
Abstract
Increasing evidence has suggested the key roles of miRNAs in the initiation and progression of human cancers. miR-383 was downregulated and played a suppressive role in a variety of cancers; however, the function of miR-383 in gastric cancer remains unclear. In this study, we found that the expression of miR-383 was significantly reduced in gastric cancer tissues and correlated with the advanced progression of these cancer patients. Functional analysis showed that overexpression of miR-383 inhibited the proliferation and upregulated the apoptosis of gastric cancer cells. Furthermore, cyclin E2 was predicted as the target of miR-383 using the bioinformatics database. miR-383 bound the 3′-untranslated region of cyclin E2 and decreased the expression of cyclin E2 in gastric cancer cells. Upregulation of cyclin E2 was observed in gastric cancer tissues compared with the normal controls. Highly expressed cyclin E2 was inversely correlated with the level of miR-383 in gastric cancer tissues. Consistent with the decreased expression of cyclin E2 with miR-383, transfection of miR-383 induced cell cycle arrest at G1 phase in gastric cancer cells. Restoration of cyclin E2 significantly reversed the inhibitory effect of miR-183 on gastric cancer cell proliferation. Collectively, our results characterized the suppressive role of miR-383 in gastric cancer partially through targeting cyclin E2.
Introduction
Gastric cancer is one of the most common malignancies around the world, which seriously threatens the health or life of the patients (Shi et al., 2017; Zheng et al., 2017; Coburn et al., 2018). Currently, surgery combined with radiotherapy and chemotherapy has been extensively applied in the clinical practice; however, the prognosis of patients with advanced gastric cancer remains very poor. Therefore, exploring the molecular mechanisms underlying the progression of gastric cancer is quite necessary to design novel strategies for the treatment of gastric cancer.
miRNAs are characterized as a class of small noncoding RNAs with the length of 20–25 nucleotides (Bartel, 2004; Mohr and Mott, 2015). miRNAs directly bind the 3′-untranslated region (UTR) of the target genes, leading to the degradation or translation inhibition of the mRNAs (Fabian et al., 2010). miRNAs play important roles in regulating cell proliferation, differentiation, and apoptosis through negatively modulating the expression of target genes (Ambros, 2004). Notably, the involvement of miRNAs in cancers has been demonstrated by increasing evidence (Kwak et al., 2010; Farazi et al., 2011; Qu et al., 2011; Gentilin et al., 2016; Momtazi et al., 2016). miRNAs decreased the expression of oncogenes or tumor suppressors and affected the tumorigenesis (Farazi et al., 2011). Mounting evidence has suggested the important roles of miRNAs in regulating the growth of gastric cancer cells. For example, miR-6852 inhibited the proliferation and invasion of gastric cancer cells through targeting the forkhead box J1 (Yu et al., 2018). Downregulation of miR-1294 in gastric cancer was correlated with the poor prognosis of the patients (Shi et al., 2018). The function of more miRNAs in gastric cancer remains to be explored.
Recent studies showed the downregulation of miR-383 in human cancers (Liu et al., 2016; Cui et al., 2018; Gu et al., 2018; Li et al., 2018; Teng et al., 2018). Overexpression of miR-383 suppressed the growth of colorectal cancer (Li et al., 2018). Upregulation of miR-383 inhibited the proliferation, invasion, and migration of colon cancer cells (Cui et al., 2018). The tumor suppressive role of miR-383 was also found in lung cancer (Gu et al., 2018). These studies suggested the critical function of miR-383 in regulating the malignant behaviors of cancer cells; however, the involvement of miR-383 in gastric cancer has not been fully understood.
In this study, the expression of miR-383 in gastric cancer tissues was detected and molecular mechanisms by which miR-383 regulated the progression of gastric cancer were investigated.
Materials and Methods
Tissue sample and cell culture
Sixty gastric cancer patients were enrolled in this study at the People's Hospital of Yichang Center from April 2012 to August 2014. Paired tumor tissues and adjacent normal tissues were obtained from these patients through surgical resection. None of these patients received chemotherapy or radiotherapy before being recruited into this study. The procedure of tissue collection was approved by the ethics review committee of the People's Hospital of Yichang Center according to the principles of the Declaration of Helsinki (The institutional approval number: 2016020366). Written informed consents were received from all the participants.
Human gastric cancer cell lines including MGC-803, MKN45, SGC7901, and U87 were purchased from the American Type Culture Center (ATCC). All the cells were cultured in DMEM supplemented with 10% FBS (Hyclone) and 1% penicillin–streptomycin solution (Invitrogen) in a 37°C incubator with 5% CO2.
Reverse transcription-quantitative PCR
Total RNA was extracted using the Trizol reagent (Invitrogen, ThermoFisher Scientific, Inc.) according to the manufacturer's instructions. The RNA concentration was determined using the Nano-Drop spectrophotometer (Thermo, Waltham, MA). Reverse transcription was performed with the High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA) according to the manufacturer's protocol. The expression of miR-383 was detected by real-time PCR with the SYBR Premix Ex TaqTM Kit (Takara, Shiga, Japan) on the 7500 Fast Real-Time PCR system (Applied Biosystem). The expression of U6 RNA was detected as the internal control. The PCR cycling conditions were as follows: 95°C for 5 min, 40 cycles at 95°C for 10 s, and 60°C for 1 min. The relative expression of miR-383 was calculated with the 2−ΔΔCq method.
Cell proliferation assay
The cell proliferation was evaluated with the Cell Counting Kit-8 (CCK-8; Dojindo, Japan) according to the manufacturer's instructions. Gastric cancer cells (∼2 × 103) transfected with miR-383 or negative control miRNA were seeded in a 96-well plate. After transfection for 24 h, 10 μL of CCK-8 reagent was added into the medium at the indicated time points and cultured for 2 h. The absorbance of each well at the wavelength of 450 nm was determined using the microplate reader (Bio-Rad).
Western blot
Gastric cancer cells transfected with miR-383 mimics or negative control miRNA were harvested and lysed with the radioimmunoprecipitation assay buffer lysis buffer (Beyotime Institute of Biotechnology, Shanghai, China). Equal amount of protein was separated by 15% SDS-PAGE and transferred onto the polyvinylidene difluoride membrane (Millipore, Billerica, MA). The membrane was blocked with 5% nonfat milk for 1 h at room temperature (RT) and then incubated with the primary antibody (anti-cyclin E2, cat#4132; Thermo Fisher, Waltham, MA; anti-GAPDH, cat#39-8600; Thermo Fisher) for 2 h at RT. After that, the membrane was washed twice with phosphate buffered saline and incubated with horseradish peroxidase-conjugated secondary antibody for 1 h at RT. The membrane was developed using the enhanced chemiluminescence kit (Pierce) according to the manufacturer's instructions.
Cell apoptosis
The percentage of cell apoptosis was detected with the Annexin V-APC/7-AAD Kit (Thermo Fisher Scientific, Inc.) according to the manufacturer's instructions. In brief, cells transfected with miR-383 mimics or control vector were harvested and resuspended to 1 × 106 cells/mL with the binding buffer. Cells were then stained with 5 μL of APC and 5 μL of 7-AAD for 15 min at RT in the darkness. After that, the cell suspension was diluted with 400 μL of binding buffer and the apoptosis was determined by flow cytometry (FACScan; BD Biosciences) according to the instructions.
Dual-luciferase reporter assay
The 3′-UTR sequence of cyclin E2 was amplified and inserted into the pGL3 plasmid (Promega Corporation, Madison, WI). Mutant 3′-UTR sequence of cyclin E2 was generated by introducing five mutations in seeding region of miR-383. Cells were seeded into a 24-well plate and transfected with the wild-type or mutant 3′-UTR of cyclin E2 in the presence of miR-383 mimics or negative control miRNA. After transfection for 36 h, cells were harvested and the luciferase activity was measured with the Dual Luciferase Reporter Assay (Promega Corporation) according to the manufacturer's instructions. The Renilla luciferase activity was determined as the internal control for normalization.
Statistical analysis
The data are presented as mean ± standard derivation. The statistical analysis was performed with SPSS software version 13.0 (SPSS, Inc., Chicago, IL). Student's t test was performed to analyze the difference between two groups. Comparison among three or more groups was determined with the one-way analysis of variance followed by least-significant difference post doc test. p < 0.05 was considered as statistical significance.
Results
miR-383 was significantly downregulated in gastric cancer tissues
To explore the potential involvement of miR-383 in gastric cancer, the expression of miR-383 in paired gastric cancer tissues and adjacent normal tissues was determined by reverse transcription-quantitative PCR (RT-qPCR). The level of miR-383 was significantly decreased in gastric cancer tissues when compared with that of the normal tissues (Fig. 1A). Consistent with this, the expression of miR-383 in gastric cancer cells was also downregulated when compared with the normal gastric epithelium GES-1 cells (Fig. 1B). To further investigate the role of miR-383 in the progression of gastric cancer, the correlation between the expression of miR-383 and the clinical features of gastric cancer patients was analyzed. The cutoff value to distinguish the low or high level of miR-383 expression was defined with the median value of miR-383 expression among all the 60 patients. As given in Table 1, compared with patients harboring relative high level of miR-383, reduced expression of miR-383 was significantly correlated with the tumor size, tumor node metastasis, stage and metastasis. However, no significant correlation was observed between the expression of miR-383 with the age and gender of gastric cancer patients (Table 1). Collectively, these results indicated the downregulation of miR-383 in gastric cancer.
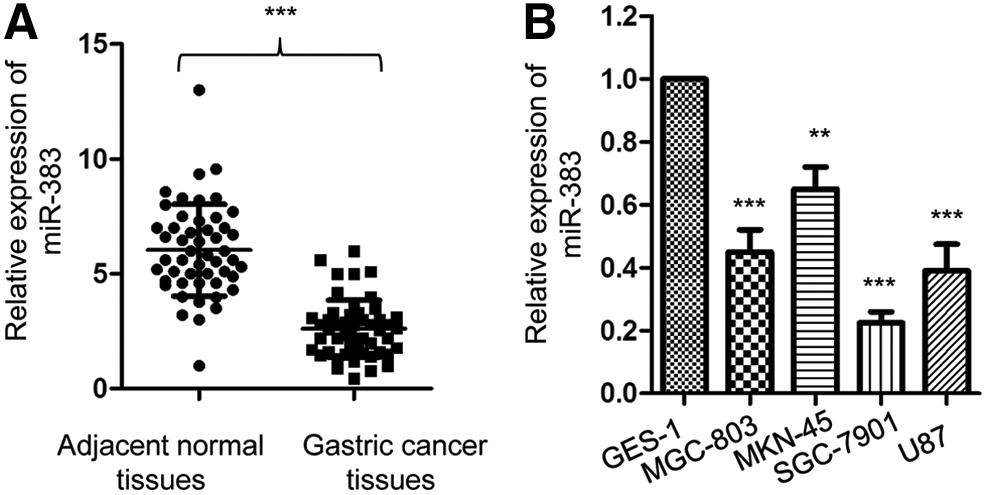
miR-383 was downregulated in gastric cancer.
Association Between the Expression of miR-383 and the Clinicopathological Characteristics of Gastric Cancer Patients
TNM, tumor node metastasis.
Overexpression of miR-383 inhibited the proliferation of gastric cancer cells
To evaluate the effect of miR-383 on the growth of gastric cancer cells, both SGC-7901 and MGC-803 cells were transfected with miR-383 mimics or control miRNA. The overexpression of miR-383 was confirmed by RT-qPCR (Fig. 2A, C). The proliferation of both SGC-7901 and MGC-803 with the transfection of miR-383 mimics or control miRNA was detected by CCK-8 assay. The results indicated that compared with the control group, overexpression of miR-383 significantly suppressed the proliferation of gastric cancer cells (Fig. 2B, D). The suppressive function of miR-383 in regulating the growth of gastric cancer cells was also validated by in vitro migration assay. As shown in Figure 2E, transfection of miR-383 significantly decreased the migration of both SGC-7901 and MGC-803 cells. Moreover, the cell apoptosis with the overexpression of miR-383 was also evaluated through FACS analysis. Compared with the control group, overexpression of miR-383 significantly promoted the apoptosis of both SGC-7901 and MGC-803 (Fig. 2F). These results demonstrated the inhibitory effect of overexpressed miR-383 in regulating the growth of gastric cancer cells.
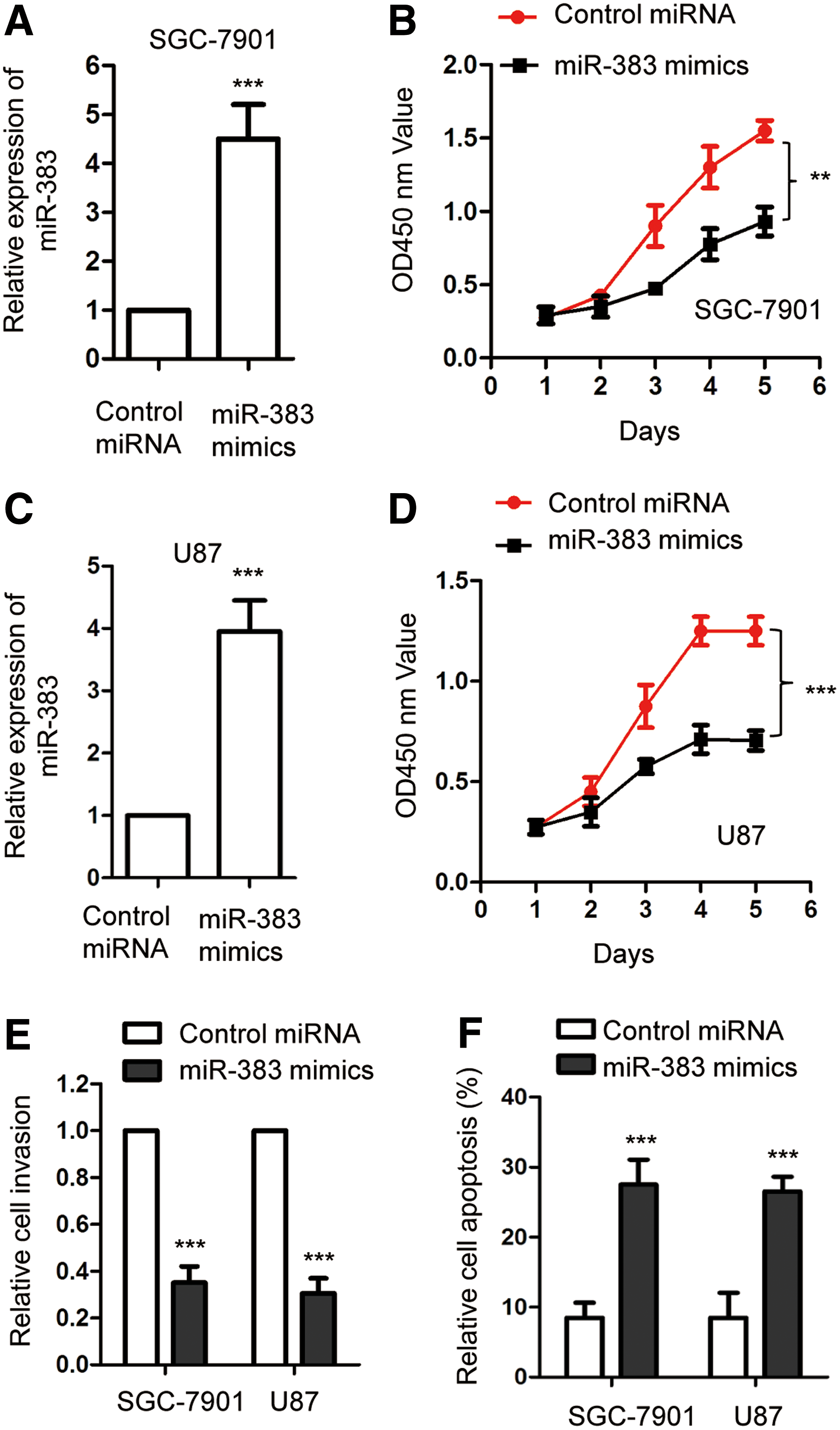
Overexpression of miR-383 inhibited the proliferation and induced the apoptosis of gastric cancer cells.
Cyclin E2 was a target of miR-383
To determine the underlying molecular mechanism by which miR-383 regulated the growth of gastric cancer cells, the potential targets of miR-383 were predicted with the miRDB database. The 3′-UTR of cyclin E2 was predicted containing the complementary seeding sequence of miR-383 (Fig. 3A). To further confirm this, luciferase reporter assay was performed by cotransfecting the reporter vector containing wild-type or mutant 3′-UTR of cyclin E2 and miR-383. As presented in Figure 3B and C, overexpression of miR-383 significantly inhibited the luciferase activity of vector harboring wild-type but not mutant 3′-UTR of cyclin E2. These data suggested the binding of miR-383 with the 3′-UTR of cyclin E2. To detect whether the binding affected the mRNA stability of cyclin E2, RT-qPCR assay was performed with SGC-7901 and MGC-803 cells expressing control or miR-383 mimics. The result showed that overexpression of miR-383 significantly decreased the mRNA level of cyclin E2 (Fig. 3D). Consistent with this result, transfection of miR-383 suppressed the protein expression of cyclin E2 in both SGC-7901 and MGC-803 cells (Fig. 3E). These results indicated cyclin E2 as a target of miR-383 in gastric cancer cells.
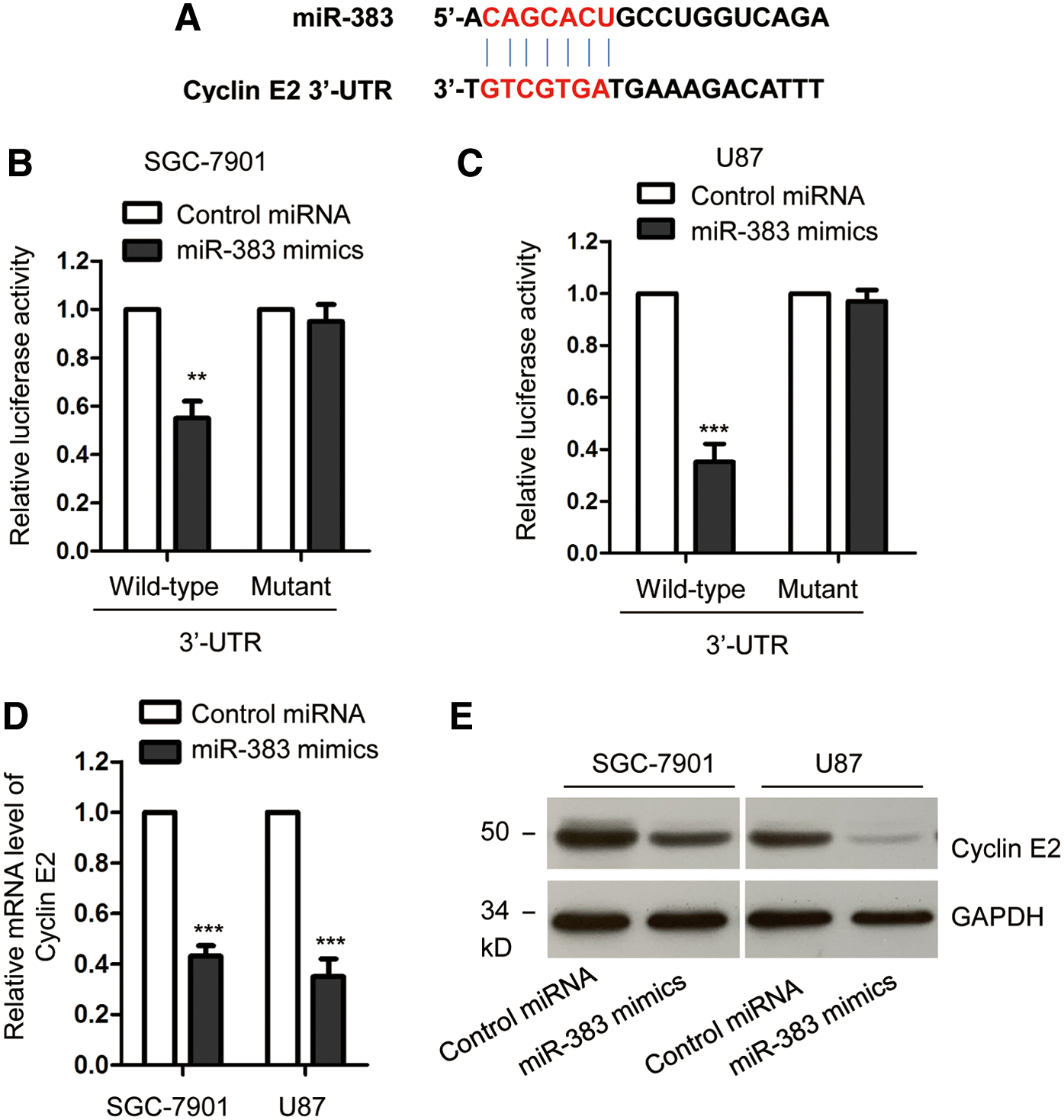
Cyclin E2 was a target of miR-383 in gastric cancer.
Cyclin E2 was upregulated in gastric cancer tissues and inversely correlated with the level of miR-383
As cyclin E2 was negatively regulated by miR-383 in gastric cancer, the expression of cyclin E2 in paired gastric cancer tissues and adjacent normal tissues was detected. As shown in Figure 4A, compared with the adjacent normal tissues, the expression of cyclin E2 was significantly increased in gastric cancer tissues (Fig. 4A). Consistent with this, the level of cyclin E2 was higher in gastric cancer cells than that in the normal GES-1 cells (Fig. 4B). The correlation between the expression of miR-383 and cyclin E2 was also analyzed with the Spearman test. As shown in Figure 4C, there was a significant negative correlation between the level of miR-383 and cyclin E2 in gastric cancer tissues. These results supported the conclusion that miR-383 targeted cyclin E2 and negatively modulated the expression of cyclin E2 in gastric cancer.

The expression of cyclin E2 was significantly inverse correlated with the level of miR-383 in gastric cancer.
Overexpression of miR-383 induced cell cycle arrest at G1 phase
Previous studies have demonstrated the essential role of cyclin E2 in regulating the cell cycle progression from G1 to S phase. As miR-383 decreased the expression of cyclin E2, to explore the effect of miR-383 on the cell cycle progression of gastric cancer cells, both SGC-7901 and MGC-803 cells were transfected with control vector or miR-383 mimics and the cell cycle was analyzed by FACS assay. The result showed that overexpression of miR-383 significantly inhibited the cell cycle progression from G1 to S phase (Fig. 5A, B).
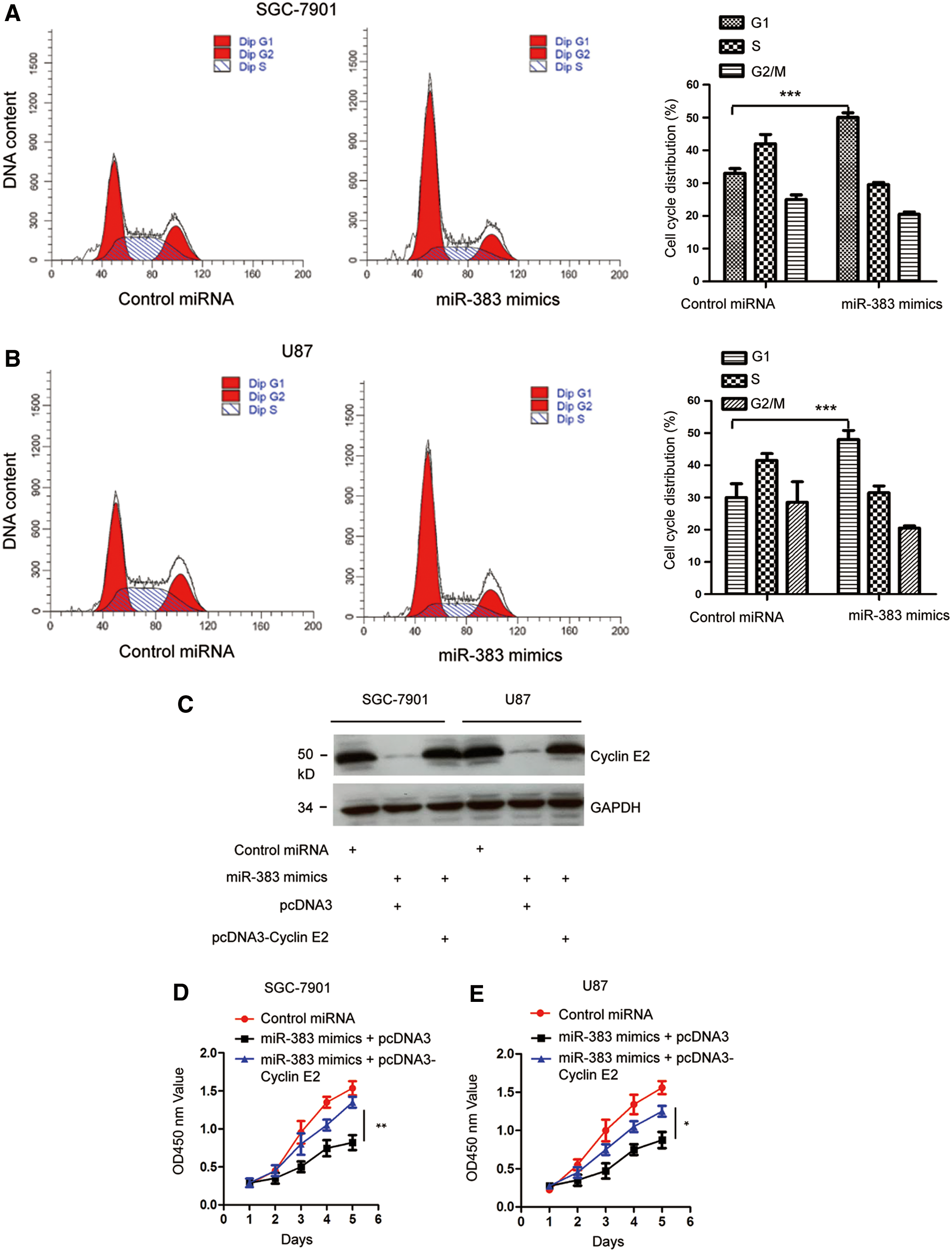
Overexpression of miR-383 induced cell cycle arrest.
To assess whether the suppressive effect of miR-383 on the growth of gastric cancer cells was through regulating cyclin E2, the expression of cyclin E2 was restored by transfecting Flag-cyclin E2 into both SGC-7901 and MGC-803 cells. The expression of cyclin E2 was validated by Western blot with anti-Flag antibody (Fig. 5C). CCK-8 assay showed that overexpression of miR-383 inhibited the proliferation of gastric cancer cells, whereas restoration of cyclin E2 significantly reversed the suppressive effect of miR-383 on the proliferation of both SGC-7901 and MGC-803 cells (Fig. 5D, E). These results demonstrated that miR-383 targeted cyclin E2 and suppressed the growth of gastric cancer cells.
Discussion
Increasing evidence has suggested the important roles of miRNAs in regulating the progression of cancers through acting as oncogenes or tumor suppressors (Iorio and Croce, 2017; Hosseinahli et al., 2018). In this study, miR-383 was significantly downregulated in gastric cancer tissues and cell lines. Reduced expression of miR-383 was associated with the poor prognosis of cancer patients, including lymph node metastasis and advanced clinical stage. These results suggested the potential involvement of miR-383 in the progression of gastric cancer.
The important regulatory function of miR-383 in cancers has been demonstrated by recent studies. For instance, miR-383 inhibited the proliferation and invasion of ovarian cancer cells (Han et al., 2017; Jiang et al., 2019). miR-383 was also identified as a promising biomarker for the prognosis of nonsmall cell lung cancer (Shang et al., 2016). In addition, the tumor suppressive function of miR-383 was also found in colon cancer and thyroid cancer (Li et al., 2018). In this study, overexpression of miR-383 significantly inhibited the proliferation, migration, and induced apoptosis of gastric cancer cells. These results indicated miR-383 as a negative regulator for the progression of gastric cancer cells. The potential tumor suppressive function of miR-383 in gastric cancer needs further confirmation through in vivo experiments.
It has been well established that miRNAs regulate cancer progression through modulating the expression of target genes. miR-383 was reported to directly bind the 3′-UTR of lactate dehydrogenase A in ovarian cancer cells and suppressed the glycolysis (Han et al., 2017). miR-383 targeted the insulin-like growth factor 1 receptor and modulated the invasion of glioma cells (He et al., 2013). In addition, the Per-Arnt-Sim domain-containing protein 1 was identified as a target of miR-383 in lung cancer (Ma et al., 2016). In this study, to further understand the role of miR-383 in gastric cancer, the binding targets of miR-383 were predicted with the bioinformatics analysis. The 3′-UTR of cyclin E2 was predicted containing the potential bindings sites of miR-383. Further experiments confirmed the binding between miR-383 and the 3′-UTR of cyclin E2. Consequently, overexpression of miR-383 significantly decreased both the mRNA and protein expression of cyclin E2 in gastric cancer cells. These results demonstrated that miR-383 targeted cyclin E2 and negatively regulated the expression of cyclin E2 in gastric cancer. Cyclin E2 activates CDK2 in G1 phase and triggers the transition from G1 to S phase in the cell cycle (Hwang and Clurman, 2005). Overexpression of cyclin E2 in cancer cells was associated with the progression of cancers (Payton et al., 2002). Interestingly, as a key regulator of the cell cycle progression, cyclin E2 has been identified as the target of a variety of miRNAs in cancers. For example, miR-30a suppressed the expression of cyclin E2 and inhibited the growth of castration-resistant prostate cancer (Zhang et al., 2016). Overexpression of miR-30d-5p inhibited the proliferation and motility of nonsmall cell lung cancer through targeting cyclin E2 (Chen et al., 2015). These studies suggested the critical function of cyclin E2 in miRNAs-mediated tumorigenesis. As miRNAs have many different targets, the involvement of other target genes deserves further investigation.
In conclusion, our results showed the downregulation of miR-383 in gastric cancer, which was significantly correlated with the progression of gastric cancer patients. Mechanism studies uncover that overexpression of miR-383 suppressed the proliferation of gastric cancer cells through targeting cyclin E2. These findings indicated miR-383 as a promising candidate to interrupt the progression of gastric cancer.
Conclusions
Our results uncovered the tumor suppressive role of miR-383 in gastric cancer cells through targeting cyclin E2. Decreased expression of miR-383 was significantly associated with the poor prognosis of gastric cancer patients. These findings suggested miR-383/cyclin E2 axis as a potential target for gastric cancer treatment.
Footnotes
Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.