
Abstract
Staging and pathological grading systems are convenient, but imperfect predictors of recurrence of head and neck squamous cell carcinoma. Therefore, to identify potential alternative prognostic markers, we investigated the methylation status of the promoter of Sal-like protein 2 (SALL2). SALL2 mRNA expression was absent in 8/9 (88.9%) University of Michigan squamous cell carcinoma cell lines, whereas two nonmalignant cell lines had stable expression. The normalized methylation value of SALL2 in cancer cell lines was significantly higher than in normal cell lines. SALL2 methylation found in 74 of 233 (31.8%) tumor specimens was correlated with the methylation status of both SALL1 and SALL3. SALL2 methylation was not associated with any difference in disease-free survival (DFS). Therefore, the presence of SALL2 methylation was statistically correlated with a decrease in DFS in patients with oral cancer (log-rank test, p = 0.032). Furthermore, it was associated with disease recurrence in 36.2% of oral cancer cases, with an odds ratio of 2.922 (95% confidence interval = 1.198–7.130; p = 0.018) by multivariate Cox proportional hazard regression analysis. This study suggests that cytosine-phosphate- guanosine (CpG) hypermethylation is a likely mechanism of SALL2 inactivation and supports the hypothesis that SALL2 could serve as an important clinical risk assessment.
Introduction
More than 90% of head and neck cancers are squamous cell carcinomas (HNSCC) that arise in mucosal surfaces of the hypopharynx, larynx, oropharynx, and oral cavity. (Parkin et al., 2005). Major risk factors for HNSCC include sex, tobacco smoking, alcohol consumption, and human papillomavirus (HPV) infection (Kumar et al., 2008). Over the past 30 years, there has been a decline in carcinomas of the hypopharynx, larynx, and oral cavity (Parkin et al., 2005) and a simultaneous increase in the incidence of oropharynx cancer (Whang et al., 2015). Oral cancer is often first diagnosed at late stages of the disease (advanced regional disease and/or metastasis) (Dionne et al., 2015). To improve the survival rate of HNSCC patients, it is necessary to identify reliable markers that provide predictive prognostic power for effective clinical management of the disease.
The Spalt (Sal) genes were first isolated in Drosophila. Mice and humans have four known Sal-related genes, Sall1 to Sall4 and SALL1 to SALL4, respectively (Karantzali et al., 2011). Sal-like protein 2 (SALL2) has been mapped to 14q11.2, and, since its identification, studies have found that it plays an essential role during embryonic development. There is increasing evidence of the association between SALL2 and human cancer; however, many aspects of SALL2 regulation, targets, and function remain unknown (Sung and Yim, 2015; Hermosilla et al., 2017). Recent studies have investigated the association between SALL2 expression and carcinogenesis: one group demonstrated that SALL2 upregulates P16 transcription through a SALL2-responsive element with a SALL2-binding site near the proximal region of the P16 promoter (Sha et al., 2015); a report showed that SALL2 methylation reduced the level of SALL2 mRNA in esophageal carcinoma (Luo et al., 2017); and another report found that aberrant hypermethylation of SALL2 contributes to carcinogenesis in ovarian cancer (Sung et al., 2013).
Previous reports from our group noted that expression of SALL1 and SALL3 mRNA is lost in HNSCC as a result of DNA methylation (Misawa et al., 2017; Misawa et al., 2018a). Other reports showed that aberrant hypermethylation of SALL1 was found to contribute to carcinogenesis in breast carcinoma (Hill et al., 2010) and chronic lymphocytic leukemia (Tong et al., 2010). SALL3 hypermethylation reduced the level of SALL3 mRNA in hepatocellular carcinoma (Yang et al., 2012), and aberrant hypermethylation of SALL3 was found to contribute to carcinogenesis in cervical cancer (Wei et al., 2015). Although results have been obtained for esophageal carcinoma and ovarian cancer, the effect of hypermethylation of SALL2 in HNSCC remains to be explored.
SALL2 methylation has been linked to loss of gene expression, and we speculated that this is a critical event in the development of oral cancer. To test this hypothesis, we investigated the site-specific methylation status of SALL2 in HNSCC at diagnosis and during follow-up to assess its clinical significance and potential as a prognostic marker for tumor recurrence and patient survival.
Materials and Methods
Tumor samples and cell lines
We examined 233 tumor samples, namely from the hypopharynx (n = 55), larynx (n = 44), oropharynx (n = 65), and oral cavity (n = 69), and 36 noncancerous mucosal samples. Primary HNSCC specimens were collected at surgery in the Hamamatsu University Hospital, Japan. Medical record information was obtained from clinical records. The mean age of the patients (male, 200; female, 33) was 65.4 years (range, 32–92 years). Written informed consent was obtained from each subject, and the study procedures were approved by the Institutional Review Board at the Hamamatsu University School of Medicine (16-072). DNA and cDNA from University of Michigan squamous cell carcinomas cell lines (UM-SCC), 81F fibroblasts, and HOK-16B were provided by Dr. Thomas E. Carey of the University of Michigan (Supplementary Table S1). Noncancer tonsillar specimens, Ton7, Ton8, and Ton10, were surgically obtained from patients with chronic tonsillitis.
Bisulfite modification and quantitative methylation-specific polymerase chain reaction
DNA was extracted from fresh-frozen tissue using QIAamp DNA Mini Kit (Qiagen, Hilden, Germany), following the manufacturer's protocol. Conversion of genomic DNA using sodium bisulfite was performed with the MethylEasy™ Xceed Rapid DNA Bisulphite Modification Kit (Human Genetic Signatures Pty Ltd., Sydney, Australia). The primer sequences and base pairs are listed in Supplementary Table S2. A standard curve was generated using serial dilutions of EpiScope Methylated HeLa genomic DNA (Takara Bio), used as the control. The normalized methylation value (NMV) was defined as follows: NMV = (SALL2-S/SALL2-FM)/(ACTB-S/ACTB-FM). SALL2-S and SALL2-FM represent SALL2 methylation levels in a sample and universally methylated DNA, respectively. ACTB-S and ACTB-FM correspond to β-actin (ACTB) in the sample and the universally methylated DNA, respectively (Misawa et al., 2014). To analyze the methylation status of SALL1 (Misawa et al., 2018a) and SALL3 (Misawa et al., 2017), primers and conditions were used as previously described.
RNA extraction, quantitative Reverse Transcription PCR, and reactivation of SALL2 expression
Total RNA was extracted from fresh-frozen tissue with RNeasy Plus Mini kit according to the manufacturer's instructions (Qiagen). cDNA was synthesized from total RNA using the ReverTra Ace qPCR RT kit (Toyobo, Osaka, Japan). Primer sequences and base pairs are shown in Supplementary Table S2. SALL2 mRNA expression was compared between samples by normalization to glyceraldehyde 2-phosphate dehydrogenase mRNA expression. At 1 h after plating, cultures were incubated for 48 h with 5-azacytidine (15 Lg/mL; Sigma-Aldrich, St. Louis) for reactivation of SALL2 expression.
Analysis of HPV status
Evaluation of HPV infection was performed by amplifying through the HPV Typing Set (Takara Bio), a polymerase chain reaction (PCR) primer set specifically designed to identify HPV genotypes 16, 18, 31, 33, 35, 52, and 58 in genomic DNA. The PCR HPV typing was performed according to the manufacturer's instructions.
The cancer genome atlas data
Aberrant DNA methylation of SALL2, SALL1, and SALL3 from The cancer genome atlas (TCGA) (accessed October 2018) was collected from MethHC (
Data analysis and statistics
We generated receiver operator characteristic (ROC) curve analysis of SALL2 genes in 36 matched paired tumor and noncancerous mucosal samples obtained from patients with HNSCC; these included 9 hypopharynx, 7 larynx, 9 oral cavity, and 11 oropharynx tumor samples, using the Stata/SE 23.0 system (Stata Corporation). The sensitivity, specificity, and area under the curve of different cutoff values of absolute and relative presence were calculated to distinguish between the methylation levels in normal and HNSCC tissues, and the NMV thresholds were calculated for each target gene. The frequency of SALL2 methylation was determined by choosing a specific cutoff value.
Associations between clinical variables were assessed using Student's t-test. Disease-free survival (DFS) was measured from the date of the initial treatment to the date of diagnosis of locoregional recurrence or distant metastasis. The differences between groups were illustrated by drawing survival curves obtained by the Kaplan-Meier method, and the log-rank test was used to compare survival rates. DNA methylation was also examined within multivariate Cox proportional hazards regression models adjusted for age (≥65 vs. <65 years), HPV status, smoking status, alcohol intake, and tumor stage (I–III versus IV). P-values of less than 0.05 were considered to indicate statistical significance. All statistical analyses were performed using of StatMate IV software (ATMS Co., Ltd.).
Results
Initial screening: 36 matched paired head and neck tumor samples and corresponding adjacent noncancerous mucosal samples.
SALL2 CpG islands and regions analyzed by quantitative methylation-specific polymerase chain reaction (qMSP) are shown in Figure 1A. First, the methylation status of the SALL2 promoter was analyzed in 36 matched paired cancerous and corresponding noncancerous mucosae. Promoter methylation levels were represented by NMVs, that is, the ratio of methylated DNA at the target sequence in each specimen to a fully methylated control DNA. The SALL2 methylation levels were significantly higher in primary HNSCCs than in noncancerous mucosal tissues (median NMV = 0.131 versus 0.019, p = 0.008; paired Student's t-test; Fig. 1B). SALL2 methylation exhibited highly discriminative ROC curve profiles, which clearly distinguished HNSCCs from normal mucosal tissues (area under the ROC curve = 0.6397). At the cutoff value of 0.041, the sensitivity and specificity were 47.2% and 91.7%, respectively (Fig. 1C).
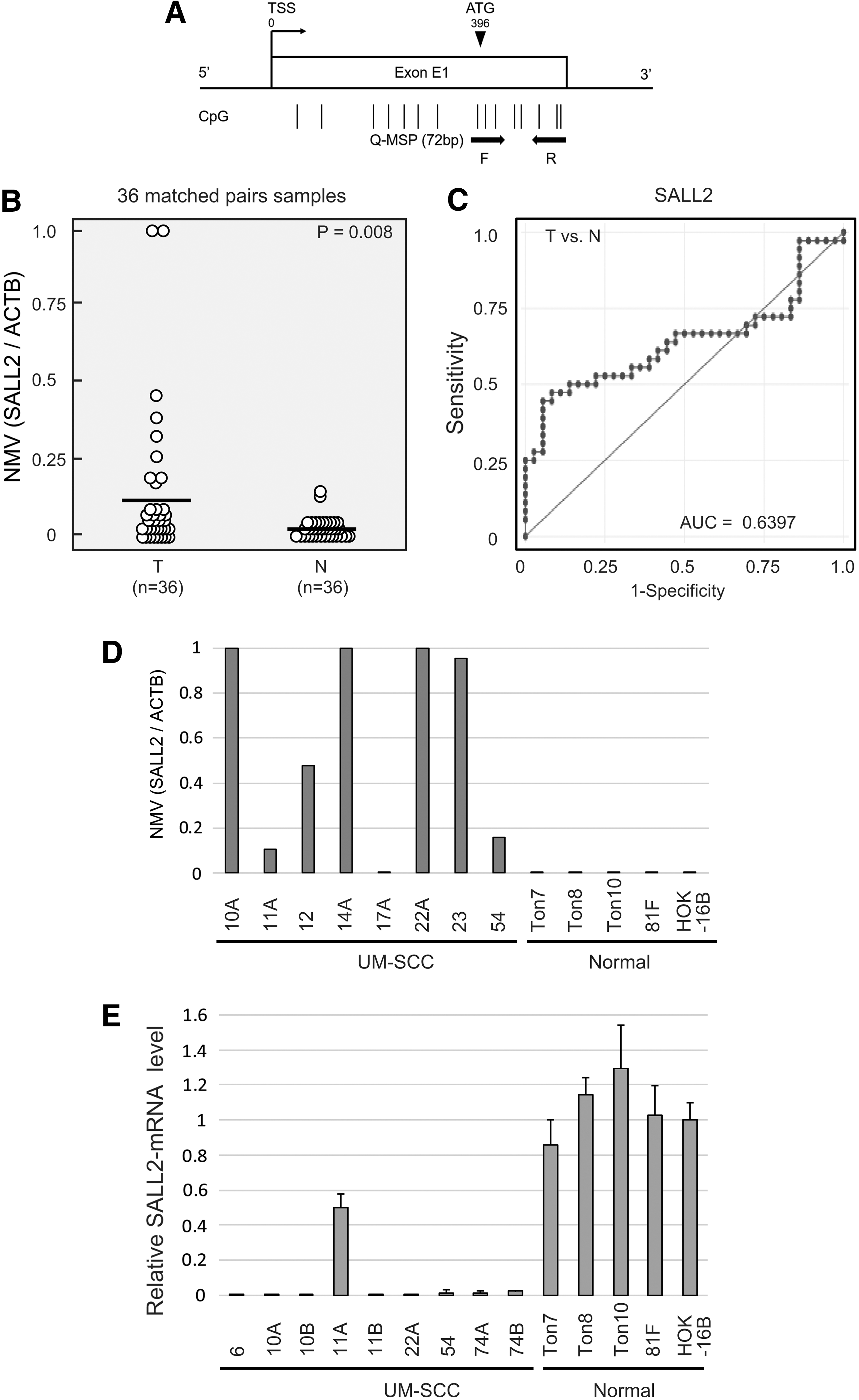
Matched pairs of HNSCC tumors and adjacent normal mucosal tissues in HNSCC patients, and SALL2 methylation and expression in UM-SCC cell lines, noncancer tonsillar tissues, fibroblasts, and keratinocyte cell lines.
SALL2 methylation and expression analysis in cancer cell lines
qMSP results indicated a significantly increased NMV of SALL2 methylation in cancer cell lines compared to normal tonsil tissues and normal cell lines (Fig. 1D). All UM-SCCs showed reduced SALL2 mRNA expression. In contrast, 81F fibroblasts, HOK-16B, Ton7, Ton8, and Ton10 displayed a stable expression of SALL2 mRNA (Fig. 1E). UM-SCC-54 cell lines, which exhibited absence of SALL2 expression and hypermethylation, were cultured with 5-azacytidine. The results show that SALL2 expression was upregulated after treatment (Supplementary Fig. S1).
SALL2 promoter methylation correlates with clinicopathologic parameters in 233 HNSCC samples
The SALL2 promoter was methylated in 74/233 (31.8%) of the cases and unmethylated in 159/233 (68.2%) of the cases (Supplementary Fig. S2). Age, sex, smoking status, alcohol intake, tumor size, lymph node status, clinical stage, recurrence events, and HPV status were not associated with SALL2 methylation status (Table 1). However, SALL2 methylation significantly correlated with SALL1 and SALL3 methylation status (p < 0.001 and p = 0.019, respectively; Table 1). Next, the association between the site-specific methylation frequency, namely in 55 hypopharynx, 44 larynx, 65 oropharynx, and 69 oral cavity tumor samples, of SALL2 genes and the clinicopathological features of the patients is summarized in Table 2. Among oropharyngeal cancers, SALL2 methylation was significantly correlated with sex (p = 0.011) and alcohol exposure (p = 0.013). In patients with oral cancers, SALL2 methylation was significantly correlated with age (p = 0.030) and recurrence (p = 0.027) (Table 2).
Sal-Like Protein 2 Gene Methylation Status in Primary Samples of Head and Neck Squamous Cell Carcinoma in 233 Patients
Fisher's exact test.
p < 0.05.
HPV, human papillomavirus.
Distribution of Sal-Like Protein 2 Methylation Status by Selected Epidemiologic and Clinical Characteristics
Fisher's exact probability test.
p < 0.05.
SALL2, Sal-like protein 2.
Prognostic value of the methylation status of SALL2 promoters
Kaplan-Meier plots indicated that methylation of SALL2 in the tumors was related to the DFS. Survival of the 233 HNSCC patients with (methylation) and without (unmethylation) SALL2 methylation (log-rank test, p = 0.111) is shown in Figure 2A. The SALL2 methylation status in patients with hypopharyngeal cancers, laryngeal cancers, or oropharyngeal cancers was not related to outcome (Fig. 2B–D). Among the patients with oral cancers, the rate of DFS of patients exhibiting SALL2 methylation was 43.8% compared with the 66.8% in the group with unmethylated SALL2 (log-rank test, p = 0.032) (Fig. 2E). Joint analysis of the three genes (SALL2, SALL1, and SALL3) showed a trend of significant decrease in DFS as the number of methylated genes increased (Fig. 3A–C).
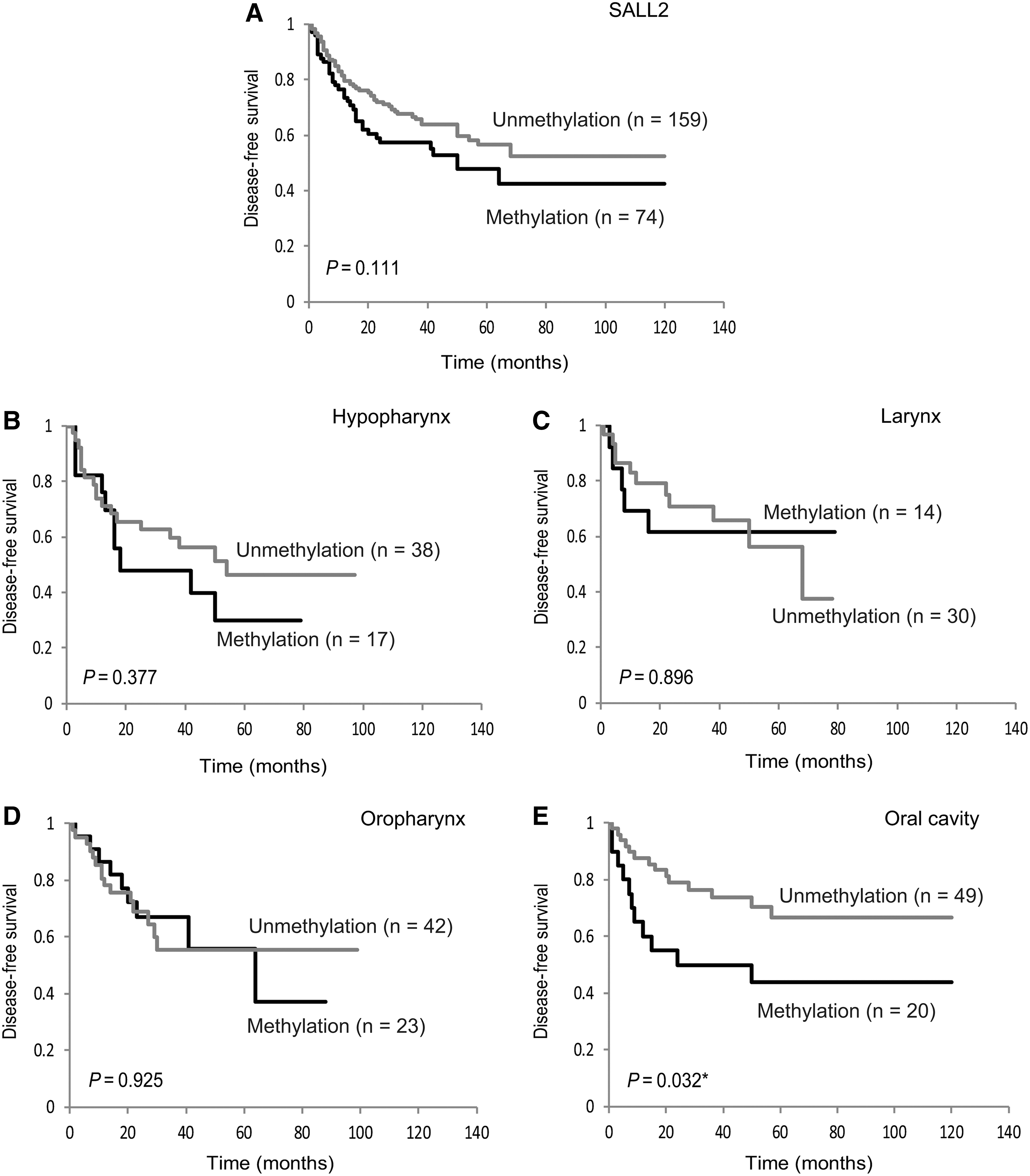
Kaplan-Meier survival curves for HNSCC patients.
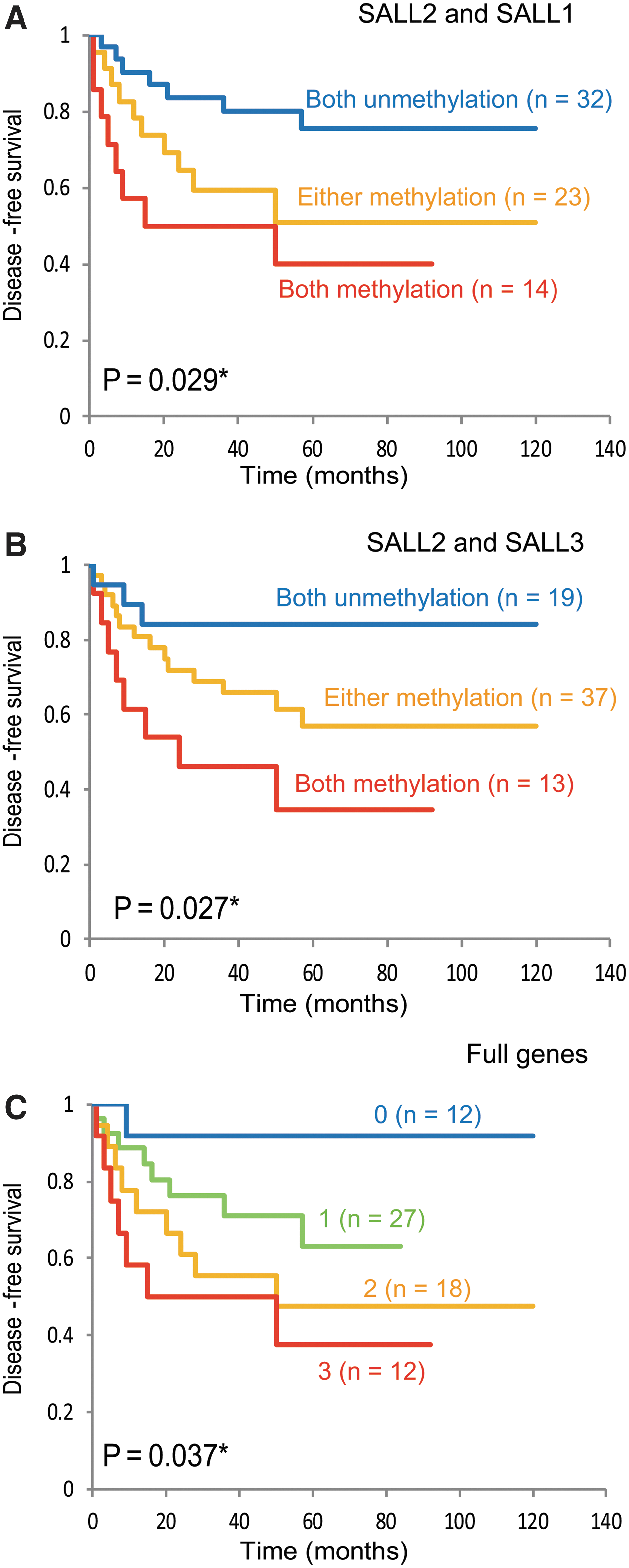
Kaplan-Meier survival curves for patients with oral cavity cancer. A joint analysis of the three genes showed a significant trend for decreased DFS as the number of methylated genes increased. DFS for HNSCC patients with relation to
In addition, results of the multivariate Cox proportional hazard regression analysis, which included age, HPV status, smoking status, alcohol consumption, and tumor stage, indicated that survival rates were not associated with SALL2 methylation: the adjusted odds ratio (OR) for recurrence was 1.39 (95% confidence interval [CI]: 0.91–2.11; p = 0.127) (Fig. 4). However, SALL2 methylation analysis in 69 oral cancer patients was associated with an OR for recurrence of 2.922 (95% CI = 1.198–7.130; p = 0.018). Specifically, these results indicate that SALL2 methylation is a predictor of poor outcome in oral cancer patients (Fig. 4).
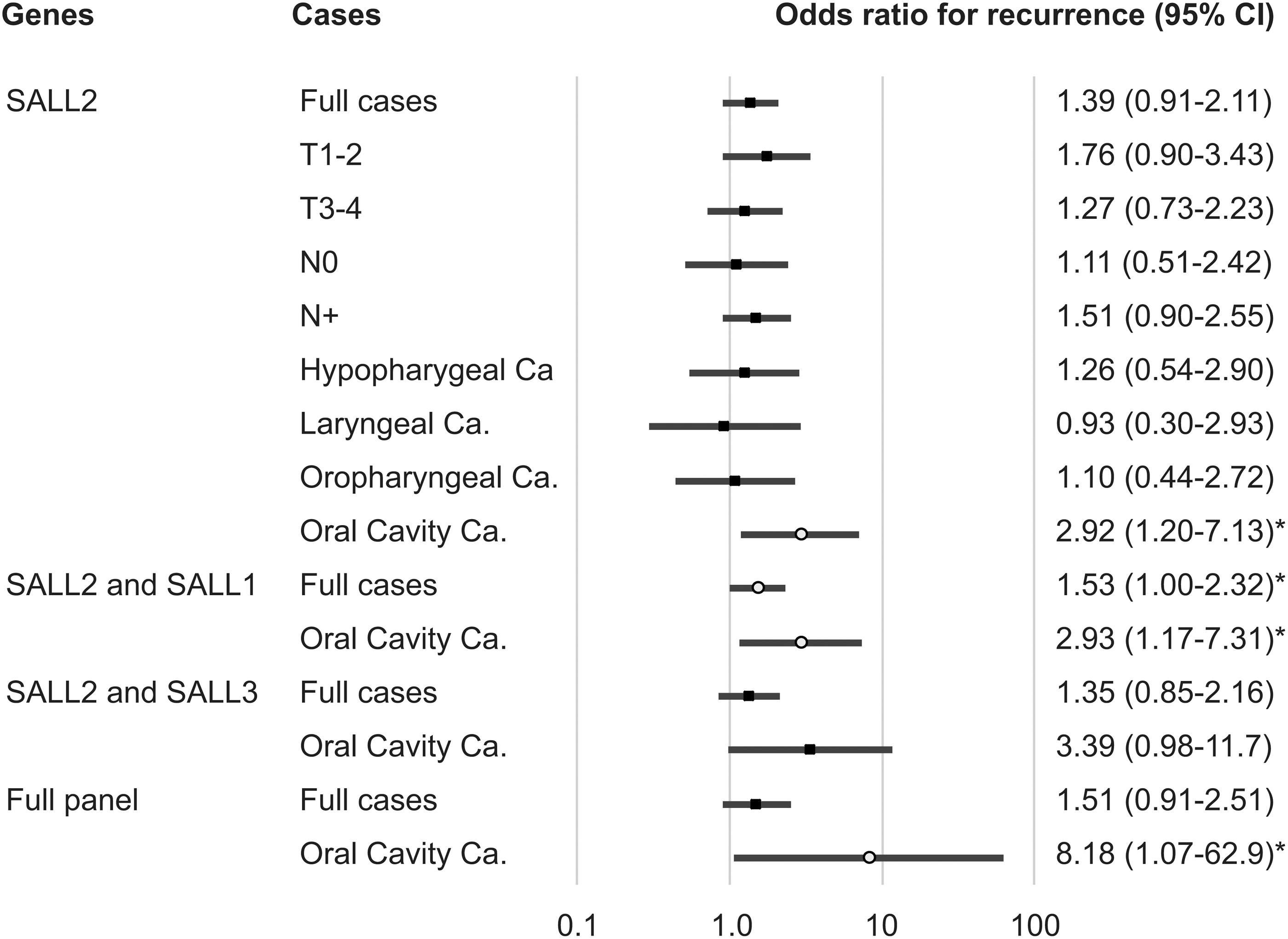
Odds ratios for recurrence based on the Cox proportional hazards model adjusted for age (≥65 years vs. <65 years), HPV status, smoking status, alcohol exposure, and tumor stage (I, II, and III vs. IV). Cox proportional hazards model, revealing the estimated odds of recurrence associated with SALL2, SALL1, and SALL3 methylation. CI, confidence interval. *p < 0.05. HPV, human papillomavirus.
External validation of TCGA data
Publicly available HNSCC data from TCGA were examined for SALL2 DNA methylation and expression (Supplementary Fig. S3). SALL2 methylation in the HNSCC TCGA cohort had an average beta value of 0.405, compared to the 0.319 in normal samples (p < 0.001, Student's t-test). The SALL2 promoter was hypermethylated in samples from eight tumor types originated from the bladder, mammary gland, colon, lung, pancreas, prostate, and rectum compared to normal samples (Supplementary Fig. S4A). Moreover, using mRNA expression data, SALL3 mRNA expression was significantly associated with sex and smoking status (p = 0.009 and p = 0.001, respectively) (Supplementary Table S3).
Discussion
Early diagnosis and treatment of HNSCC are essential. A clinical risk assessment that can improve the survival rate is urgently needed to ensure optimal treatment of HNSCC patients (Dasgupta et al., 2012). We investigated SALL2 methylation profiles in 233 HNSCC patient tissues. This study demonstrated that SALL2 promoter methylation was associated with transcriptional inhibition and was independently associated with aggressive tumor behavior in oral cancer, suggesting its role in tumor suppression and potential to serve as a clinical risk assessment in oral cancer patients.
Three other SALL genes have been identified, namely SALL1 on chromosome 16q12.1, SALL3 on chromosome 18q23, and SALL4 on chromosome 20q13.2 (Parrish et al., 2004). SALL1 expression is associated with the expression levels of several genes involved in epithelial–mesenchymal transition and with survival in breast cancer patients (Wolf et al., 2014). Its hypermethylation has been reported in several tumors such as non-small cell lung cancer, prostate cancer, colon cancer, and acute lymphocytic leukemia (Hill et al., 2010, Tong et al., 2010). Previously, our group suggested that SALL1 acts as a tumor suppressor gene and can serve as a prognostic biomarker for early T1 and T2 HNSCC (Misawa et al., 2018b). Hypermethylation of the promoter region of SALL3 is, together with HPV infection, reportedly involved in cervical cancer: the expression levels of SALL3 mRNA were found to be downregulated in cervical cancer (Wei et al., 2015). Another report showed SALL3 was silenced by DNA methylation and that SALL3 interacts with DNA methyltransferase 3 alpha (DNMT3A) in hepatocellular carcinoma (Shikauchi et al., 2009). Our group reported that the transcriptional inactivation of SALL3 was associated with aberrant methylation of other tumor-related genes, as well as with TET1, TET2, and DNMT3A levels, in HNSCC (Misawa et al., 2017). Finally, SALL4, an important stem cell regulator, is expressed in hematopoietic stem cells and various malignancies (Yang, 2018). Upregulation of SALL4 causes myelodysplastic syndrome, lung cancer, gastric cancer, and hepatocellular cancer (Lin et al., 2013; Wang et al., 2017; Liu et al., 2018; Pan et al., 2018; Zhang et al., 2018). Findings suggest that SALL4 can be increased in several cancers, worsening the prognosis (Nicole et al., 2017).
Clarifying the epigenetic regulation of SALL2 can reveal the mechanism of tumorigenesis and disease recurrence in various types of tumor (Sung and Yim, 2017). This epigenetic modification was confirmed in many primary tumors and correlated with negative SALL2 expression in esophageal cancer and ovarian cancer (Sung et al., 2013; Luo et al., 2017). Other studies indicate that loss or reduction of SALL2 expression may be involved in leukemogenesis (Chai, 2011) and breast cancer (Liu et al., 2007). On the other hand, SALL2 is found upregulated in Wilms tumor (Li et al., 2002), synovial sarcoma (Nielsen et al., 2003), tongue cancer (Estilo et al., 2009), and testicular cancer (Alagaratnam et al., 2011), indicating that the role of SALL2 in cancer may depend on tumor type.
Interestingly, recent studies support the role of SALL2 as a tumor suppressor (Miao et al., 2017). Its effect on cell cycle progression is associated with the transcriptional repression of cyclins D1 and E1 (V et al., 2018). SALL2 transcriptionally increases the expression of cyclin-dependent kinase inhibitors P21 and P16 and represses the proto-oncogene c-Myc (Parroche et al., 2011; Wu et al., 2015). It inhibits cell proliferation by repressing G1 to S phase cell cycle transition (V et al., 2018). Similarly, SALL2, which positively regulates P21 transcription, binds to viral oncogenic proteins, including HPV16 E6 (Parroche et al., 2011). Evidence of an association of SALL2 with human cancer is increasing; therefore, SALL2 expression might be regulated by tissue-specific transcription factors and recruited by different binding partners with different activities, depending on the cellular context and conditions (Sung and Yim, 2017). In a similar manner, the influence of various stress conditions, age, sex, and the consumption of alcohol and tobacco, may operate by impacting a variety of signaling molecules that may be capable of inducing specific changes in SALL2 methylation.
Conclusions
This study suggests that SALL2 hypermethylation may play a role in HNSCC tumorigenesis and serve as an important clinical risk assessment. Indeed, we found that the SALL2 methylation was increased in tumor tissues compared to the corresponding noncancerous tissues. The SALL2 methylation was associated with disease recurrence in oral cancer patients. Further analyses of SALL2 will help define its biologic role in head and neck carcinogenesis and its utility as a marker for early detection and prognosis.
Ethics Approval and Consent to Participate
The tissue samples were obtained with informed written consent following protocols approved by the research ethics board of the Hamamatsu University School of Medicine.
Footnotes
Acknowledgments
The authors would like to thank Ms. Yuko Mohri for her excellent technical support. Funding: this work was supported by Grants-in-Aid for Scientific Research (no. 16K11228, 16K20239, 17K11380, 17K16903, and 17K16904) from the Ministry of Education, Culture, Sports, Science, and Technology of Japan. Funders had no role in study design, data collection, analysis, or publication.
Authors' Contributions
K.M. designed the experiments. A.I., Y.M., and K.M. conducted all experiments. A.I. and K.M. wrote the article. D.M., T.K., S.E., M.M., S.Y., T.K., and H.K. performed the data analysis and discussed the results. All authors read and approved the final article.
Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.