
Abstract
The vital role of long noncoding RNAs (lncRNAs) on the acute myeloid leukemia (AML) has been increasingly recognized. This study aims to explore the unknown function of lncRNA LINC00152 in the leukemogenesis of AML. LINC00152 is determined to be upregulated in the AML samples, and the overexpression of LINC00152 is also authenticated in the advanced French–American–British (FAB) AML patients and closely correlated with the poor outcome of AML patients. The functional experiments state that knockdown of LINC00152 suppresses the proliferation, accelerates the apoptosis, and induces the cycle arrest of AML cells. The mechanical experiments state that LINC00152 and CDK9 were both targeted by miR-193a with the complementary binding sites at 3′-UTR. Moreover, in the rescue experiments, the enhanced LINC00152 expression could regain the suppression of tumor behavior induced by LINC00152 knockdown. In conclusion, this research reveals the important role of lncRNA LINC00152 in the AML leukemogenesis through targeting miR-193a/CDK9 axis. This finding could indicate the important pathogenesis of ncRNA and the vital roles of epigenetic regulation.
Introduction
Acute myeloid leukemia (AML) is generated from the uncontrolled proliferation, apoptosis blockage, and differentiation obstacle of hematopoietic cells in bone marrow and other hematopoietic tissue (Harada et al., 2017; Castelli et al., 2018). Although the series of therapies, such as chemotherapy and stem cell transplantation, have applied to the clinical treatment, the prognosis is still fatal and pessimistic (Lagunas-Rangel et al., 2017). Moreover, the chemotherapy resistance, metastasis, and recurrence make it difficult to overcome the spread (Andresen and Gjertsen, 2017; Aziz et al., 2017). Emerging pieces of evidence have indicated the increasing roles of epigenetic dysregulations for the AML, such as acetylation, noncoding RNAs (ncRNAs), and DNA methylation (Finn et al., 2017).
More and more researchers are beginning to notice the importance of ncRNAs in the human disease, especially the long ncRNAs (lncRNAs). lncRNAs are characterized by the no longer than 200 nucleotides without encode protein. For the human cancers, lncRNAs modulate series of the pathophysiological process, for example proliferation, cycle regulation, and cancer progression (Huang et al., 2018). For example, lncRNA LINC00152 is overregulated in hepatocellular carcinoma and LINC00152 facilitates the cell cycle progression through endogenous regulating miR-193a/b-3p targeting CCND1 (Ma et al., 2018). In glioma, LINC00152 level is increased with glioma grade, especially in the mesenchymal TCGA subtype, serving as a potential prognostic biomarker in patients with high-grade glioma (Wang et al., 2018b). However, the role of LINC00152 in the AML is still unclear. Evidence have indicated that miR-193a represses the expression of multiple target genes, such as AML1/ETO, DNMT3a, HDAC3, KIT, CCND1. miR-193a silencing binds the AML1-binding sites and recruits chromatin-remodeling enzymes (Li et al., 2013).
In this study, we focus on the oncogenic RNA gene, long intergenic nonprotein coding RNA 00152 (LINC00152), for the AML tumorigenesis. LINC00152 is identified to be upregulated in the AML patients. Mechanically, the ectopically expressed LINC00152 accelerates the AML proliferation and targets the miR-193a/CDK9 axis to exert its functions.
Materials and Methods
Clinical samples collection
This clinical research was approved by the Ethics Committee of Huai'an Hospital Affiliated to Xuzhou Medical College. All work was done according to the recommendations of the Declaration of Helsinki and the informed consent forms have been obtained from volunteers. Bone marrow samples were enrolled from 33 AML patients diagnosed by French–American–British (FAB) criteria and 15 healthy donors.
AML cell lines and culture
Human AML cell lines (HL-60, THP-1, Kasumi-1, MOLM-13) were purchased from ATCC and normal marrow cells (HS-5) were purchased from Shanghai cell bank of Chinese Academy of Science. AML cells were cultured in Iscove's modified Dulbecco's medium (IMDM; Gibco, Grand Island, NY) with 10% fetal bovine serum (FBS; Thermo Fisher Scientific, Waltham, MA). The atmosphere was set at 37°C containing 5% CO2.
Silencing oligonucleotides and plasmid transfection of cell lines
AML cell lines silenced expression of LINC00152 and enhanced CDK9 expression were performed using siRNA and pcDNA3.1 plasmid. These oligonucleotides were chemically synthesized by Shanghai Genepharm Company (China), and the pcDNA3.1 plasmid was provided by Addgene, Inc. (Cambridge, MA). The sequences are presented in the Supplementary Table S1. The transfection was conducted using Lipofectamine 2000 transfection reagent (Thermo Fisher Scientific) according to the manufacturer's instructions.
Quantitative real-time PCR
Total RNA was extracted from the samples and cells using TRIzol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. Then, the cDNA synthesis was performed using 1 μg RNA (total 20 μL) with SuperScript First-Stand Synthesis system (Invitrogen) by ABI PRISM 7900 thermocycler. Real-time PCR was performed using TaqMan probe-based detection kit (TaqMan PCR universal mastermix, Applied Biosystems, Darmstadt, Germany). GAPDH was the endogenous control. Measuring of lncRNA or miRNAs was performed using the mirVana qRT-PCR miRNA detection kit, according to the manufacturer's instructions. The list of primers for RNA is presented in Supplementary Tables S1.
Colony formation and proliferation assay
The proliferative ability of AML cells was detected using colony formation and proliferation assay. AML cells (THP-1, HL-60) were suspended in single-cell suspensions at the density of 1 × 103 cells in six-well plate. Two weeks later, the colonies were stained with Wright-Giemsa, and then counted. The CCK-8 assay was carried out as previously described (Wang et al., 2016).
Flow cytometry for apoptosis and cycle analysis
The AML cells (THP-1 and HL-60) were transfected with oligonucleotides and trypsinized and washed with phosphate buffered saline twice. The cells were conjugated with FITC Annexin V (5 μL) stain and propidium iodide stain (5 μL) was administrated in the dark for 15 min. Finally, the labeled cells were analyzed by flow cytometry. The cycle was analyzed using BD Cycletest Plus DNA Reagent Kit (BD Pharmagen) and the apoptosis was measured using FACS Canto II flow cytometry (BD Biosciences).
Western blot assay
Western blot analysis was performed as previously described (Tang et al., 2018). The protein (40 μg) was transferred to the SDS-PAGE and then to the polyvinylidene fluoride membrane. The proteins were incubated using primary and secondary antibodies. The primary antibodies included CDK9 (ab39098, 1:1000; Abcam) and GAPDH (ab8245, 1:1000; Abcam). Finally, the blots were visualized using enhanced chemiluminescence reagents.
Luciferase reporter assay
For the luciferase reporter assay, LINC00152 3′-UTR wild type or mutant type containing complementary binding sites was cloned into the pGL4 plasmid (Promega). miR-193a or control RNA was cotransfected with the LINC00152 wild type and mutant type using the Lipofectamine 2000 transfection reagent. Luciferase ability was normalized to the Renilla by Dual Luciferase Reporter Assay Kit (Promega).
Statistical analysis
All statistical analyses were generated by GraphPad Prism (GraphPad Software Inc., San Diego, CA). The intragroup significance was determined by one-way analysis of variance (ANOVA) and two-tailed Student's t-test. p-Value <0.05 is considered to be statistically significant.
Results
LINC00152 is highly expressed in the AML specimens and cells
To identify whether the lncRNA LINC00152 was ectopically expressed in the AML, RT-PCR was carried out to measure the LINC00152 expression in both clinical specimens and cells. RT-PCR analysis showed that lncRNA LINC00152 was increased in the AML samples compared with the healthy controls (Fig. 1A). However, LINC00152 expression was not significantly different in the FAB subtype of M1–M5 (Fig. 1B). Then, lncRNA LINC00152 was similarly increased in the AML cell lines, especially THP-1 and HL-60 cells (Fig. 1C). Clinical prognosis analysis and survival curves by Kaplan–Meier and log-rank test indicated that the AML patients with high LINC00152 level had poor outcome compared with those with low levels (Fig. 1D). In conclusion, data stated that LINC00152 is highly expressed in the AML specimens and cells, and indicates the poor prognosis.
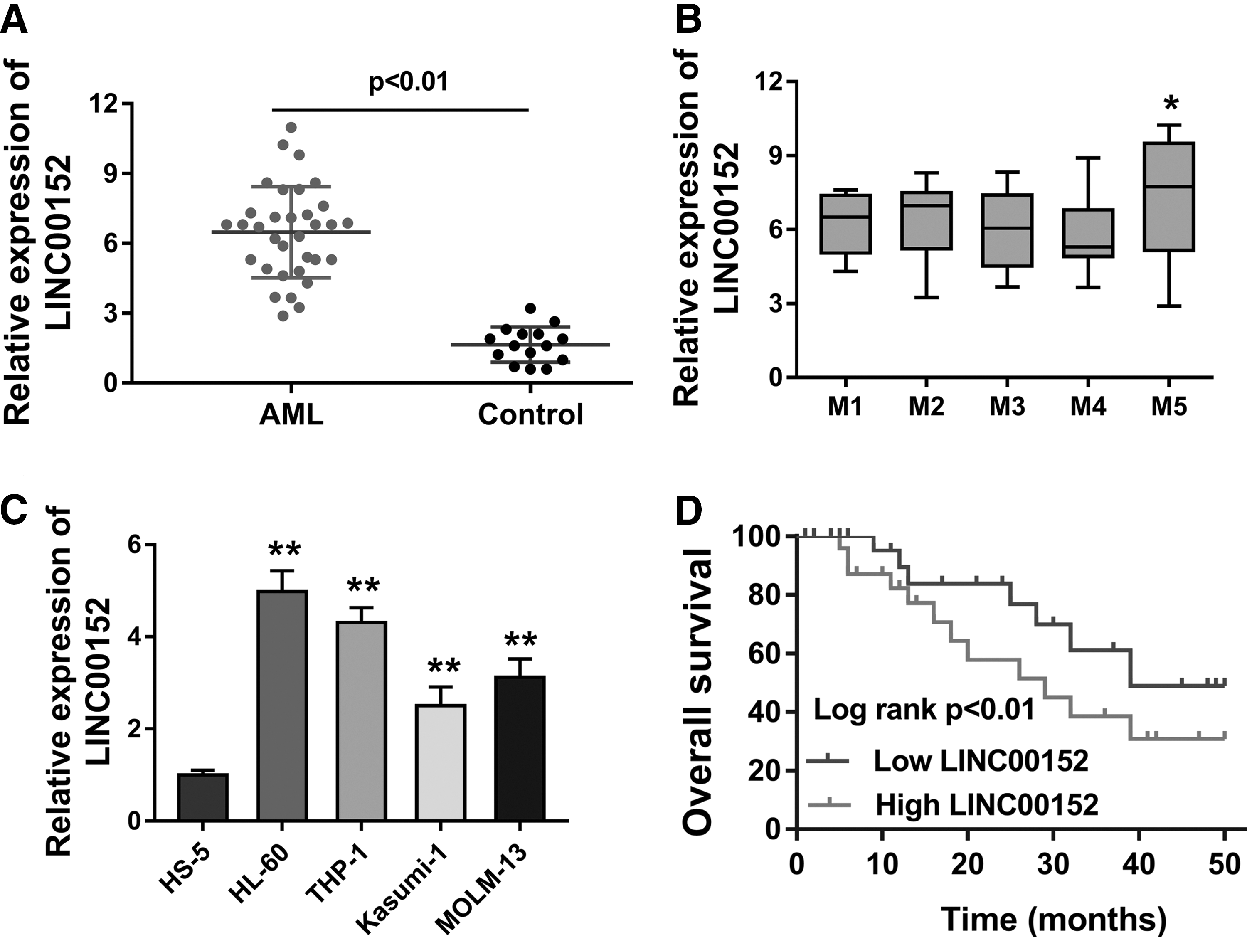
LINC00152 is highly expressed in the AML specimens and cells.
Knockdown of LINC00152 suppresses the proliferation, accelerates the apoptosis, and induces the cycle arrest of AML cells
Because of the overexpression of LINC00152 in the AML tissue and cells, we constructed the loss-of-functional experiments with the siRNA transfection to investigate the roles of LINC00152 (Fig. 2A). The proliferative analysis of AML cells using colony formation assay and CCK-8 assay showed that the knockdown of LINC00152 suppresses the proliferation of AML cells (THP-1 and HL-60) compared with the blank control transfection (Fig. 2B, C). Subsequently, the apoptosis analysis using flow cytometry stated that the knockdown of LINC00152 accelerates the apoptosis of AML cells (THP-1 and HL-60) (Fig. 2D). Furthermore, the cell cycle analysis using flow cytometry stated that the knockdown of LINC00152 impeded the cell progression from G1 to S phase, inducing the cycle arrest at G1/S phase (Fig. 2E). Thus, we determined that knockdown of LINC00152 suppresses the proliferation, accelerates the apoptosis, and induces the cycle arrest of AML cells.

Knockdown of LINC00152 suppresses the proliferation, accelerates the apoptosis, and induces the cycle arrest of AML cells.
LINC00152 positively regulates CDK9 through sponging miR-193a
To determine the deepgoing mechanism that LINC00152 modulates the AML pathogenesis, we try to identify the potential molecular binding of LINC00152. In the THP-1 cells transfected with si-LINC00152, numerous miRNAs were detected and we found that miR-193a had the increasing expression being compared with others (Fig. 3A). Luciferase reporter assay was carried out and validated the molecular binding within miR-193a and LINC00152, revealing the miRNA sponge functions (Fig. 3B). In the AML cells (THP-1, HL-60), miR-193a levels were profoundly decreased, showing the opposite phenotype (Fig. 3C). Interestingly, we again found that miR-193a targeted the 3′-UTR of CDK mRNA (Fig. 3D). Again, in the AML cells (THP-1, HL-60), the miR-193a inhibitor transfection could upregulate the CDK mRNA levels (Fig. 3E). Besides, the si-LINC00152 transfection could increase the miR-193a expression, and decreased the CDK mRNA level (Fig. 3F). So, LINC00152 positively regulates the CDK9 through sponging miR-193a.
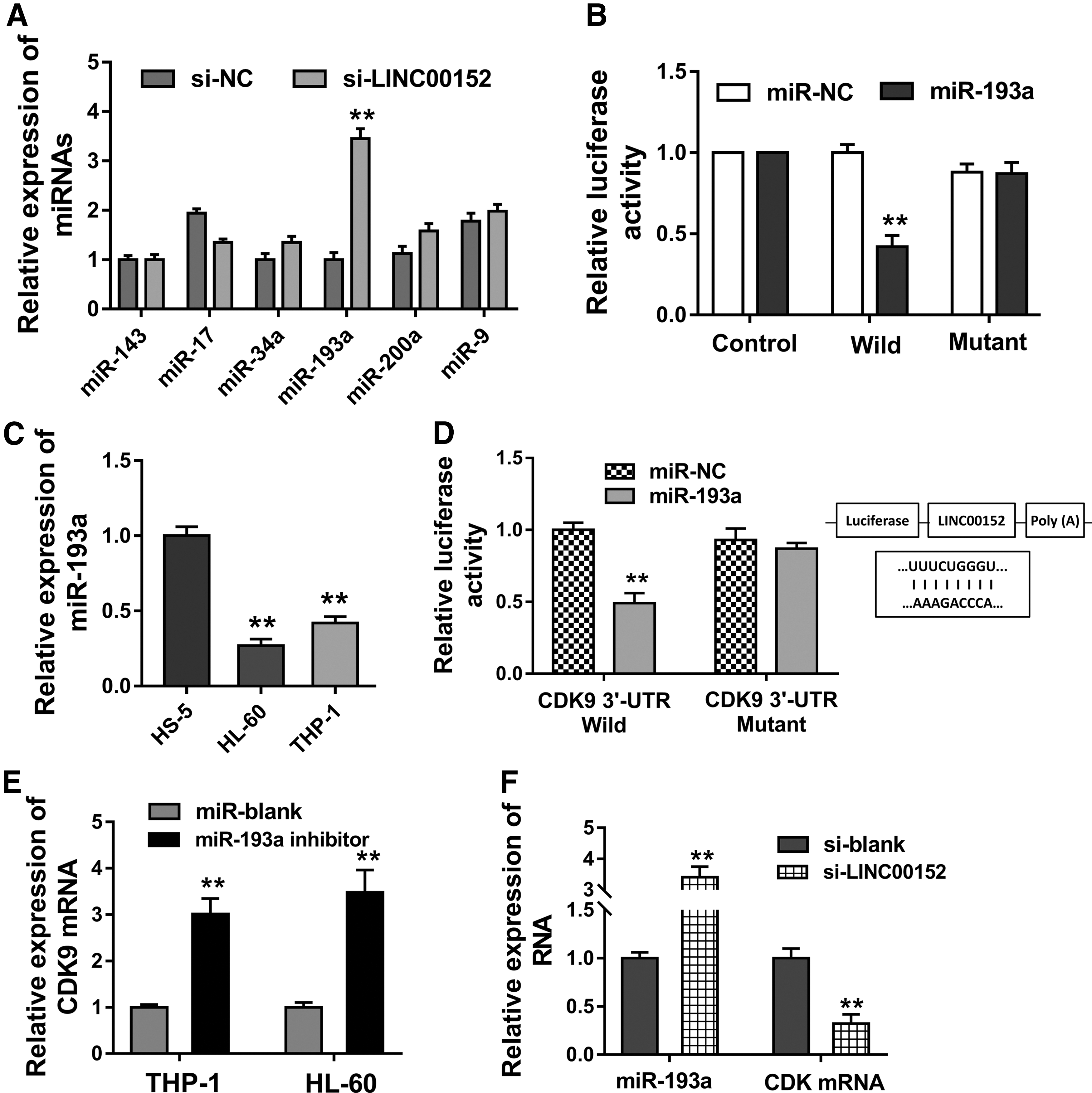
LINC00152 positively regulates the CDK9 through sponging miR-193a.
CDK9 and LINC00152 regulate the AML cells' phenotype
The mentioned data have illustrated the function of LINC00152 on AML's phenotype through miR-193a-CDK9. Then, the rescue experiments were conducted to validate the regulation within LINC00152 and CDK9. Western blot analysis stated that the si-LINC00152 transfection could decrease the CDK9 protein levels (Fig. 4A). Besides, the colony formation assay and CCK-8 assay stated that enhanced CDK9 expression promoted the colonies and proliferative absorbance of AML cells (THP-1) and the si-LINC00152 transfection could decrease them (Fig. 4B, C). Moreover, the apoptotic analysis and cycle analysis by flow cytometer stated that enhanced CDK9 expression promoted the apoptosis and recovered the cycle progression of AML cells (THP-1) and the si-LINC00152 transfection could decrease them (Fig. 4D, E).

CDK9 and LINC00152 regulate the AML cells' phenotype.
Discussion
lncRNA LINC00152 has been identified to be an oncogenic RNA in human cancers, for example, hepatocellular carcinoma, retinoblastoma, and glioma (Li et al., 2018; Ma et al., 2018; Wang et al., 2018b). In this study, we found that the lncRNA LINC00152 is upregulated in the AML and the functions on the AML cells were investigated.
With the rapid development of next-generation sequencing technology and bioinformatics tools, more and more lncRNAs have been discovered in the human tissue, especially the cancers (Chen et al., 2018a; Dong et al., 2018; Guo et al., 2018; Wang et al., 2018a; Wu et al., 2019). For instance, lncRNA RP11-69I8.3 is not significantly different in the AML, but low expressed in acute lymphoblastic leukemia (Si et al., 2018). Moreover, the SNHG5 expression level is overexpressed in the bone marrow and plasma of AML patients and frequently occurred in AML patients with advanced FAB classification (Li and Sun, 2018). For instance, lncRNA ANRIL is upregulated in AML patients, however, it is downregulated after remission, and ANRIL represses AdipoR1 on the glucose metabolism, implying the ANRIL-AdipoR1-AMPK/SIRT1 signaling pathway in regulating cell glucose metabolism (Sun et al., 2018). Similar to these results, our research finds that lncRNA LINC00152 is upregulated in the AML samples and its level is determined to be highly expressed. Besides, the overexpression of LINC00152 is also authenticated in the advanced FAB cooperative group AML patients. And, the overexpression of LINC00152 is closely correlated with the poor outcome of AML patients.
The lncRNA LINC00152 has been identified to be oncogenic RNA in human cancers (Reon et al., 2018). For example, LINC00152 and myeloid cell leukemia-1 (MCL-1) were targeted by miR-125b, which was negatively correlated with the expression of LINC00152 and MCL-1 to regulate the cell proliferation in ovarian cancer (Chen et al., 2018b). In glioma, LINC00152 is found to be significantly upregulated in glioma cells and samples, and LINC00152 knockdown suppresses the glioma cell invasion and proliferation (Chen et al., 2018c). Identically, LINC00152 is found to be upregulated in the AML cells and clinical samples by our data.
Mechanically, LINC00152 is found to be targeted by the miR-193a at the 3′-UTR, acting as an miRNA “sponge.” Besides, the miRNA-193a also targeted the CDK gene 3′-UTR. Luciferase assay and confirmative assay reveal the negative correlation within LINC00152, miR-193a, and CDK9 in the AML cells. In the rescue experiments, we found that CDK9-enhanced expression levels could activate the AML cells tumor behaviors and phenotype in cellular assays. Thus, we could conclude that CDK9 functions as an oncogenic protein for the AML, and moreover, LINC00152 accelerates AML by positively targeting CDK9.
The AML is involved in the gene mutations in the patients (Manchev et al., 2017; Vezzali et al., 2018). The lncRNA ectopically expressed in the AML could significantly regulate the tumor progression. CDK9 has been found to be an oncogenic gene in the human cancer (Opoku-Temeng et al., 2018; Ren et al., 2018; Yu et al., 2018). And, Ajiro et al. (2018) report that CDK9 inhibitor (FIT-039) could inhibit human papillomavirus (HPV) replication of E6/E7 viral oncogenes, restoring tumor suppressors (p53, pRb) in cervical cancer cells (HPV positive) (Ajiro et al., 2018). Thus, the cancer promoting function of CDK9 is confirmed in the AML.
In conclusion, this research has revealed the important role of lncRNA LINC00152 in the AML leukemogenesis through targeting miR-193a/CDK9 axis. This finding could reveal the important pathogenesis of ncRNA and the vital roles of epigenetic regulation.
Footnotes
Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.