
Abstract
O6-Methylguanine-DNA methyltransferase (MGMT) is an enzyme that repairs the DNA damage caused by the tobacco habit, and low activity of this enzyme has been associated with a risk of lung cancer (LC). Our objective was to determine the association of the promoter methylation and the rs12917 polymorphism of MGMT with formation of DNA bulky adducts and the risk of LC in the Mexican Mestizo population. In this study are included 431 subjects. High-resolution melting analysis was used to determine the polymorphism MGMT rs12917 and methylation levels. DNA bulky adducts were determined by 32P-postlabeling. Our results showed that MGMT rs12917 and higher levels of methylation in the MGMT promoter are associated with the risk of LC. The levels of adducts are related with the phe/phe genotype and, only in the cases group, with the hypermethylation (>50%) of MGMT; however, this last association was not statistically significant.
Introduction
Lung cancer (LC) is the second cause of cancer deaths worldwide including Mexico (Ferlay et al., 2015). Factors such as age, gender, genetic background, life style, and ethnicity are associated with the risk of developing this cancer. However, the reported main risk factor associated with the etiology of LC is a tobacco habit, since ∼85% of LC patients are or were smokers (Ferlay et al., 2015).
This carcinogenic potential of tobacco is due because it contains >20 carcinogenic compounds, of these, the tobacco-specific nitrosamine 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK) is most often associated with the etiology of LC (Liu and Gerson, 2006). NNK requires metabolic activation to exert its carcinogenic effects; this depends on the relative efficiency of the enzymes involved in the activation and elimination of the mutagenic metabolites and repair of the DNA damage.
The major metabolic activation pathways of NNK and its main produced metabolite is the 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanol (NNAL), which leads to the formation of two types of DNA methyl adducts, 7-methyguanine and O6-methylguanine (MeG), this being one of the promutagenic DNA adducts that causes [G:C]A:T mutations during DNA replication (Kaina et al., 2007; Ensminger et al., 2014), MeG can be removed only by the enzyme O6-methylguanine-DNA methyltransferase (MGMT) (Pegg, 2000). This enzyme repairs alkylation adducts but irreversibly inactivates MGMT itself in the process. In the absence of active MGMT, MeG forms MeG/T mismatches during replication (Sabharwal and Middleton, 2006; Kaina et al., 2007; Sharma et al., 2009).
The low activity of the MGMT enzyme has been associated with risk to develop different types of cancer, including LC. Interindividual variation in MGMT activity may be attributable to allelic variants in the MGMT gene (Margison et al., 2005). Three common nonsynonymous single nucleotide polymorphisms (SNPs), rs12917, rs2308321, and rs2308327, have been reported in the coding region of MGMT (Kaur et al., 2000; Egyházi et al., 2002). These polymorphisms may affect the primary structure, expression, and DNA repair activity of MGMT (Hill et al., 2005; Pegg et al., 2007; Kokunai et al., 2014). Specifically the C > T polymorphism may affect Zn2+ binding to MGMT, this is known to enhance MGMT activity (Zhang et al., 2003), and also has been associated with increased risk for various cancers (Shen et al., 2005; Han et al., 2006; Hall et al., 2007; Huang et al., 2007), including LC, possibly caused by the formation of adducts, as Molina et al. (2013) demonstrated an association between MGMT84 rs12917 with bulky DNA adduct levels.
In contrast, also aberrant methylation in the promoter region of the MGMT gene is implicated in a mechanism of gene silencing leading to tumorigenesis and has been frequently found in lung and several other tumor types (Esteller et al., 1999; Tranah et al., 2006).
The relationship between germline genetic variations and somatic epigenetic events is an emerging area of investigation. Certainly, germline genetic variants are quite common in the general population, in contrast to heritable germline epimutations that are rare events (Ogino et al., 2007). A possible relationship has been shown between germline SNPs and loss of methylation in an imprinted MGMT gene (Ogino et al., 2007). Recently, a study found in LC a weak association between a promoter SNP and the methylation reporter of the MGMT gene (Leng et al., 2011).
The objective of this study was to determine the relationship between promoter methylation and L84F polymorphism of MGMT with the risk of LC in a sample of Mexican Mestizo population, and whether this polymorphism and the hypermethylation of MGMT could have some relationship with the formation of adducts in the DNA, leading to LC.
Materials and Methods
Retrospective study of cases and controls
We included 431 subjects, 237 controls (without diagnosis of LC) and 194 cases (with primary LC diagnosis).
Subjects with LC diagnosis (cases), who attended in 2011–2014 to the Instituto Nacional de Enfermedades Respiratorias (INER), “Ismael Cosío Villegas” in Mexico City, were evaluated histologically with specimens from lung biopsy or resection; professional pathologists diagnosed the primary LC. These cases were >30 years old (mean age 58.0 ± 13.8 years old).
Control samples (subjects without diagnosis of LC) were collected from blood donors between 2006 and 2007, from the Hospital “20 de Noviembre” in Mexico City (mean age 49.8 ± 14.3 years old).
Cases and controls were unrelated subjects, born in Mexico, with parents and grandparents also born in Mexico. Informed consent was obtained from all participants.
The research protocol was approved by the Bioethics Committees of the Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México, and the INER, “Ismael Cosío Villegas” (Mexico City). The Hospital “20 de Noviembre” gave permission to use in this study the collected buffy coat of blood bank samples.
DNA extraction
DNA samples used for the genotyping analyses were extracted from blood samples collected and stored at −20°C until used. Mononuclear white cells and the genomic DNA were isolated as described by Lahiri and Numberger (1991).
Genotyping
High-resolution melting (HRM) analysis was used to determine the polymorphism MGMT rs12917. The sequences of primers were as follows: 5- CTGCTAATGCCTATTTCC-3 (forward primer) and 5- AACACCGCAGATGGCTTAGT-3 (reverse primer). PCR consisted of initial denaturing at 94°C for 5 min followed by 35 cycles of denaturing at 94°C for 20 s, annealing at 62°C for 20 s and extension at 72°C for 20 s, and a final extension at 72°C for 10 min. HRM analysis was performed using a 80–90°C temperature range, with 0.1°C increments. PCR amplifications using EvaGreen were carried out in 10 μL volumes containing 2 × HRM master mix (contains HotStar Taq Plus DNA Polymerase, EvaGreen dye, 10 μM each primer, and 10 ng template DNA). PCR and subsequent melt curve analysis were carried out using the Rotor-Gene 6000 (Corbett Research). The increase and decrease in fluorescence of EvaGreen during PCR and the melt phase were acquired on the HRM of the Rotor-Gene.
Analysis of methylation
Approximately 1 μg of DNA extracted from mononuclear white cells was sodium bisulfite modified using an EZ DNA Methylation Kit™ (Zymo Research, Orange, CA), according to the manufacturer's instructions. Methylation-specific PCR was performed with primers specific for either methylated or the modified unmethylated DNA. Primer sequences for the methylated reaction were 5′-TTTCGACGTTCGTAGGTTTTCGC-3′ (forward primer) and 5′-GCACTCTTCCGAAAACGAAACG-3′ (reverse primer), and for the unmethylated reaction they were 5′-TTTGTGTTTTGATGTTTGTAGGTTTTTGT-3′ (forward primer) and 5′-AACTCCACACTCTTCCAAAAACAAAACA-3′ (reverse primer). The annealing temperature was 59°C. HRM analysis was performed.
CpGenome universal methylated DNA (Merck millipore) was used as a positive control for methylated MGMT, and CpGenome universal unmethylated DNA (Merck millipore) was used as a control for unmethylated MGMT. Template-free H2O was included as a negative control for the PCR. Standard dilution series of 100%, 75%, 50%, 25%, 10%, and 0% methylation levels were prepared by dilutions of the fully methylated DNA into unmethylated DNA.
The resulting PCR products were extracted from agarose gels using a QIAquick Gel Extraction Kit (Qiagen, Germany) and sequenced with ABI BigDye using an ABI 310 sequencer (Applied Biosystems). The sequences obtained were compared with sequences from NCBI database using BLAST (
DNA adducts analysis
DNA adducts were analyzed by 32P-postlabeling (Phillips et al., 2000). In brief, 4 mg of DNA was digested overnight by micrococcal endonuclease and spleen phosphodiesterase, and enrichment for DNA adducts was obtained by nuclease P1 treatment. Resulting DNA was labeled with 50 mCi 32P-gATP per reaction and DNA adducts were resolved by multidirectional thin layer chromatography (TLC) on 10 cm × 10 cm polyethylenimine (PEI)-cellulose plates. Solvent systems used for TLC were according to Salcido-Neyoy et al. (2015).
Statistical analyses
Epidemiological risk factors and risk of developing LC were examined using Fisher's exact tests. The tests were carried out using GraphPad Prism 5 software.
To explore a possible association between LC risk and the polymorphism, an unconditional logistic regression analyses to calculate the crudes odds ratios (ORs) with 95% confidence intervals, χ2 tests, and Fisher's exact tests were performed in GraphPad-Prism 5. Groups having the lowest risk were used as reference value, assuming a biological activity gradient (codominant model).
Repeated-measures analysis of variance (ANOVA) was used to evaluate the relationship between DNA adducts and tobacco habit, MGMT methylation and rs12917 polymorphism. A criterion of p ≤ 0.05 was used to determine statistical significance. ANOVAs were performed with the Statistical Package for Social Sciences (SPSS-PC; SPSS, Inc., Chicago, IL).
Results
The characteristics of the population included in this study are displayed in Table 1, which shows that cases and controls have a mean age of ∼50 years and for the cases group there are a greater number of women than in the control group.
Characteristics of Healthy Controls and Patients with Lung Cancer
Fisher's exact test.
Significant values.
CI, confidence interval; OR, odds ratio.
With respect to smoking history, a significant difference was found between cases (70.6%) and controls (45.1%), highlighting the importance of the smoking history in the etiology of LC.
In the analysis of association between polymorphisms and the risk of LC (Table 2), we found positive associations with homozygous mutants MGMT T/T (OR = 2.04). As in both groups, gender and smoking habit were statistically different and, therefore, could have influence in the results of associations with the risk of LC, we decided to make an analysis of logistic regression to adjust data with these two variables and then we found that the variant heterozygous was negatively associated (OR = 0.75) and the variant homozygous mutant had a positive relationship with the risk of developing LC (OR = 1.30).
Association of Genetic Polymorphisms with Lung Cancer
Reference value.
Significant values.
With respect to the MGMT promoter methylation, Table 3 shows statistically significant association with the risk of LC when DNA methylation is >50%.
Association of the Percentage of Methylation with the Risk of Lung Cancer
Reference value.
Significant values.
Table 4 shows the association of genotypes and the percentage of the methylation of MGMT promoter with the risk of LC; we found a negative association of heterozygosity with risk of LC when the MGMT gene promoter is 75% and 100% methylated.
Association of Genotypes and Percentage of MGMT Promoter Methylation with the Risk of Lung Cancer
Reference values.
Fisher's exact test.
Significant values.
As for the adduct levels, we separated them according to tobacco habit (Fig. 1) and found higher levels of adducts in smokers; however, this difference was not statistically significant.

Detection of PAH-DNA adducts, according to the Phillips et al., 2000, protocol. Data represent the mean ± SD. White bars correspond to controls, black bars correspond to cases. One-way ANOVA. ANOVA, analysis of variance; PAH, polycyclic aromatic hydrocarbon.
Figure 2 shows that in the control group, the levels of adducts in smokers and nonsmokers are not related to the genotype of the MGMT rs12917 polymorphism. Interestingly, in smokers from cases group, we found there are a statistically significant levels in the T/T genotype in comparison with the C/C and C/T genotypes.

Association of genotypes of MGMT rs12917 in smokers and nonsmokers with PAH-DNA adduct levels. One-way ANOVA.
In contrast, the levels of adducts in relation to methylation showed higher levels when the subjects were smokers and showed hypermethylation of the MGMT gene; however, there were no statistically significant differences between cases and controls (Fig. 3).
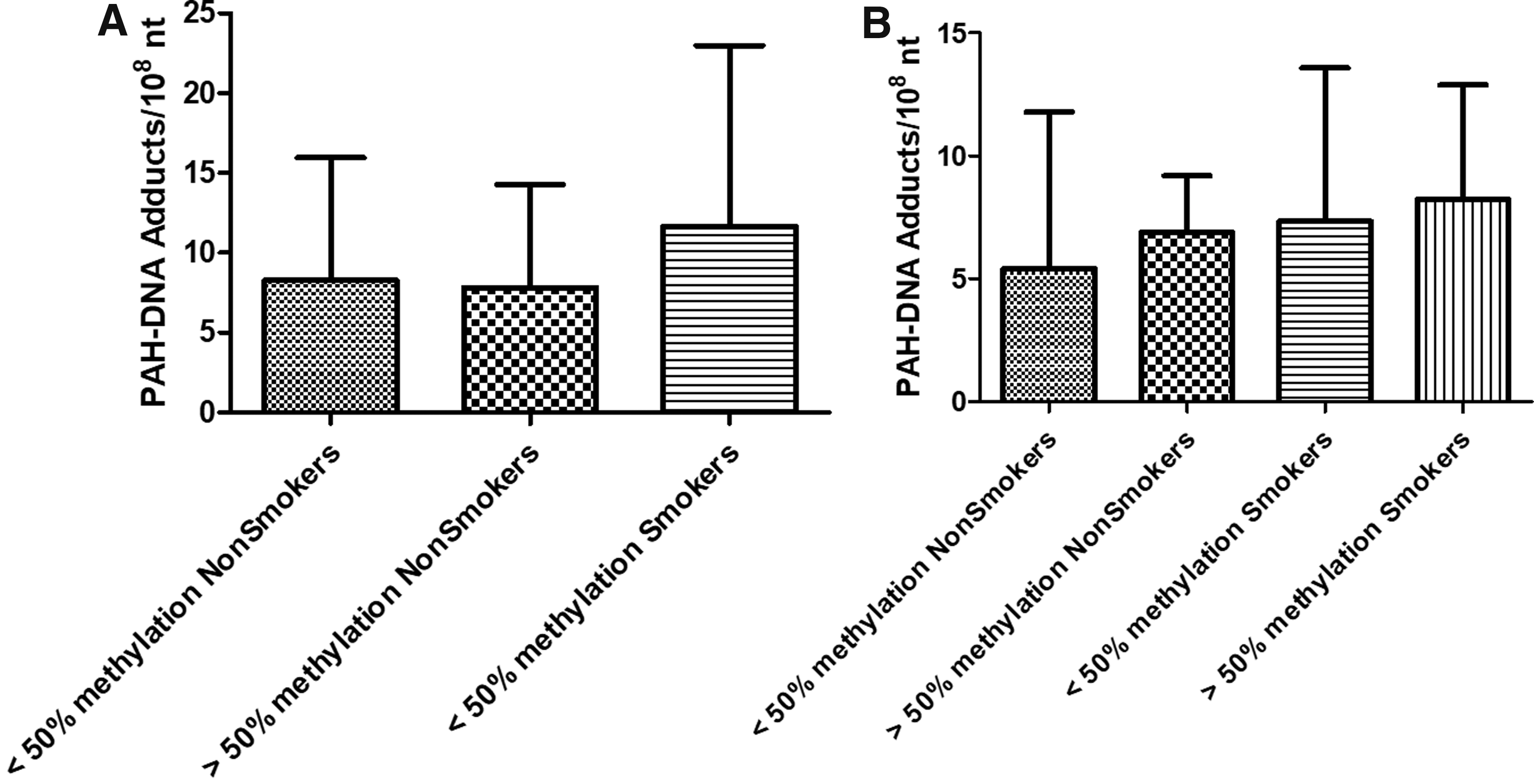
Association of percentage of MGMT promoter methylation in controls
Discussion
This study included 431 subjects, 194 cases and 237 controls, and, as reported in many other studies, we also found a very clear relationship between LC and smoking habit, as our cases group had significantly greater number of smokers than the control group, showing a risk near to two times greater to develop LC in smokers compared with nonsmokers (Subramanian and Govindan, 2007; Thompson et al., 2012).
We found that homozygous mutants MGMT T/T are associated with risk of LC; this could be related to DNA repair capability and enzymatic activity, thereby leading to the risk for breast cancer and LC. (Lu et al., 2015).When we adjust the data by gender and smoking habit, we find a negative association with heterozygous genotypes, this association could be explained based on the reports that propose that the heterozygocity has a significant effect in reducing the risk for developing some diseases, such as cancer; this effect is termed “Heterozygous advantage,” which refers that the heterozygous genotype having a copy of each allele has several advantages over the wild-type homozygous, and the nonwild homozygous type genotypes have not (Sellis et al., 2011).
In reference to the association of rs12917 polymorphism and the percentage of methylation of MGMT, it is important to highlight that the relationship between allelic variants and somatic epigenetic events has been an emerging area of investigation. Certainly, genetic variants are quite common in the general population, in contrast to epigenetics marks. In reports preview, a possible relationship between allelic variants and methylation in some genes (IGF2 and MLH1) has been shown, without statistical significance. However, there has been no compelling evidence for the relationship between a polymorphism and promoter methylation (and silencing) of the same gene (Murrell et al., 2004; Hitchins et al., 2007).
MGMT promoter methylation is very common in LC; however, the results about the association between the level of MGMT methylation and LC were different in our study as we found positive association between MGMT and LC in the Mexican Mestizo population.
On the other hand, the MGMT gene dysfunction due to the promoter hypermethylation could generate DNA damage through the formation of adducts. We determined the formation of adducts in the cases group and in control group and we separated both groups in smokers and nonsmokers, this is because Molina et al. (2013) showed that the adducts are significantly related to the tobacco habit.
We found higher levels of adducts in smoker patients with LC with the MGMT Phe/Phe genotype, these results agree with what was previously reported in healthy individuals and reaffirm the relationship of the MGMT rs12917 (C>T) polymorphism with increased mutation frequency in the lymphocytes of smokers compared with smokers carrying the wild-type allele. In contrast, we found higher levels of adducts in smoking cases and control groups, regardless of the MGMT methylation percentage.
This may be due to the aberrant methylation of promoter regions in DNA repair genes, which is a key event in the formation and progression of cancer and epigenetic change of CpG islands in the gene promoter region (Zhang et al., 2003; Gu et al., 2013).
Conclusions
In this study, we provide evidence that in a sample of Mexican Mestizo population, the MGMT gene is associated with the etiology of LC because there is a positive association with the rs12917 polymorphism and the hypermethylation of gene promoter the MGMT with the risk of LC. In addition, the levels of adducts were related with the T/T genotype and hypermethylation (>50%) of MGMT in cases, this last association was not statistically significant.
Our results contribute to explain the high frequency of LC in the Mexican Mestizo population; however, this information must be verified in future studies with a greater number of samples to corroborate the association of methylation with DNA bulky adduct levels.
Footnotes
Acknowledgments
This study was supported, in part, by the Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica (PAPIIT, IN210111, and IN208314 to J.R.), Programa para el Desarrollo Profesional Docente (PRODEP, DSA/103.5/15/3073 to O.C.M.-R.), and Consejo Nacional de Ciencia y Tecnología (CONACYT, CB-2012/178860 to J.R.). We thank all our blood donors for their valuable contribution to this project and David Rico for computer support.
Ethical Approval
This work was performed under the approval of the Ethics Committees of Instituto de Investigaciones Biomédicas, UNAM, and Instituto Nacional de Cancerología (INCan).
Authors' Contributions
Study conception and design were carried out by O.C.M.-R., P.P., M.E.G., C.C.-H., and J.R. Support for access to biological samples from patients was provided by M.V.-F. Analysis and interpretation of data were carried out by O.C.M.-R., R.P.-M., and J.R. Drafting of the article was performed by O.C.M.-R. and J.R. Critical revision of the article was carried out by R.P.-M., M.E.G., V.P.L.-S., C.C.-H., L.C.-Á., M.V.-F., and J.R.
Disclosure Statement
No competing financial interests exist.