for 30 s, annealing at 55°C for 30 s, and extension at 72°C for 1 min; ➁ to ➃, for 35 cycles; and a final extension at 72°C for 5 min. Gel electrophoresis was performed by using a 1.5% agarose gel for 35 min at 80 V. A gel imager (Bio-Rad Gel Doc XR+) was used to expose bands. Table 1 shows the RNA primer sequences.
List of Primers Used in This Study
Protein extraction and Western blotting
Each group of adenovirus-infected cells was washed with PBS and collected via centrifugation. The cells were fully lysed on ice by using RIPA buffer, and the protein concentration was measured by using an Enhanced BCA Protein Assay Kit (Beyotime). The loading of each well was controlled at 50 μg, and the lysates were subjected to 6–12% sodium dodecyl sulfate-polyacrylamide gel (SDS-PAGE). Separated proteins were transferred to polyvinylidene difluoride membranes. Membranes were blocked in 5% skim milk for 1 h at room temperature and incubated overnight at 4°C with the primary antibodies anti-hes1 (Cell Signaling), anti-GAPDH (Bioss, China), and anti-Col2a1, anti-aggrecan, anti-hey1, anti-hey2, anti-heyL, and anti-NICD-1 (Abcam). The membranes were washed repeatedly with TBST and incubated with a horseradish peroxidase-conjugated secondary antibody. Membranes were washed, and a gel imaging system (Vilber Lourmat, France) and BeyoECL Plus (Beyotime) were used for band visualization.
Statistical analysis
Results were calculated as the mean ± standard deviation. Two groups were paired, and the difference was analyzed by using a t-test. Differences in multiple groups were repeatedly matched and analyzed by using one-way analysis of variance (ANOVA) and Tukey's post hoc test using SPSS 22.0 version statistical software. p < 0.05 was regarded as statistically significant.
Results
Identification of NPCs
The intervertebral disk is composed of the endplate, AF and NP, which is located in the middle of the endplate and the AF (Gogate et al., 2011). The NP was isolated and digested, and the NPCs were extracted and cultured. Primary NPCs are round or polygonal under a microscope and grow in a cluster. The P1 generation was polygonal or fusiform (Fig. 1B). NPCs are chondrocyte-like cells (Poiraudeau et al., 1999) that express a cartilage-like ECM, such as aggrecan and Col2a1, to absorb and transmit external force concussions; this ECM differentiates NP from AF and endplate tissue (Rodrigues-Pinto et al., 2013). The presence of aggrecan and Col2a1 in the cells was demonstrated by using immunofluorescence (Fig. 1C), which indicated that the cells were NPCs.
Expression of BMP9 in NPCs
BMP9 was originally isolated from mouse liver (Song et al., 1995), but its expression in human NPCs was not known. We determined its endogenous expression in NPCs by using semi-quantitative PCR and whether it was successfully expressed after Ad-BMP9 infection of NPCs. The results demonstrated no endogenous BMP9 expression observed in the NPCs, but transfection significantly increased BMP9 expression (Fig. 2A).
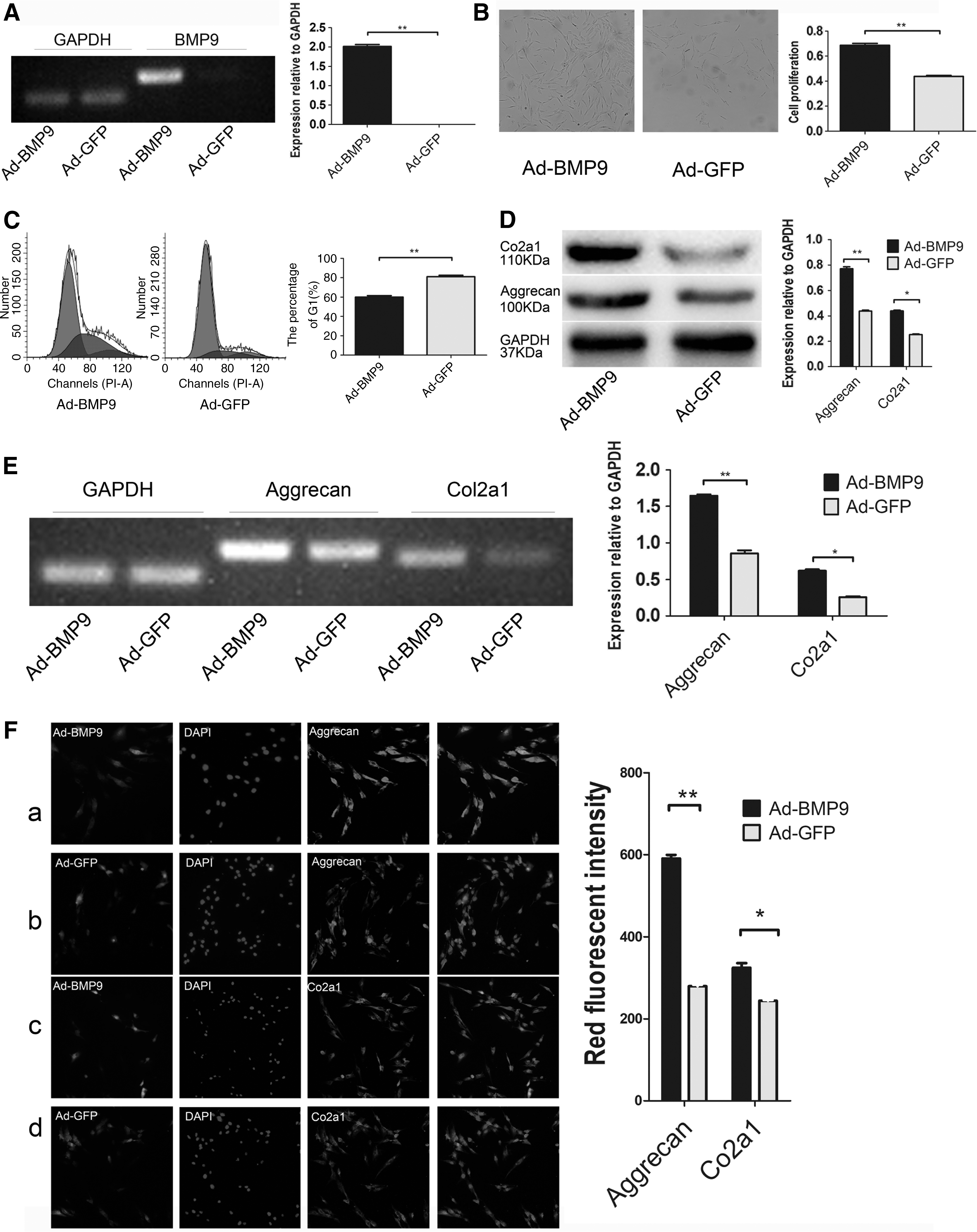
BMP9 promotes proliferation and ECM of NPCs. Ad-BMP9 as the experimental group, and Ad-GFP as the control group.
BMP9 promotes NPC proliferation
The number of NPCs is decreased in degenerated NP, and the growth is slow. BMP9, also known as growth differentiation factor-2, significantly promotes cell proliferation (Song et al., 1995). The CCK8 assay found that BMP9 accelerated the growth of NPCs (Fig. 2B). Flow cytometry revealed that the G1 phase of NPCs transfected with BMP9 decreased significantly, but the S phase increased, and the G2-M phase increased slightly (Fig. 2C).
BMP9 promotes ECM expression in NPCs
Aggrecan and Col2a1 are the major components of the ECM that maintain the cartilage-like properties of the NP (Risbud et al., 2006; Wang et al., 2017a). We detected the expression of related RNA by using semi-quantitative PCR. Western blotting and immunofluorescence were used to detect proteins in the cells and to demonstrate the effect of BMP9 on ECM. The results revealed an upregulation of related RNA expression in the NPCs infected with Ad-BMP9, and protein was increased, especially aggrecan (Fig. 2D–F).
Relationship between BMP9 and the Notch signaling pathway
The BMP family plays a physiological role via classical Smads and MAPK signaling pathways (Imamura et al., 2006). However, the mechanism of action of BMP9 does not follow the traditional classical pathway and may be related to Notch signaling (Kang et al., 2009; Sharff et al., 2009). We used semi-quantitative PCR and WB to detect the expression of the activated form of the Notch signaling pathway, NICD-1 (Active form of NICD), and target genes after BMP9 infection (Bolós et al., 2007; Watt et al., 2008). NICD-1 and the target genes hes-1 and hes-5 were reduced significantly (Fig. 3A, B). The target genes hey1, 2, and L were also altered to varying degrees, which indicates that BMP9 is closely related to Notch signaling.

Association between BMP9 and the Notch signaling pathway.
Expression of Notch signaling in NPCs
The Notch signaling pathway is composed of the Notch 1, 2, 3, and 4 receptors and the ligands Jagged1 and 2 and DLL1, 3, and 4 (Zanotti et al., 2016). We investigated the expression of Notch ligands and receptors in the control group. Jagged1 was the highest expressed ligand, and no Jagged2 or DLL3 expression was found (Fig. 3C). The expression of each receptor was different (Fig. 3C).
Effect of BMP9 on Notch signaling ligands and receptors in NPCs
The experiments cited earlier indicated that BMP9 was closely related to the Notch signaling pathway. We detected changes in ligands and receptors in NPCs after BMP9 infection by using semi-quantitative PCR. The results demonstrated that Notch1 and DLL1 decreased significantly, and the other receptors and ligands changed slightly (Fig. 3D).
Effect of Notch ligands and receptors on ECM
Previous studies demonstrated that BMP9-infected NPCs exhibited reduced Notch1 and DLL1 expression, and we examined which molecule affected the expression of ECM. Negative expression Notch1 (Ad-dnNotch1) and overexpression DLL1 (Ad-DLL1) were used to infect NPCs to differentiate the role of each molecule. Jagged1 expression was significantly higher than the other ligands and receptors before and after infection. Therefore, we suspected that Jagged1 was involved in promoting ECM expression. We used the following groups for analyses: (1) Ad-GFP+Ad-RFP, Ad-BMP9+Ad-RFP, Ad-GFP+Ad-DLL1, Ad-BMP9+Ad-DLL1; (2) Ad-GFP+Ad-RFP, Ad-BMP9+Ad-RFP, Ad-GFP+AdJagged1, Ad-BMP9+Ad-Jagged1; and (3) Ad-GFP+Ad-RFP, Ad-BMP9+Ad-RFP, Ad-GFP+Ad-dnNotch1, Ad-BMP9+Ad-dnNotch1. Three groups of viruses were infected with NPCs by using the combinations mentioned earlier; proteins and RNA of aggrecan and Col2a1 were extracted, detected, and compared with each other in each group. The results show that: DLL1 group: Ad-BMP9+Ad-RFP>Ad-GFP+Ad-RFP>Ad-GFP+Ad-DLL1, Ad-BMP9+Ad-DLL1. Ad-GFP+Ad-DLL1 and Ad-BMP9+Ad-DLL1 had no statistical difference (Fig. 4A); Jagged1 group: Ad-BMP9+Ad-RFP>Ad-GFP+Ad-RFP>Ad-GFP+Ad-Jagged1, Ad-BMP9+Ad-Jagged1. Ad-GFP+Ad-Jagged1 and Ad-BMP9+Ad-Jagged1 had no statistical significance (Fig. 4B); and dnNotch1 group: Ad-BMP9+Ad-dnNotch1>Ad-GFP+Ad-dnNotch1>Ad-BMP9+Ad-RFP>Ad-GFP+Ad-RFP, and Ad-BMP9+Ad-dnNotch1>Ad-GFP+Ad-dnNotch1>Ad-BMP9+Ad-RFP had no statistical significance (Fig. 4C).
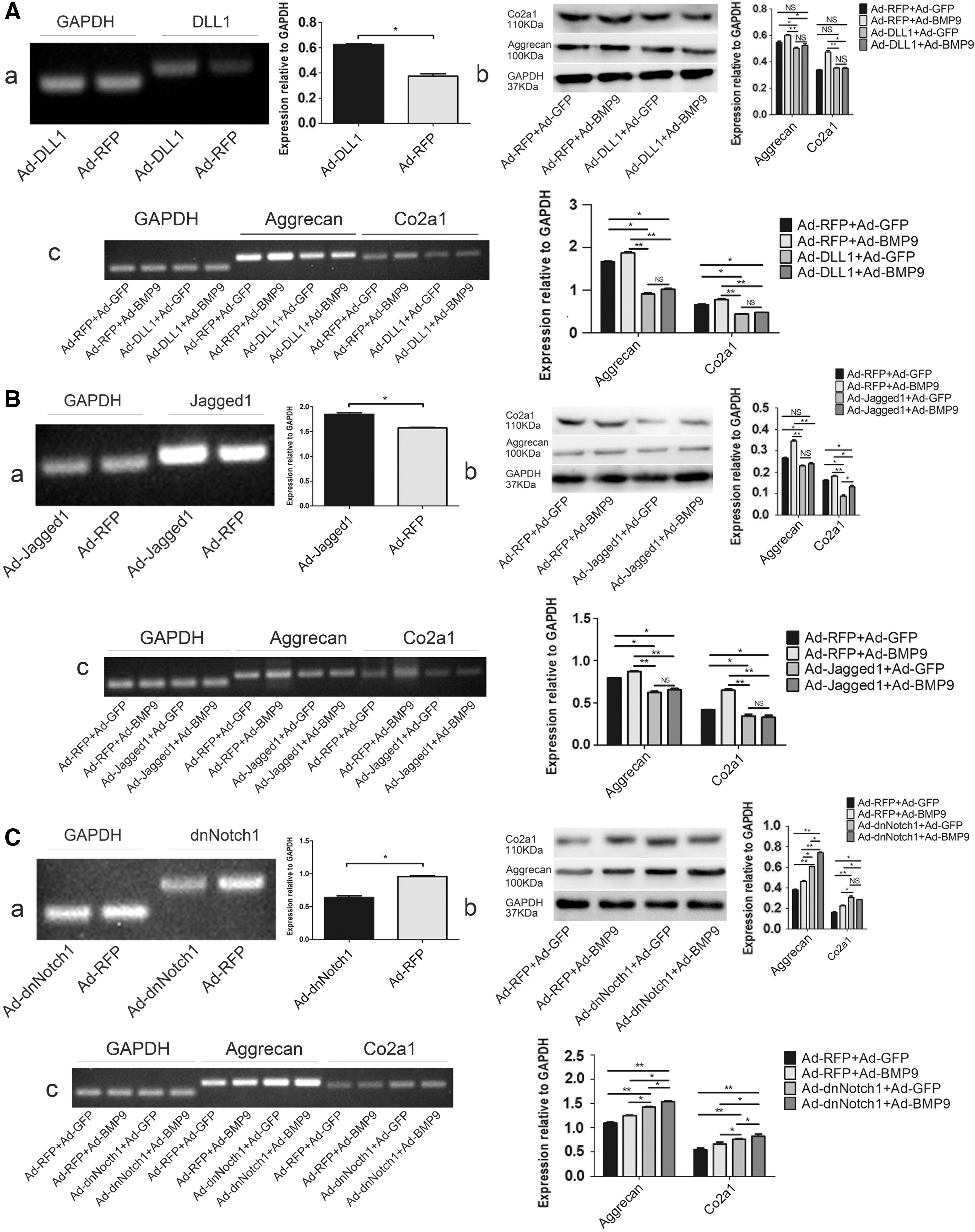
Effects of dnNotch1, DLL1, and Jagged1 on the ECM of NPCs.
Discussion
The phenotypic characteristics of the degenerated NPCs include differentiation from chondrocytes-like to fibroblasts, which further inhibits secretion of the ECM (Henry et al., 2018). The ability of degenerated NPCs to grow was very weak. Our study found that primary NPCs required at least 3 days to exhibit a very small number of adherent cells, and the cell confluence rate in the T25 flask reached 90% in ∼13–15 days. We attempted to seed the cells in a 96-well plate at 1000, 2000, and 5000 cells/well; the CCK8 assay revealed an absorbance of ∼0.2, and it was difficult to reach the baseline value of 0.4. Therefore, we increased the number of cells to 10,000/well for testing. Flow cytometry revealed that the degenerated NPCs were mostly in the G0-G1 phase, which indicates that NPCs grew extremely slowly. The growth ability of NPCs cultured to the P3 generation was extremely low and severe differentiation occurred, and the ability to secrete ECM was significantly reduced. Therefore, only P1 generation NPCs were used in this study. NPCs cultivation requires high-quality sera and media with a strong dependence on serum concentration. The degenerated NPCs were also relatively fragile, and centrifugal speeds greater than 1000 rpm/min caused irreversible cell damage and destruction. These characteristics of degenerative NPCs underlie the degeneration of the IVD and the entry into a vicious circle. Therefore, maintaining and reversing the number of NPCs and promoting ECM by using external interventions is the goal of treatment.
Notch signaling is involved in the regulation of a range of pathophysiological processes (Bolós et al., 2007). The activation pathway of the classical Notch signaling pathway is also called the RBPjk pathway. The ligand acts on the receptor, which releases the intracellular activation form of Notch protein, NICD (ICN), via a series of reactions. NICD enters the nucleus and binds to the CSL protein (RBPjk) to form a ternary complex transcriptional activator (NICD-CSL-MAMK) with the nuclear transcriptional activator family MAM. This complex activates the target genes, hes and hey, to exert physiological effects (Wang, 2011; Xu et al., 2017). Our study found that the expression of NICD-1 (active form of NICD), hes1 and hes5, decreased after infection with BMP9 in NPCs, which also proved that BMP9 and Notch signaling pathways are closely related. Hes1 is an RBPjk-dependent target gene that plays an important role in the inhibition of cartilage development in vitro (Dong et al., 2010), and it may synergize with hes5, forming an interdependent relationship to inhibit cartilage formation. As a chondrocyte-like, the main component of ECM expressed by NPCs is consistent with cartilage, we suggested that BMP9 downregulates hes1 and hes5 to promote ECM expression. Hey1, hey2, and heyL exhibit no obvious relationship with cartilage formation (Rutkowski et al., 2016).
We also found that BMP9-infected NPCs significantly reduced Notch1 and DLL1. Jagged1 expression was quite stable and much higher than other ligands and receptors. Further studies compared the expression of ECM in each group: Ad-GFP+Ad-RFP>Ad-GFP+Ad-DLL1, Ad-BMP9+Ad-DLL1, which indicated that DLL1 had a strong inhibitory effect on ECM; whereas there was no statistical difference between Ad-GFP+Ad-DLL1 and Ad-BMP9+Ad-DLL1, which indicated that overexpression of DLL1 offset the inhibition of DLL1 by BMP9. Ad-BMP9+Ad-dnNotch1>Ad-GFP+Ad-dnNotch1>Ad-BMP9+Ad-RFP>Ad-GFP+Ad-RFP, indicating that dnNotch1 has a role in promoting ECM, and Ad-BMP9+Ad-dnNotch1>Ad-GFP+Ad-dnNotch1>Ad-BMP9+Ad-RFP has statistical significance, indicating that dnNotch1 and BMP9 have synergistic effects, and dnNotch1 is more effective than BMP9. So, we demonstrate that BMP9 promotes ECM by inhibiting DLL1 and Notch1. Ad-GFP+Ad-RFP>Ad-GFP+Ad-Jagged1, Ad-BMP9+Ad-Jagged1, indicating that Jagged1 can inhibit ECM. There is no significant difference between Ad-GFP+Ad-Jagged1 and Ad-BMP9+Ad-Jagged1, indicating that overexpression of Jagged1 offsets the effect of BMP9. Overexpression of Jagged1 inhibited ECM in vitro, whereas Jagged1 exerted a strong cartilage-forming effect in vivo (Hosaka et al., 2013). Therefore, NPCs, which are chondrocyte-like cells, may express a large number of Jagged1 ligands, which reflects the high consistency of biological properties and functions.
In conclusion, our study first demonstrated that BMP9 interfered with NPCs via inhibition of Notch1 and DLL1 in the Notch signaling pathway, which downregulated NICD-1, reduced the target genes hes1 and hes5, and ultimately promoted ECM expression. This study suggests that BMP9 may be a target for the treatment of IDD.
Footnotes
Acknowledgment
This work was supported by the National Natural Science Foundation of China (NNSFC; project approval no. 81501922).
Disclosure Statement
No competing financial interests exist.