
Abstract
Tea plant is an important economic crop on a global scale. Its yield and quality are affected by abiotic stress. The calcineurin B-like protein (CBL) and CBL-interacting protein kinase (CIPK) family genes play irreplaceable roles in plant development and stress resistance. More and more CBL-CIPK genes have been identified, but a few CBL-CIPK genes have been cloned and characterized in tea plants. In this study, 7 CsCBLs and 18 CsCIPKs were identified based on the tea plant genome. Physicochemical properties, phylogenetic, conserved motifs, gene structure, homologous gene network, and promoter upstream elements of these 25 genes were analyzed. Conserved motifs of these genes varied with phylogenetic tree node. From the genetic structure, members of the tea plant CIPK gene family can be divided into two types: intron rich and no intron. Many stress-related elements were found in the 2000 bp upstream of the promoter, and PlantCARE predicted that CsCBL4 contained 30 stress-related elements. PlantPAN2 shows that CsCIPK6 contains 48 ABRELATERD1; CsCIPK17 contains 37 GT1CONSENSUS; CsCIPK3 contains 64 MYBCOREATCYCB1; CsCBL3 contains 52 SORLIP1AT; CsCBL5 contains 65 SURECOREATSULTR11; and CsCIPK11 contains 83 WBOXATNPR1. In addition, eight genes were selected for quantitative real-time PCR (RT-qPCR) to detect their expression profiles under high-temperature, low-temperature, salt, and drought treatments. These genes were found to be responsive to one or more abiotic stress treatments. The expression levels of CsCBL4, CsCIPK2, and CsCIPK14 were similar, and they were homologous to AtSOS3 and AtSIP3 and AtSIP4 in Arabidopsis, which were involved in the SOS pathway. This study provides insight into the potential functions of the CsCBL and CsCIPK of tea plant.
Introduction
Calcium signaling is involved in multiple physiological and developmental processes (Harmon, 2003). Ca2+ is a common second messenger in plant cell signaling, which regulates various signal transduction pathways (Kolukisaoglu et al., 2004). The calcineurin B-like protein (CBL) belongs to a unique group of calcium sensors in plants (Cheong et al., 2010). CBLs were known to provide the specificity of signal by associating and regulating specific member of serine–threonine protein kinase family, CBL-interacting protein kinases (CIPKs). The complex formed by the combination of CBL and CIPK was mainly involved in many environmental signal responses and ion flux modulation (Nagae et al., 2003; Liu and Wu, 2013).
All of the CBL proteins contain four elongation factor (EF)-hand motifs that can bind at most four Ca2+ ions, and the number of amino acids spaced between the four EF-hand motifs is very conservative (Nagae et al., 2003; Sanchez-Barrena et al., 2005). Differences in the EF-hand motifs may result in different ways of binding to Ca2+ (Mahajan et al., 2006). CIPK possesses a conserved N-terminal kinase domain and a less conserved C-terminal regulatory domain, and is separated by a junction domain (Akaboshi et al., 2008). The regulatory domain of CIPK contains a conserved 24 amino acids NAF domain (also known as the FISL motif), which is a self-inhibiting domain and mediates interaction with CBL (Guo et al., 2001). Totally, the calcium signature can bind to the EF-hand domains of the CBL proteins. Then, the CBL proteins bind to the NAF domain of the C-terminal of the CIPKs (Hashimoto et al., 2012). Binding led to the activation of the kinase in a calcium-dependent manner.
The network of CBLs and CIPKs is widely involved in plant stress responses (Batistič and Kudla, 2009). To our knowledge, the first identified CBL-CIPK pathway was the salt overly sensitive (SOS) pathway (Mahajan and Tuteja, 2005). The SOS pathway contains three key points: SOS1 (Na+/H+ antiporter), SOS2 (CIPK24), and SOS3 (CBL4), which has been well characterized in mediating cellular signaling under salt stress, to maintain ion homeostasis (Ji et al., 2013). SOS1 pumps Na+ from cells, and the ion concentration increases to form salt stress. SOS2-SOS3 forms a complex at the root that phosphorylates SOS2 and activates the transport properties of SOS1 (Qiu et al., 2002). Overexpression of a single or several SOS pathway genes can enhance the salt tolerance of transgenic plants (Ma et al., 2014). On the contrary, plants lacking the ability to activate SOS1 have a reduced ability to discharge ions and accumulate more Na+. (Ariga et al., 2013). In addition, AtCBL2 mediates the activation of AKT1 by CIPK proteins (AtCIPK6, AtCIPK16, and AtCIPK23) in response to low potassium conditions and in the context of stomatal movement (Xu et al., 2010). It probably involved in regulating signaling responses to abscisic acid (Thoday-Kennedy et al., 2015).
Tea is a natural nonalcoholic beverage, rich in various secondary metabolites, such as flavonoids, theanine, and caffeine (Li et al., 2018). The flavor of tea is mellow, irritating, and healthy, making it one of the three most popular nonalcoholic beverages in the world (Wang et al., 2016a, 2016b; Cui et al., 2018). Tea plant [Camellia sinensis (L.) O. Kuntze] is mainly distributed in the Southeast Asian monsoon region and has a commercial value with ∼2000 years of history in China (Chen et al., 2010). Common environmental stresses, such as drought, cold, heat, and salt, can affect the metabolism of unique compounds in tea (Li et al., 2014). These stresses pose serious threat to the yield and quality of tea (Zhou et al., 2014). “Longjing 43” is bred from the Longjing group through a clonal selection by the Tea Research Institute of the Chinese Academy of Agricultural Sciences (Lou et al., 2015). It is widely cultivated for its high yield and stability, and is also used as a model for tea plant genetic breeding research.
In addition to the model crops Arabidopsis and rice, other species have CBL-CIPK interaction networks, such as cassava and tomato (Kolukisaoglu et al., 2004; Kabir and Wang, 2010; Mo et al., 2018), which can respond to abiotic stress, hormones, and heavy metals through an interactive network. However, there are similarities and differences in the number, structure, location, and function of genes family among different species. Although the tea genome sequence has been published (Wei et al., 2018), up to now, the CBL-CIPK family has not been fully characterized in tea plant. How CBL-CIPK genes respond to abiotic stress in tea plant is still unclear.
In this study, we performed a genome-wide identification and analysis of CBL and CIPK family members in tea plants. The phylogeny, conserved motifs, gene structure, Go enrichment analysis, and interaction networks were analyzed. Previous studies have reported the function of the CBL and CIPK genes in abiotic stress resistance. Based on Arabidopsis thaliana expression data under different stresses, and homologous genes between A. thaliana and tea plant, eight genes were selected to analyze the expression levels of heat, cold, salt, and drought in tea plant (Table 1) (Huang et al., 2011; Pandey et al., 2015). This analysis provides a framework for exploring the CBL-CIPK family in tea plant and serves as the basis for future research.
The Characteristics of CBL and CIPK Family Members of Tea Plant
CBL, calcineurin B-like protein; CDS, coding sequence; CIPK, CBL-interacting protein kinase; EF, elongation factor; MW, molecular weight; pI, isoelectric point.
Materials and Methods
Identification of CsCBL and CsCIPK genes
All candidate CBL and CIPK sequences of tea plant (Supplementary Table S1) were filtered from the tea plant genome. The candidate sequences were obtained by retrieving the names of CBL and CIPK for the genome annotation information (Wei et al., 2018). Then, the online programs of SMART and National Center for Biotechnology Information's (NCBI's) Conserved Domain Database were used to survey the conserved domains of the candidate proteins. The CBL protein sequences were only accepted if they simultaneously contained the EF-hand calcium binding domains (PSSM-ID: 330173) (Supplementary Table S2), as well as CIPK sequences contained the conserved Pkinase domain (PSSM-ID: 328722) and the NAF domain (PSSM-ID: 213380) (Supplementary Table S3).
Phylogenetic and amino acid sequences analysis
Protein properties, including molecular weight (MW) and isoelectric point (pI) of CsCBLs and CsCIPKs, were predicted using the online tool Compute pI/MW. Closely related protein sequences of the Arabidopsis and rice were downloaded from the NCBI. Multiple sequence alignments of CBL and CIPK proteins were performed using ClustalX (Thompson et al., 2002). Maximum-likelihood (ML) trees were constructed using the aligned sequences by Molecular Evolutionary Genetics Analysis (MEGA) 7.0 software (Stecher et al., 2016). The genes belonging to tea plant were marked with red color. Motif analyses of CsCBLs and CsCIPKs were performed by Multiple Em for Motif Elicitation (MEME) and TBtools software with default parameters. The sequence files, gene structure files, and upstream sequence of the genes promoter were downloaded from the Tea Plant Information Archive (TPIA) (Wei et al., 2018). Gene structures were drawn using the gene structure display server (GSDS) program (Hu et al., 2015). The PlantCARE and PlantPAN2 online sites were used to predict the cis-acting elements and transcription factor binding sites (TFBSs) contained in the 2 kb sequence upstream of the genes (Chow et al., 2016). The cis-acting elements and TFBS associated with stress were screened using Excel. TBtools software was used for data visualization analysis.
Gene Ontology annotation
Functional annotation of the identified CsCBL and CsCIPK proteins was analyzed and visualized using Blast2GO v5.2 (Gotz et al., 2008). Proteins were described in terms of three categories under Gene Ontology (GO) classification, namely biological processes, molecular functions (MFs), and cellular components.
Expression levels under different organizations and treatments
Expression data for eight different tea plant tissues, as well as drought, salt, and Methyl jasmonate (MeJa) treatments, were downloaded from TPIA. In these databases, the tea plant cultivar “Shuchazao” was used to research. To better understand the expression mechanism of CsCBLs and CsCIPKs in tea plants, we obtained expression data for tea plants under drought and salt treatment at 24, 48, and 72 h; MeJA treatment at 12, 24, and 48 h; and expression data of different tissues of tea plant buds, flowers, fruits, first leaves, second leaves, third leaves, roots, and stems. The heat map was prepared using HemI software (Deng et al., 2014).
Preparation of plant materials
Tea plant cultivar “Longjing43” was grown in the greenhouse at the Tea Science Research Institute, College of Horticulture, Nanjing Agricultural University (Nanjing, China). The environmental parameters of the plant culture room are as follows: the photoperiod was 12 h, and the light intensity was 220 μmol·m−2·s−1. The relative humidity was 75%. The 2-year-old tea seedlings were placed in a container of 840 × 280 mm 98-hole black plug. The matrix consists of peat, vermiculite, and perlite (volume ratio D 3:2:1). In April 2018, tea plants were selected and treated in four different ways: cold (4°C), high temperature (38°C), and drought (irrigated with 20% PEG 6000) and salt stress (watered with 200 mM NaCl solution). Young leaves of the test tea plants were harvested at 0, 2, 4, 8, and 24 h after treatments. Five tea plants were randomly selected for each treatment, and the sample was a mixture of the first leaves, the second leaves, and the third leaves. The collected leaves were rapidly frozen in liquid nitrogen and then stored at −80°C for further analysis.
RNA isolation and quantitative real-time PCR analysis
Total RNA was isolated from young leaves of tea plants by an RNA quick isolation kit (Aidlab, Beijing, China). The extracted RNA concentration and purity were measured on a NanoDrop ND-2000 Spectrophotometer (Thermo Scientific). RNA integrity was determined by 1.5% denaturing agarose gel. Then, cDNA was synthesized using the PrimeScript RT reagent Kit with gDNA Eraser (TaKaRa, Dalian, China) according to the manufacturer's instructions. Genomic DNA was removed using gDNA Eraser before reverse transcription. The quantitative real-time PCR (RT-qPCR) assay was performed with three independent biological replicates and three technical replicates.
The specific primers used in this experiment were designed by Primer Premier 6.0 software, and the specificity of these primers was examined by Basic Local Alignment Search Tool (BLAST). CsGAPDH was used as the reference gene (Wu et al., 2016). The primer sequences used in this study are listed in Supplementary Table S4. The RT-qPCRs were performed on Bio-Rad CFX96 fluorescence RT-qPCR platform. The program of the RT-qPCR reaction was set as follows: denaturation at 95°C for 300s, 39 cycles at 95°C for 10s, 60°C for 10s and 72°C for 30s. Finally, the 2−ΔΔCT method was used to calculate the relative expression level (Livak and Schmittgen, 2001).
Statistical analysis
Data were analyzed using Statistical Package for Social Science (SPSS) statistical version 17.0 (SPSS, Inc., Chicago, IL). Differences in expression levels of different treatments were tested at a p < 0.05 level using Duncan's multirange test. Data are expressed as mean ± standard deviation.
Results
Identification of CsCBL and CsCIPK genes from tea plant
Candidate CBL and CIPK sequences were obtained from the annotation information of the tea plant genome. All of these CsCBL proteins contained EF-hand calcium binding domains. The CIPK sequences contained the conserved Pkinase domain and the NAF domain. As a result, 7 CsCBLs and 18 CsCIPKs were identified. The coding sequence length of CBL ranges from 501 to 1227 bp, and the encoded protein is 166–408 amino acids in length. The corresponding MW ranges from 19.05 to 46.64 kDa, and isoelectric point (pI) values range from 4.55 to 5.67.
The coding sequence length of CIPKs ranges from 1020 to 2040 bp, and the encoded protein is 339–679 amino acids in length. The corresponding MW ranges from 38.43 to 62.78 kDa, and the isoelectric point (pI) values range from 4.87 to 9.59. Furthermore, an orthologous analysis between C. sinensis and Arabidopsis showed that 25 sequences of tea plant were linked to their corresponding orthologs in Arabidopsis (Table 1 and Supplementary Table S5).
Phylogenetic and evolutionary analysis
To understand further the structural features of CBL and CIPK proteins in tea plant, multiple sequence alignment of full-length CsCBL and CsCIPK proteins was performed (Fig. 1). The evolution relationships between C. sinensis, A. thaliana and Oryza sativa were investigated by constructing ML tree. Most of the CBL and CIPK genes from tea plants can find homologous genes in Arabidopsis and rice. From the ML tree, CsCBL6 is close to AtCBL8, OsCBL5, and OsCBL4. CsCBL4 and AtCBL4, 5 have a close genetic distance. CsCBL2, 5 is in the same branch as OsCBL1 and AtCBL1, 9. The CIPK phylogenetic tree was divided into two subbranches. CsCIPK7 is closer to AtCIPK8 and OsCIPK8. CsCIPK9, 11, 12 are close to AtCIPK23 and OsCIPK23. CsCIPK14 and AtCIPK6 are close to each other. CsCIPK2, 3, 6 and AtCIPK11 are in the same branch.

Phylogenetic tree of CBL
Conserved motif analysis of CsCBL and CsCIPK
To explore the difference of the CBL and CIPK proteins among tea plants, Arabidopsis and rice, conserved motifs were predicted. The conserved motifs were predicted using the MEME program. Five conserved motifs were defined in CBLs, and 17 motifs in CIPKs (Figs. 2 –5). Members of a close evolutionary relationship displayed uniform or similar motif compositions. In the majority of the CBL proteins, motifs 1–4 corresponded to the conserved domain EF hand. CIPK possesses a conserved N-terminal kinase domain and a less conserved C-terminal regulatory domain. The NAF motif (motif 9) is included in all CIPK sequences. Most of the conserved motifs are same at the N-terminal conserved domain region. The motifs have differences in the diversified C-terminal region of the CIPK proteins. As a whole, the distribution of the main conserved motifs in tea plants was similar to other species.
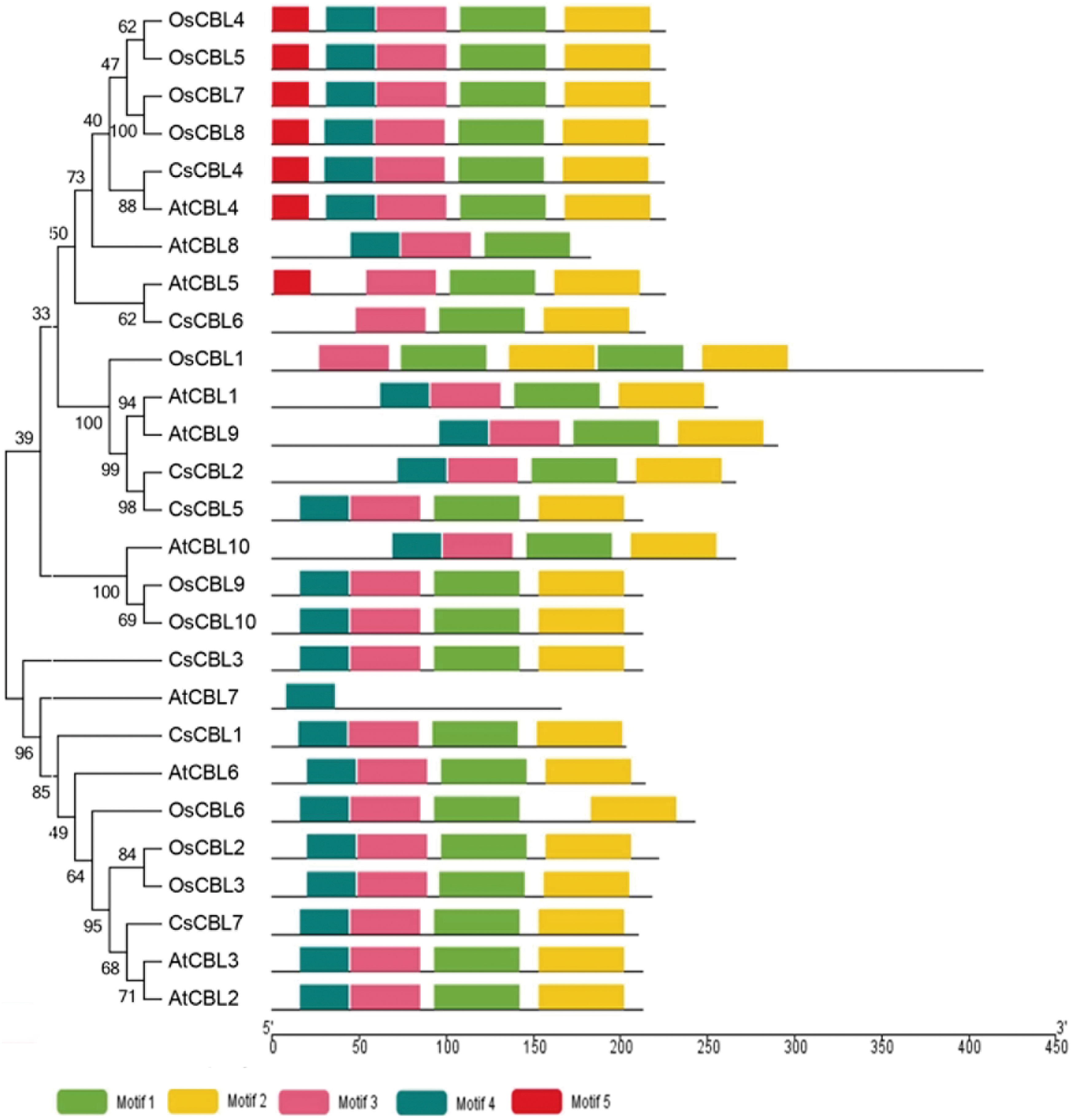
Motif analysis of CBL proteins from C. sinensis, A. thaliana and O. sativa. A phylogenetic tree was constructed by MEGA7.0. Motifs in the CBL proteins were elucidated by MEME. Different motifs are represented by different color boxes. MEME, Multiple Em for Motif Elicitation. Color images are available online.
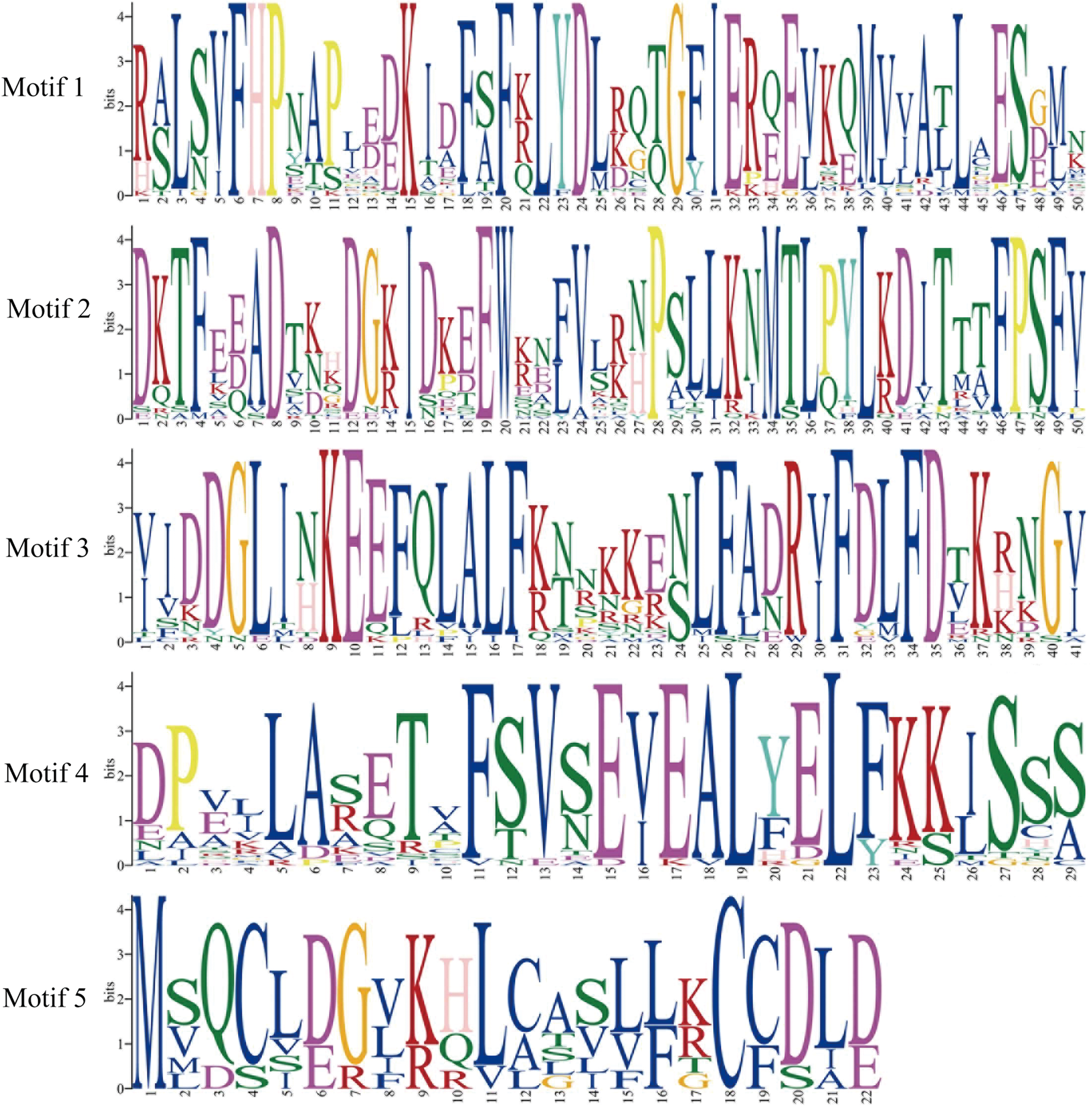
The regular expressions of conserved motifs from CBL proteins. Color images are available online.
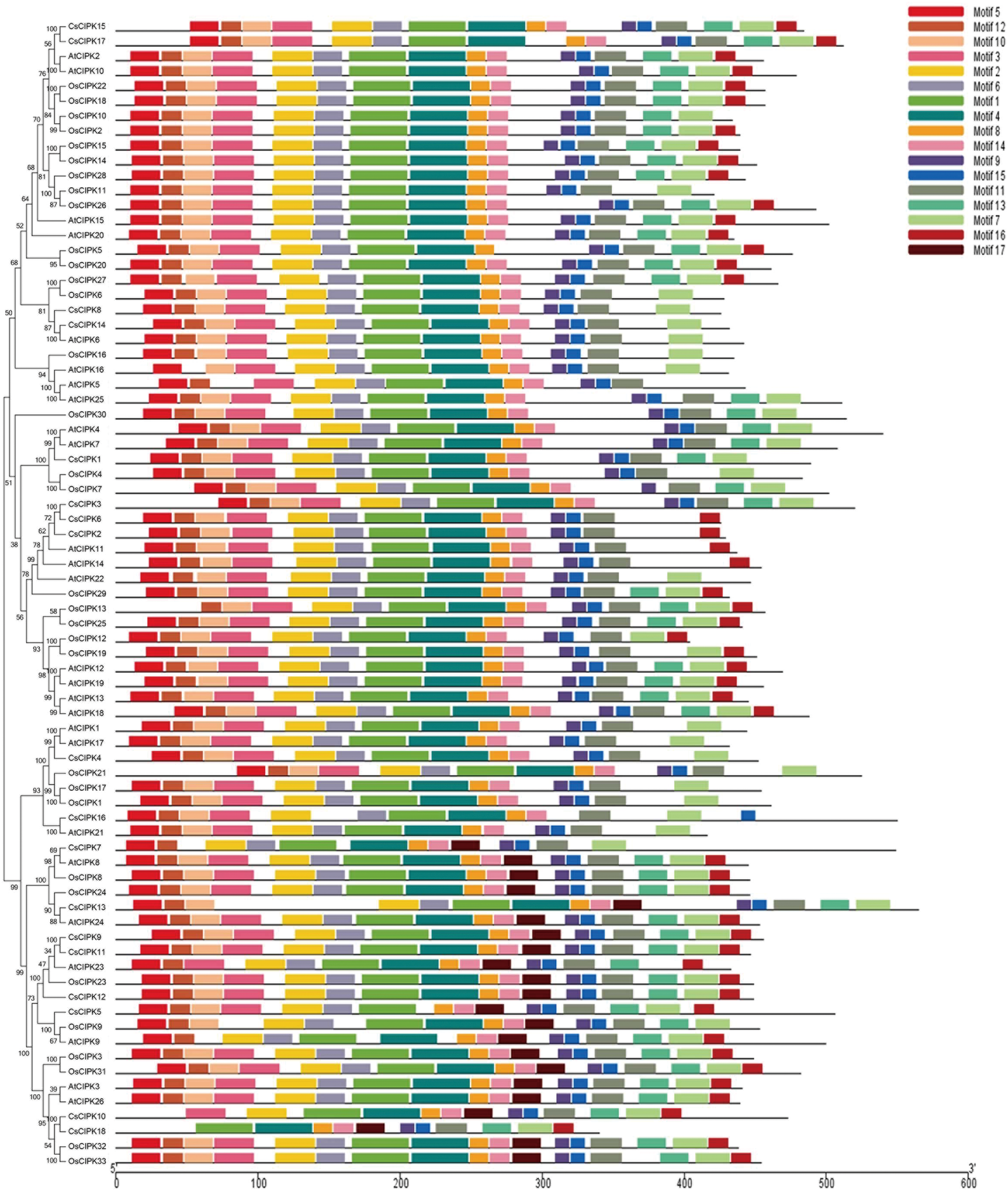
Motif analysis of CIPK proteins from C. sinensis, A. thaliana and O. sativa. A phylogenetic tree was constructed by MEGA7.0. Motifs in the CBL proteins were elucidated by MEME. Different motifs are represented by different color boxes. Color images are available online.
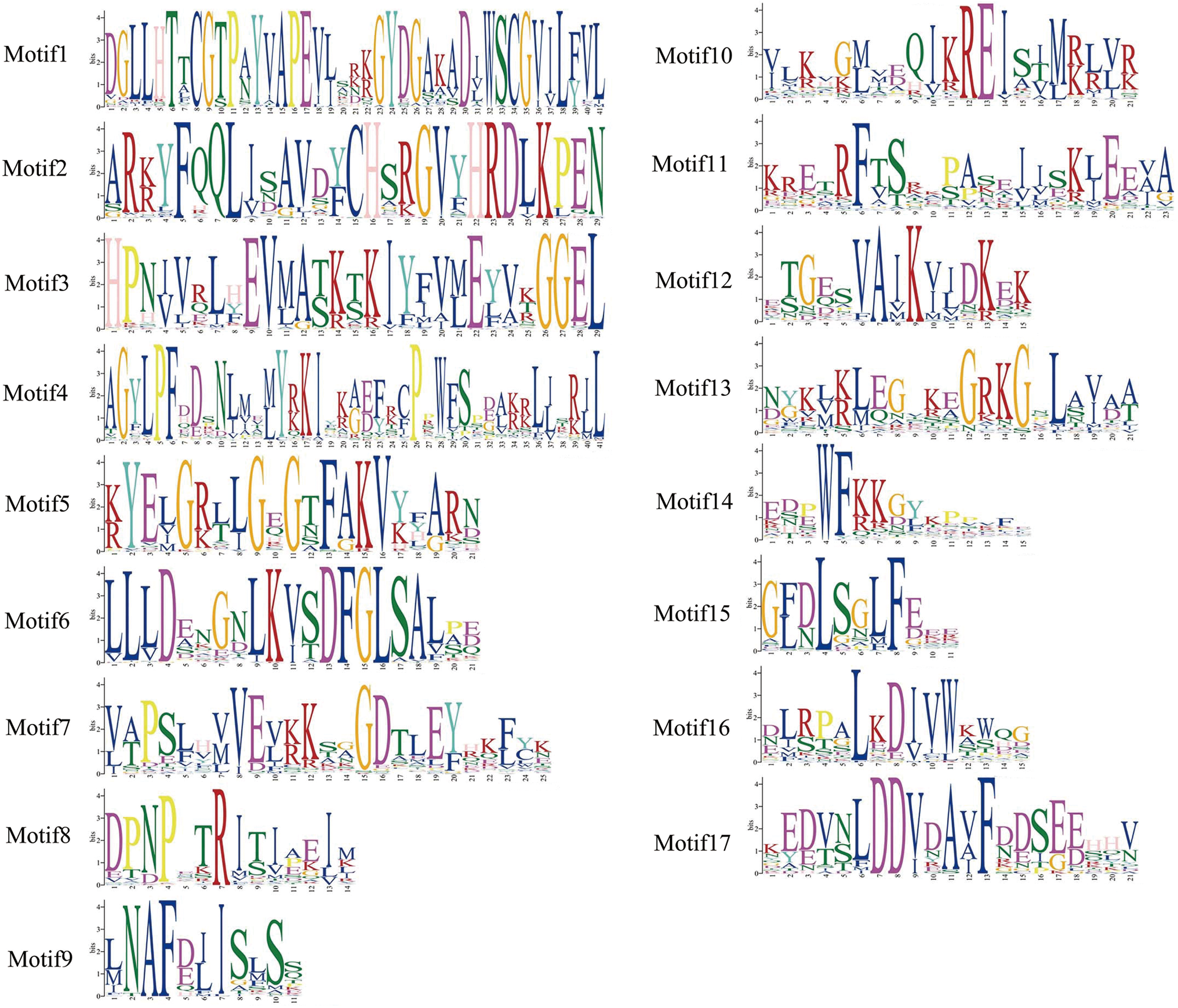
The regular expressions of conserved motifs from CIPK proteins. Color images are available online.
Gene structure analysis of CsCBLs and CsCIPKs
The genetic structure of CsCBLs and CsCIPKs was analyzed. Each CsCBL has multiple introns in its gene structure (Fig. 6a). CsCBL6 has three introns. CsCBL3 has 10 introns. CsCBL1, 2, 4, 5, 7 have untranslated region. The intron and exon positions of the CsCIPKs show two distinct types (Fig. 6b). One group including CsCIPK1, 2, 3, 6, 8, 14, 15, and 17 show no introns for most of them; while the rich exome includes CsCIPK4, 5, 7, 9, 10, 11, 12, 13, 16, and 18. Not only are there a large number of introns, but most of them contain untranslated region.
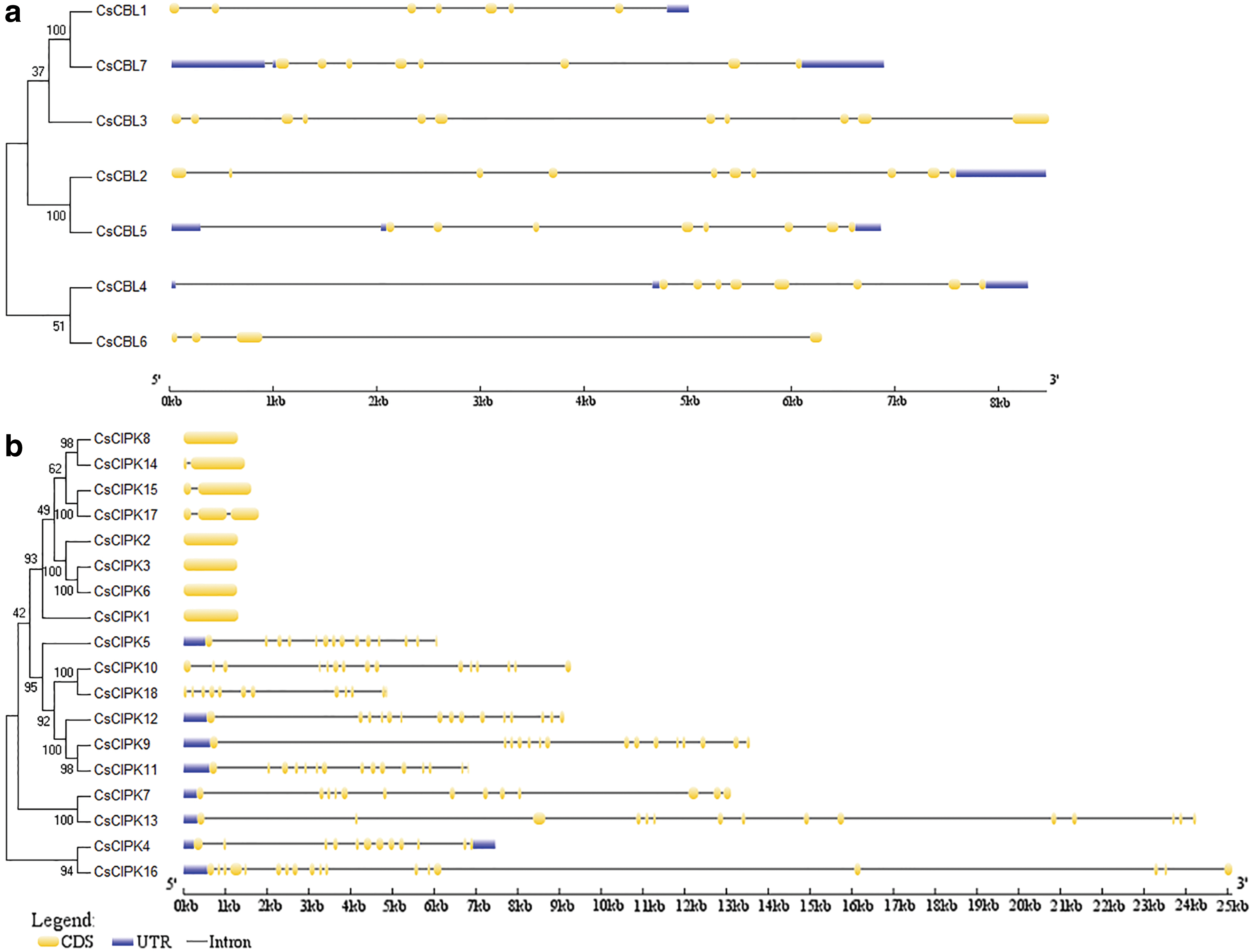
Gene structures of CsCBLs
GO annotation
Blast2GO v5.2 was applied to assign information on the biological function. A total of 7 CsCBLs and 18 CsCIPKs annotations were assigned to GO terms of biological processes, MF, and cellular components. Most of the CsCBLs' annotations were assigned to MF of ion binding. The distribution of GO levels, which is the closest level with regard to the number of terms, is shown in Figure 7.
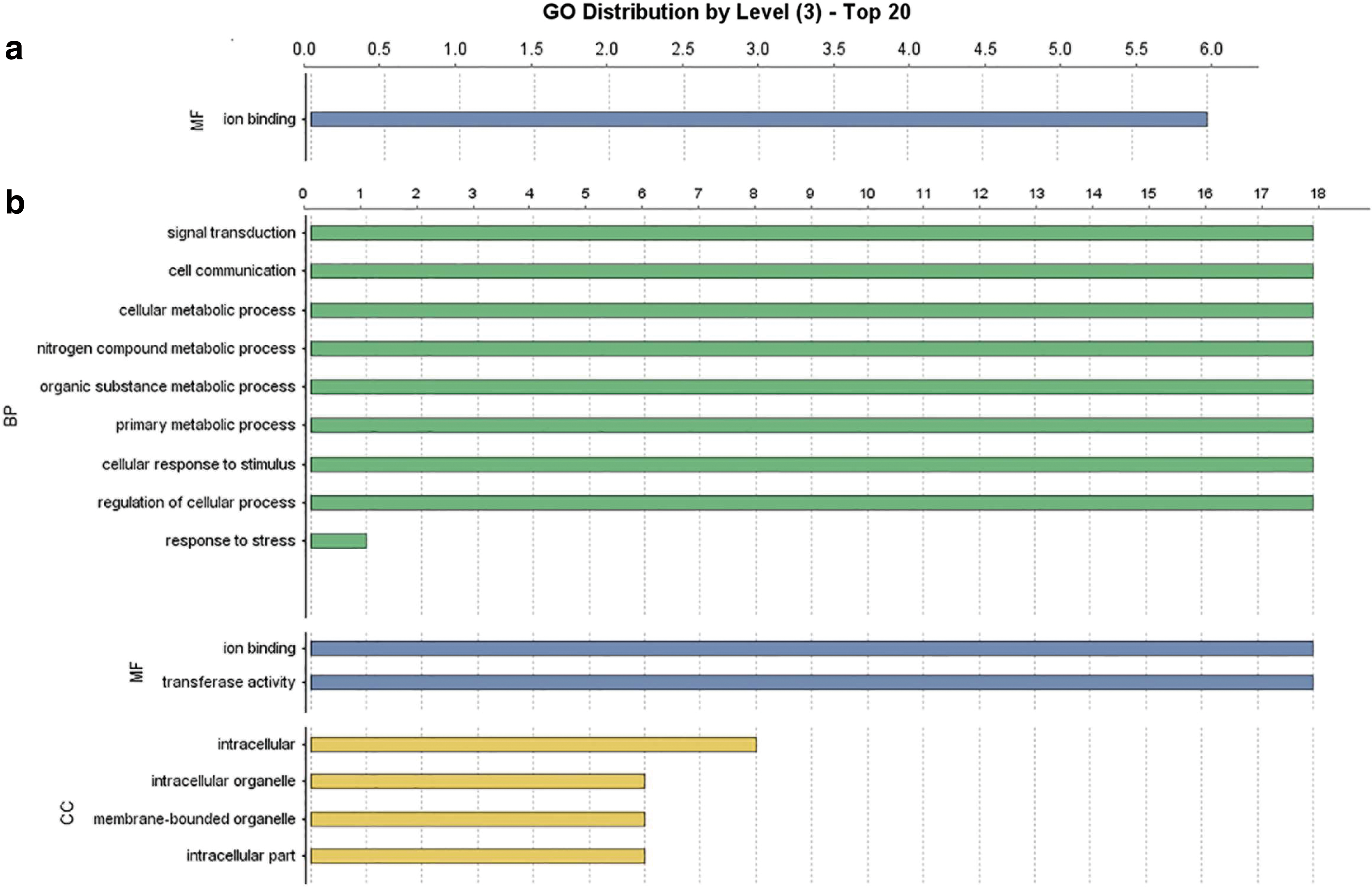
GO distribution of CsCBL
Interaction network of CBL-CIPK between C. sinensis and Arabidopsis
The protein–protein interactions of CBL-CIPK between C. sinensis and Arabidopsis were constructed. The function of proteins and physical interactions in C. sinensis were predicted (Fig. 8). Protein–protein interactions are shown in gray, and chemical–protein interactions are indicated in green. Most proteins are involved in calcium regulation. These CBL-CIPKs are involved in the interaction network, indicating that they might have complicated connections. CsCIPK2, CsCIPK13, CsCIPK14, CsCBL4, and CsCBL6 are highly similar to CIPK6 (SIP3 [SOS3-interacting protein]), CIPK11 (SIP4), CIPK24 (SOS2), and CBL4 (SOS3), which are involved in the Arabidopsis SOS pathway, suggesting that these three genes may involve in the SOS pathway of tea plant. Previous studies showed that AtSOS3 protein together with the interacting protein kinase AtSIP3 modulates the activity and plasma membrane (PM) targeting of the K+ channel AKT2 (Held et al., 2011).

Functional interaction networks of CsCBL and CsCIPK proteins in tea plant according to orthologs in Arabidopsis. Protein–protein interactions are shown in gray; chemical–protein interactions indicated in green. Color images are available online.
cis-Regulatory elements and TFBS analysis of CsCBL and CsCIPK
Stress-responsive elements were investigated in the promoter regions of CsCBLs and CsCIPKs. To explore the possible responsive mechanisms to abiotic stress, we characterized promoter regions in these 25 genes (2000 bp upstream of the translation start site). In the promoter regions, seven kinds of cis-elements were involved in responding to abiotic stresses in the promoter regions of CsCBLs and CsCIPKs (Fig. 9a). The number of stress-responsive elements ranged from 2 (CsCIPK9) to 30 (CsCBL4). The promoter region of the 14 genes contains W box element, where CsCIPK17 contains four W box elements. The promoter regions of the 10 genes have WUN-motif elements, and some genes contain multiple WUN-motif elements. The LTR element was found in 15 genes. Almost all promoter regions of genes have MYB, MYC, and MBS components.
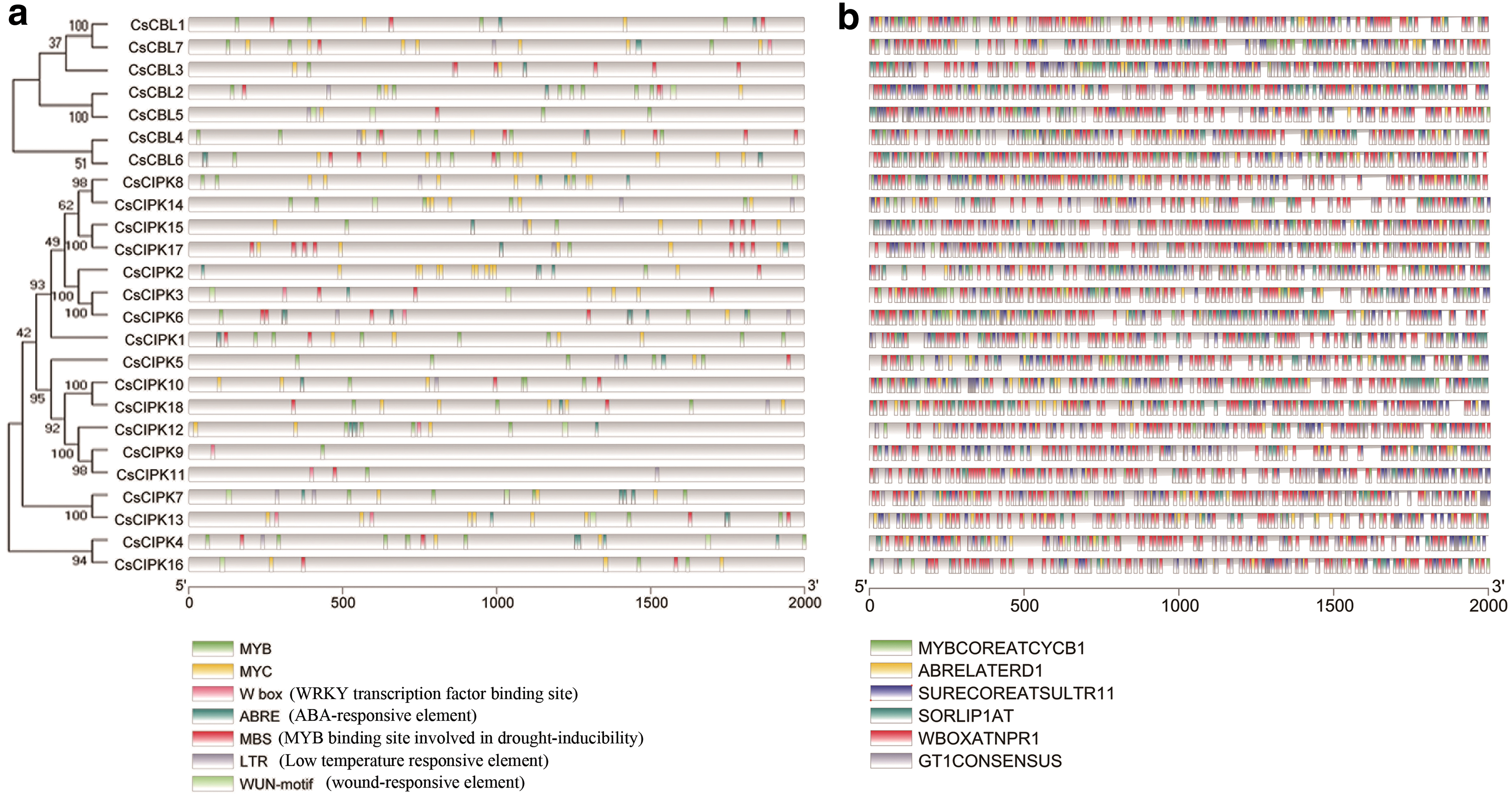
Putative cis-elements and transcription factor binding sites in the promoter regions of CsCBL and CsCIPK genes. PlantCARE online sites were used to predict the cis-acting elements
The PlantPAN2 online site predicts TFBSs for the 2 kb sequence upstream of the gene. The six largest TFBSs were screened out (Fig. 9b). The upstream region of each gene contains a large number of abiotic stress and stress-resistant regulatory elements MYBCOREATCYCB1, ABRELATERD1, SURECOREATSULTR11, SORLIP1AT, WBOXATNPR1, and GT1CONSENSUS. Among them, the upstream 2 kb sequence of the CsCIPK6 promoter has 48 ABRELATERD1; upstream of CsCIPK17 contains 37 GT1CONSENSUS; upstream of CsCIPK3 contains 64 MYBCOREATCYCB1; upstream of CsCBL3 contains 52 SORLIP1AT; upstream of CsCBL5 contains 65 SURECOREATSULTR11; and upstream of CsCIPK11 contains 83 WBOXATNPR1.
Expression-level profiles under different tea tissues and treatments
The differential expression of 25 genes was shown in heat map (Fig. 10) (Supplementary Tables S6, S7, S8). The expression levels of CsCBL6 and CsCIPK8 were not detected under drought and salt treatment. The expression levels of CsCBL1 and CsCIPK18 were very low. Under drought treatment, the expression levels of CsCBL7, CsCIPK3, CsCIPK7, CsCIPK11, CsCIPK15, and CsCIPK17 continued to increase. The expression levels of CsCIPK1 and CsCIPK6 continued to decline. Under salt treatment, CsCBL7, CsCIPK3, CsCIPK7 expression continued to rise. The expression levels of CsCIPK5, CsCIPK9, CsCIPK12, and CsCIPK14 continued to decrease.
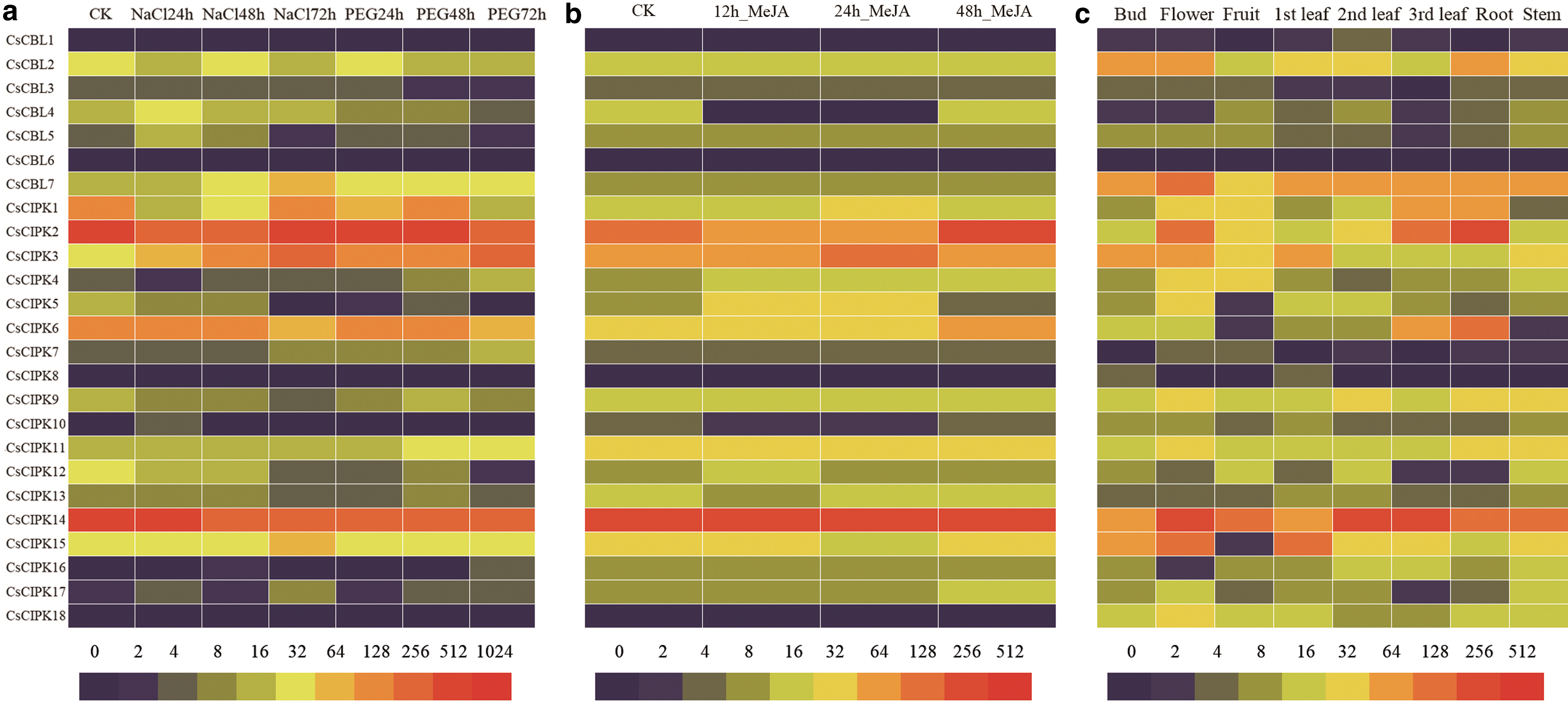
Heat map for expression of CsCBLs and CsCIPKs under different tissues and treatments. In the heat map, expression intensity is shown by different colors (red, high expression; blue, no expression):
The expression level of CsCBL6 was not detected under MeJA treatment. The expression levels of CsCBL1, CsCIPK8, and CsCIPK18 were low. The expression level of CsCIPK6 was elevated. The expression levels of CsCBL6 were not detected in eight different tea plant tissues. In the eight tea tree tissues, most of the genes showed significantly different expression patterns. However, CsCBL7 and CsCIPK14 are highly expressed in various tissues. In the same tissue, the expression levels of some genes are similar. For example, CsCIPK9 and CsCIPK11 are closer in phylogenetic relationships and have similar levels of expression in various tissues. The expression levels of CsCBL2, CsCBL3, CsCBL5, CsCBL7, CsCIPK15, and CsCIPK18 were highest in flowers. The expression levels of CsCIPK1, CsCIPK2, and CsCIPK6 were highest in the roots. The expression levels of CsCIPK9, CsCIPK11, CsCIPK12, and CsCIPK13 were highest in the stem.
Expression profiles of the CBL-CIPKs genes under abiotic stress treatments
Environmental stress can adversely affect plant growth and productivity, and trigger a range of morphological, physiological, biochemical, and molecular changes. Considerable evidence has shown that CBL-CIPK networks play vital roles in response to abiotic stresses (Li et al., 2009).
To explore the roles of CBL-CIPK gene in tea plant under diverse environmental conditions, eight genes, including two CsCBLs and six CsCIPKs, were selected, and the expression levels were detected in response to multiple stress treatments, including drought (20% PEG 6000), salt (200 mM NaCl), heat (38°C), and cold (4°C).
High-temperature treatment
Heat stress led to relatively low expression levels of CsCIPK5 and CsCIPK12 than the control (Fig. 11a). The levels of CsCIPK7 were higher than that of the control at 4, 8, and 24 h. Furthermore, five genes were highly induced under high-temperature treatment. CsCIPK2, CsCIPK14, CsCIPK18, CBL4, and CsCBL7 were upregulated compared with the control. During 24 h, the change tendency of the relative expression level was similar. The expression levels of CsCIPK2 and CsCIPK14 were 8.5 times and 12.5 times those in the control at 24 h.

Expression levels of CsCBLs and CsCIPKs under four abiotic stress treatments:
Low-temperature treatment
Under low-temperature treatment, CsCIPK5 and CsCIPK12 were downregulated (Fig. 11b). In contrast, CsCIPK7 was gradually upregulated and it reached its highest level at 24 h. CsCIPK2 showed a similar expression pattern to CsCIPK14, both of which decreased slightly at 2 h. Then, their expression levels began to rise and peaked at 8 h. Although CsCIPK18 was upregulated at 4 and 24 h, its relative expression levels were lower than those observed in controls at 2 and 8 h. The expression level of CsCBL4 was slightly higher than that in the control at 4 h. The expression level of CsCBL7 was higher than that in the control under whole treatment.
Salt treatment
Under salt stress treatment, the relative expression levels of CsCIPK2, CsCIPK5, and CsCIPK14 decreased first and then increased, and reached the maximum at 8 h (Fig. 11c). The expression trends of CsCIPK7 and CsCIPK18 were similar. The expression level of CsCIPK12 was relatively stable, and the change range was not large. The expression level of CsCBL4 at 2, 4, and 24 h was lower than that in the control. The expression level of CsCBL7 was the highest at 2 h, which was 17.4 times that in the control. The relative expression levels of CsCBL4, CsCIPK2, and CsCIPK14 showed similar trends.
Drought treatment
Under drought treatment, the relative expression levels of CsCIPK2, CsCIPK12, and CsCBL4 were similar. However, the expression level of CsCIPK12 was low at 4 h. The expression level of CsCIPK5 at 8 h was 3.5 times that of the control. CsCIPK7 and CsCIPK18 had similar trends, which first increased, then decreased, and later gradually increased. The highest expression level of CsCIPK14 was 7.2 times more than the control at 2 h. The expression trend of CsCBL7 involved a gradual increase, reaching a maximum at 8 h, which was 4.7 times that of the control, and then the expression decreased (Fig. 11d).
Discussion
Calcium is involved in signal transduction reactions under various environmental stimuli. As a calcium sensor, CBL proteins typically form a complex with their target kinase CIPK protein to modulate plant responses to abiotic stress (Kudla et al., 2010). CBL-CIPK signaling networks have been studied in many plants, such as Arabidopsis, rice, peas, pear, and wheat (Kolukisaoglu et al., 2004; Mahajan et al., 2006; Sun et al., 2015; Tang et al., 2016). However, there are few studies about CBL-CIPK proteins in tea plants.
All CBL proteins contain four EF-hand motifs, the EF-hand domain is less conserved and may affect functional diversity, and the linker between each EF motif is absolutely conserved among CBL proteins (Zhang et al., 2008). In this study, 7 CBL and 18 CIPK proteins were identified based on the genome data of tea plant. Phylogenetic analysis indicated that they are highly homologous to Arabidopsis proteins (Fig. 1). The CIPK proteins have been shown to contain two domains: an N-terminal kinase catalytic domain and a C-terminal regulatory domain carrying a NAF/FISL motif and a PPI motif. The NAF/FISL motif is required for the interaction between CIPK and CBL proteins (Guo et al., 2001). All CsCIPKs identified from tea plant have a NAF domain in the C-terminal region. It can be seen that the phylogenetic relationship is closer; the types and locations of their motifs are more similar (Figs. 2 and 4). Some motifs are only present in a partial CBL or CIPK amino acid sequence. These findings suggested that these motifs may be involved in functional diversity of the protein. The motif near the N-term is highly conserved, which may be related to the N-terminal kinase catalytic domain of CIPK. The distribution of the conserved motifs indicated that CIPK protein functional characteristics may be mainly determined by the N-terminal conserved domain (NAF), and that C-terminal region may also be involved in determining the functions of these proteins.
The transcripts of eukaryotic gene structures are usually subdivided into two regions, exons and introns, which is a key feature. The intron region is excised during post-transcriptional processing, while the exon region is ultimately retained in the mature mRNA molecule (Matera and Wang, 2014). Gene structure analysis showed that CsCBLs have more introns (Fig. 6a). CsCIPKs can be divided into two types: no introns and rich introns (Fig. 6b). Similar intron-rich/poor CIPK family member patterns were also observed in monocotyledonous and dicotyledonous species, such as Arabidopsis, rice, poplar, soybean, maize, and cassava (Roy and Penny, 2007; Chen et al., 2011; Zhu et al., 2016). These results indicated that CIPK may be related to the product of gene duplication. Thus, the expansion of the CIPK family relies on intron loss of gene replication along the ancient multiple intron CIPK lineages. Whether the genes on the same branch have similar functions remains to be further studied.
Proteins can interact through protein–protein interactions, as a network of functional contacts between proteins and other proteins (de la Rubia et al., 2010). A network of the CBL-CIPK between tea plant and Arabidopsis was constructed. CsCBL4, CsCIPK2, and CsCIPK14 are similar with SOS3 and its kinase SIP3 and SIP4, which belong to the SOS pathway. SIP3, SIP4, and SOS3 play essential roles in salt tolerance in Arabidopsis (Ye et al., 2013; Sardar et al., 2017). CsCBL4, CsCIPK2, and CsCIPK14 proteins may interact with each other in the salt response pathway of tea plant. It is predicted that there are interactions between multiple proteins in tea plants, including interaction between CsCBL4, 6 and CsCIPK8, 14. CsCIPK9, 11 and 12 may interact with CsCBL7. CsCIPK5 and CsCIPK7 may interact with CsCBL1. Protein–protein interactions need to be further verified experimentally in the future.
cis-Regulatory element (CRE) is a region of noncoding DNA. It normally regulates gene transcription by binding to a transcription factor. A single transcription factor can bind to many CREs, thus controlling the expression of many genes. The CREs, such as W-box (WRKY TFBS), WUN-motif (wound-responsive element), MBS (MYB binding site involved in drought inducibility), ABRE (ABA-responsive element), and LTR (low-temperature responsive element), have been extensively characterized for their important roles under stress conditions (Yamaguchi-Shinozaki and Shinozaki, 2005; Hernandez-Garcia and Finer, 2014). The promoters of these 25 genes contain many MBS, ABRE, and W-box repeat elements, indicating that these genes are under pressure regulation. In addition, the promoters of these genes contain many MYB recognition site elements (MBS). Among these elements, the MYB recognition site was found in all the genes examined. The results indicated that the expression of structural genes may be regulated by the CsMYB gene.
In addition, the interactions between CIPK and CBL in Arabidopsis have demonstrated response to different environmental stresses such as abscisic acid, cold, high pH, high salt, and osmotic stress. The CBL9–CIPK3–ABR1 pathway regulates seed germination and ABA-dependent physiological processes in Arabidopsis (Sanyal et al., 2017). CsCIPK10 and CsCIPK18 are homologous to AtCIPK3 and may be involved in the germination of the tea seed. Iron deficiency causes an increase in cytosolic Ca2+ concentration, and an increase in Ca2+ allows it to bind to CBL1/CBL9, resulting in activation of CIPK23. The CBL–CIPK23 complex acts as a sensor for sensing and regulating the homeostasis of minerals in Arabidopsis (Tian et al., 2016). From Figure 8 and Table 1, it was found that CsCIPK9, CsCIPK11, and CsCIPK12 all are homologous to AtCIPK23, which may be related to gene duplication. Plants are particularly susceptible to polyploidization and enhance their gene pool (Rensing, 2014). Overexpression of OsCIPK3, OsCIPK12, and OsCIPK15 did significantly improve rice tolerance to cold, drought, and salt stress, respectively. CsCIPK5 is genetically close to OsCIPK3, but CsCIPK5 is not sensitive to low temperature, and drought and salt stress will increase its expression. This indicated that the CIPK gene does participate in the response to abiotic stress. The degree of response may be different due to differences between plants. These data showed the complexity of plant signaling pathways in response to abiotic stress. In view of the complexity of plant stress regulation, predictions based on bioinformatics analysis require further experimental studies to validate the regulatory pathways and mechanisms of CsCBL and CsCIPK in tea plants.
CsCBLs and CsCIPKs are highly expressed under high-temperature, low-temperature stress, salt stress, and drought stress. The trends of several genes under stress treatment were similar, indicating that there is genetic redundancy in tea plant. There may be interactions between different CsCBLs or CsCIPKs, and various CsCBLs and CsCIPKs may be involved in the signal transduction reactions of these stresses.
Conclusion
Plants are often exposed to environmental stress to meet these challenges, and have evolved a variety of complex molecular mechanisms to survive. These mechanisms involve the induction to stress responses and stress tolerance factors. CBL-CIPK is one of the common stress resistance mechanisms. In this study, we conducted a genome-wide survey of the CBL-CIPK family in tea plants. A total of 7 CsCBLs and 18 CsCIPKs were identified. Then, the physicochemical properties and phylogeny were analyzed. It was found that CIPKs were divided into two branches, and multiple CsCIPKs are closely related to one AtCIPK. Conserved motifs were analyzed, and it was found that the types of motifs between sequences did not change much. By analyzing the gene structure, we found that CsCIPK can be divided into multiple introns and no introns. Similar intron/poor CIPK family member patterns are also found in other monocotyledonous and dicotyledonous species. In combination with TFBSs, a large number of elements and sites related to stress were found, indicating that CsCBL and CsCIPK may be involved in abiotic stress responses. The protein–protein interaction network of CBL-CIPK between tea plant and Arabidopsis was analyzed to explore the possible protein interaction network in tea plants. The expression of CsCBL and CsCIPK in different tissues of tea plants was analyzed, and most of the genes were found to be highly expressed in flowers. Finally, we selected eight genes to study gene expression profiles under abiotic stress, indicating that CsCBL and CsCIPK participate in abiotic stress responses. This study provides a basis for future studies on the CBLs and CIPKs of tea plant response to abiotic stress.
Footnotes
Acknowledgment
The research was supported by the National Natural Science Foundation of China (31570691; 31870681).
Authors' Contributions
J.Z. and H.L. conceived and designed the experiments. H.L., Y.-X.W., H.L., R.-M.T., and Y.W. performed the experiments. H.L., Y.-X.W., and H.L. analyzed the data. J.Z. contributed reagents/materials/analysis tools. H.L. wrote the paper. J.Z. and Y.-X.W. revised the article. All authors read and approved the final article.
Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Supplementary Table S7
Supplementary Table S8
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.