
Abstract
Increasing evidence has indicated that long noncoding RNAs (lncRNAs) could participate in diverse cancers. Among these, lymphoid enhancer-binding factor 1 antisense RNA 1 (LEF1-AS1) was recently identified as an oncogenic lncRNA, but little is known about its function in non-small-cell lung cancer (NSCLC). In the present study, we found that LEF1-AS1 was markedly upregulated in lung cancer tissues and could promote NSCLC cell proliferation and migration in vivo and in vitro. LEF1-AS1 could bind with miR-489 and further negatively regulate miR-489 to promote SRY-related HMG box transcription factor 4 (SOX4) expression. In conclusion, these data suggested that LEF1-AS1 promoted NSCLC tumorigenesis dependent on the miR-489-SOX4 axis and implicated the potential application of LEF1-AS1 for the prognosis and treatment of NSCLC.
Introduction
Lung cancer is the most common malignant disease and represents the leading cause of cancer-related deaths worldwide (Mahmood et al., 2017; Yang et al., 2017). Furthermore, non-small-cell lung cancer (NSCLC) accounts for vast majority of all lung cancers in patients (Yang et al., 2017). Some small molecule kinase inhibitors and immunotherapies, such as checkpoint-related therapies, have significantly improved the outcomes of NSCLC patients. However, the disease still remains incurable once metastatic, and a great number of the patient population needs effective treatments. Thus, studies regarding the underlying mechanisms contributing to the proliferation and metastasis of NSCLC is essential for identifying potential therapeutic targets and developing alternative NSCLC treatments.
Recently, many studies have characterized the regulatory effects that ncRNAs may have on cancer progression. These ncRNAs are closely related to the occurrence, development, metastasis, and drug resistance of cancer. Among these functional ncRNAs, microRNAs (miRNAs) and long non-coding RNAs (lncRNAs) have attracted most attention. lncRNA is a heterogeneous class of transcripts with minimum length of 200 bases and limited protein-coding potential (Schmitz et al., 2016). lncRNAs are known to regulate biological progression through diverse molecular mechanisms, acting as scaffolds to regulate the interactions between proteins and genes, serving as decoys to bind proteins or miRNAs, and as a guide for chromatin modifiers (Karreth and Pandolfi, 2013; Morris and Mattick, 2014).
The aberrant expression of lncRNAs have been reported to contribute to the development of multiple tumors by regulating cell proliferation, apoptosis, metastasis, and invasion (Philip et al., 2013; Vance et al., 2014; Malouf et al., 2015). Along with the evolution of cancer research, many cancer-specific lncRNAs have been found participating in cancer progression, such as HOX transcript antisense RNA (HOTAIR), metastasis-associated long antisense transcript 1 (MALAT-1), antisense noncoding RNA in the INK4 locus (ANRIL), and HOXA transcript at the distal tip (HOTTIP). These abovementioned lncRNAs were found upregulated in lung cancer tissues, their higher expression was correlated with higher TNM stages, and poorer overall survival of patients (Loewen et al., 2014; Deng et al., 2015; Feng-Qi et al., 2015; Zhang et al., 2015a).
Although the increasing roles of lncRNAs involved in the NSCLC process have been identified in the past decade, new therapeutic and diagnostic targets for NSCLC still need further investigation.
In the present study, we proved that lymphoid enhancer-binding factor 1 antisense RNA 1 (LEF1-AS1), a newfound lncRNA, could exert oncogenic activity in NSCLC cells. In detail, we determined that LEF1-AS1 was upregulated in NSCLC and could act as competitive endogenous RNAs (ceRNAs) to promote SRY-related HMG box transcription factor 4 (SOX4) expression through the competitive regulation of miR-489 to facilitate NSCLC cell progression and metastasis.
Materials and Methods
Patient tumor samples
A total of 48 paired NSCLC tissues and adjacent nontumor tissues were obtained from patients who had undergone surgery at the Second Affiliated Hospital of Fujian Medical University (Quanzhou, China) during 2016–2017. After resection, all the tissue samples were collected, immediately snap frozen in liquid nitrogen, and stored at −80°C for subsequent RNA extraction. The study was approved by the Research Ethics Committee of Fujian Medical University (FMU2016-062-16). The written informed consent was obtained by every patient enrolled in the study. The clinical features of the patients are listed in Table 1 and Supplementary Table S1.
Association Between LEF 1-AS 1 Expression and Clinicopathological Characteristics
TNM, tumor–node–metastasis staging system.
p < 0.05 two-sided χ2 test.
Cell culture and cell transfection
The human NSCLC cell lines, H1299 and A549, used in this study were obtained from American Type Culture Collection (ATCC). All the cells were cultured at 37°C in humidified air atmosphere containing 5% carbon dioxide in DMEM (Gibco) supplemented with 10% fetal bovine serum (Gibco), 100 U/mL penicillin, and 100 μg/mL streptomycin. Cell transfection was performed using Lipofectamine 2000 (Invitrogen) according to the manufacturer's instructions. miR-489 mimic, SOX4 siRNAs, LEF1-AS1 overexpression vector, and Lentivirus were bought from RiboBio (Guangzhou, China).
RNA isolation and qRT-PCR
Total RNA from the NSCLC tissues and cell lines in this study were extracted with TRIzol Reagent (Invitrogen) according to the manufacturer's protocol. First-strand cDNA was synthesized using the PrimeScript RT Reagent Kit (TaKaRa, Japan). Real-time PCR was conducted using SYBR green (TaKaRa). GAPDH was used as a housekeeping gene for baseline expression.
Flow cytometry
The apoptotic cells were measured using the Apoptosis Detection Kit (KeyGen Biotech, China). Briefly, after transfection for 48 h, cells were collected and washed with PBS three times, followed by staining with Annexin V (AV) and Propidium Iodide in the dark. The apoptotic cells were tested using flow cytometer (FACScan). Experiment was repeated at least three times.
Subcellular fractionation
The separation of the nuclear and cytosolic RNA was performed using the PARIS Kit (Life Technologies) according to the manufacturer's instructions. Cells were incubated with hypotonic buffer for 5 min and 1% NP-40 was added. After centrifuging at 5000 g for 5 min, the supernatant was collected as the cytosolic fraction. Then the pellets were suspended in nuclear suspension buffer and incubated at 4°C for 30 min. Finally, the nuclear fraction was collected after removing insoluble membrane debris by centrifuging at 12,000 g for 10 min.
Cell proliferation assay
Cell proliferation was detected using the CCK-8 reagent (Roche Diagnostics, Japan). Briefly, 3 × 103 cells were seeded in 96-well plates and incubated for 96 h at 37°C, the CCK-8 reagent was added at different time points and incubated for further 2 h. Absorbance was measured using a Microplate Reader (Bio-Tek) at test wavelengths of 450 nm.
Transwell assays
The transwell (BD Biosciences) assay was used to test cell migration and invasion abilities. A total of 2 × 105 cells were suspended in serum-free medium and plated into the upper chamber. In the meanwhile, complete medium was added to the lower chamber of the inserts. After incubation for 24 h (migration assay) or 48 h (invasion assay) at 37°C in a humidified incubator with 5% CO2, the inserts were fixed with 4% formaldehyde and cells were stained with Crystal Violet and then photographed and quantified by counting the cell numbers in five random fields.
Western blot assay
Total cell proteins were extracted from different cells with RIPA lysis buffer and protein concentrations were determined using the BCA Protein Assay Kit (Beyotime, China). Antibodies against SOX4 (Abcam), Bcl-2 (Abcam), Bax (CST), Caspase-3 (Abcam), E-cadherin (CST), N-cadherin (CST), Vimentin (Abcam), Fibronectin (CST), and GAPDH (Abcam) were used overnight at 4°C. After incubating with horseradish peroxidase-conjugated secondary antibodies (CST) for 1 h at room temperature, it was followed by blotting with an enhanced chemiluminescence system (Pierce) according to the manufacturer's protocol.
Luciferase reporter assay
LEF1-AS1 sequence containing the potential miR-489-binding sites or mutants of each binding site were constructed into pmirGLO. Similarly, the predicted miR-489 response element (wild-type or mutant) in the 3′-UTR of SOX4 was cloned into pmirGLO plasmid. Vectors and miR-489 mimics were cotransfected, and luciferase activity was measured using the Dual Luciferase Assay System (Promega) following the manufacturer's instructions. The experiment was performed in duplicate in three independent experiments.
RNA-binding protein immunoprecipitation assay
According to the manufacturer's protocol, RNA immunoprecipitation assays were conducted using the RIP Assay Kit (Millipore Corp.). Protein–RNA complexes were isolated, and anti-Ago2 or IgG was added to the reaction system for immunoprecipitation. After RNA purification, the immunoprecipitated RNA was detected and the expression levels of LEF1-AS1 and miR-489 were verified by qPCR. IgG served as a negative control.
Animal assays
To evaluate the effect of LEF1-AS1 on tumor growth, subcutaneous xenograft mouse model was adopted. Athymic nude mice (BALB/c, 4 weeks old, n = 4 per group) were obtained from the Vital River Laboratories (Beijing, China) and animal studies were approved by the Animal Care and Use Committee of Fujian Medical University (FMU-ACUC18-068). A total of 2 × 106 cells were injected into the flank of each mouse. After inoculation for 24 days, mice were sacrificed and tumors were excised, weighed, and processed for further experimentation. Tumor size was determined using measurements of the shortest diameter (a) and the longest diameter (b) with a caliper. The volume was calculated using the formula V = (a2b)/2.
For metastasis experiments, A549 cells were stably transfected with vector lentivirus or LEF1-AS1 recombinant lentiviruses. Around 1 × 106 of the abovementioned cells were injected into the tail vein of mice (n = 5 for each group). After injection, metastases were examined by bioluminescence imaging (BLI) using an IVIS Spectrum Xenogen Imaging System (Xenogen). After scanning, intact lungs was isolated for counting metastasis nodules and HE staining.
Statistical analysis
Data were presented as the mean ± SD. All data were analyzed with GraphPad Prism 5.0 software. In all cases, p-value less than 0.05 indicated that data were statistically significant. Differences between two groups or among more than two groups were separately analyzed with Student's t-test or one-way ANOVA.
Results
lncRNA LEF1-AS1 is upregulated in NSCLC tissues and is associated with poor prognosis
To determine whether lncRNA LEF1-AS1 was associated with the progression of NSCLC, we analyzed that RNA-Seq data (from The Cancer Genome Atlas [TCGA]) of lncRNAs of NSCLC were from TANRIC (Li et al., 2015) (
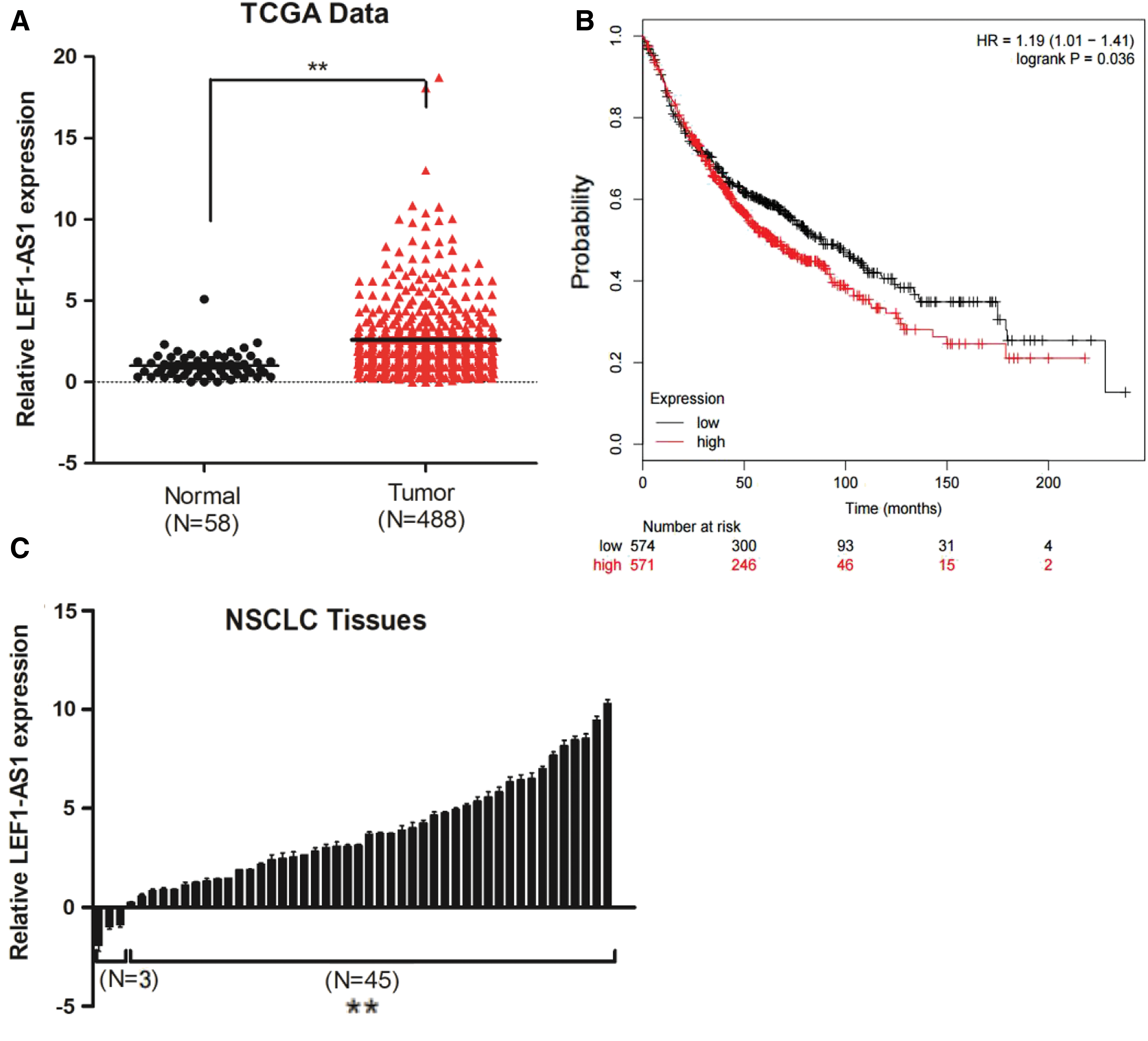
lncRNA LEF1-AS1 is upregulated in NSCLC tissues and is associated with poor prognosis.
To further explore the clinical significance of LEF1-AS1, we analyzed LEF1-AS1 RNA levels in 48 pairs of human NSCLC tissues. Compared with corresponding noncancerous tissues, LEF1-AS1 was significantly upregulated in NSCLC tissues (Fig. 1C). Next, to assess the correlation of LEF1-AS1 expression with clinicopathological characteristics, the 48 tumor tissues were divided into low (n = 24) or high (n = 24) groups using the median expression of LEF1-AS1 as a cutoff. Correlation regression analysis showed that the LEF1-AS1-high group was significantly correlated with tumor size (p = 0.037) and TNM stage (p = 0.04) (Table 1). These results indicated that LEF1-AS1 was upregulated in NSCLC tissues, which gave further evidence that it may play a pivotal role in the progression of human NSCLC.
LEF1-AS1 promoted NSCLC cell proliferation and migration in vivo and in vitro
To investigate the potential role of LEF1-AS1 in the pathogenesis of NSCLC, LEF1-AS1 overexpression plasmid was used to exogenously manipulate the expression of LEF1-AS1 in both the A549 and H1299 cell lines. As shown in Figure 2A, the expression levels of LEF1-AS1 were significantly increased in LEF1-AS1-pcDNA3.1-transfected cells compared with the empty pcDNA3.1-transfected counterparts. LEF1-AS1 overexpression significantly increased the proliferative ability (Fig. 2B, C) and restrained the apoptosis rate in comparison with control cells (Fig. 2D–F). In subcutaneous xenograft assay, LEF1-AS1 overexpression significantly induced the proliferation of A549 cells in mice (Fig. 2G, H), which was consistent with the in vitro data.
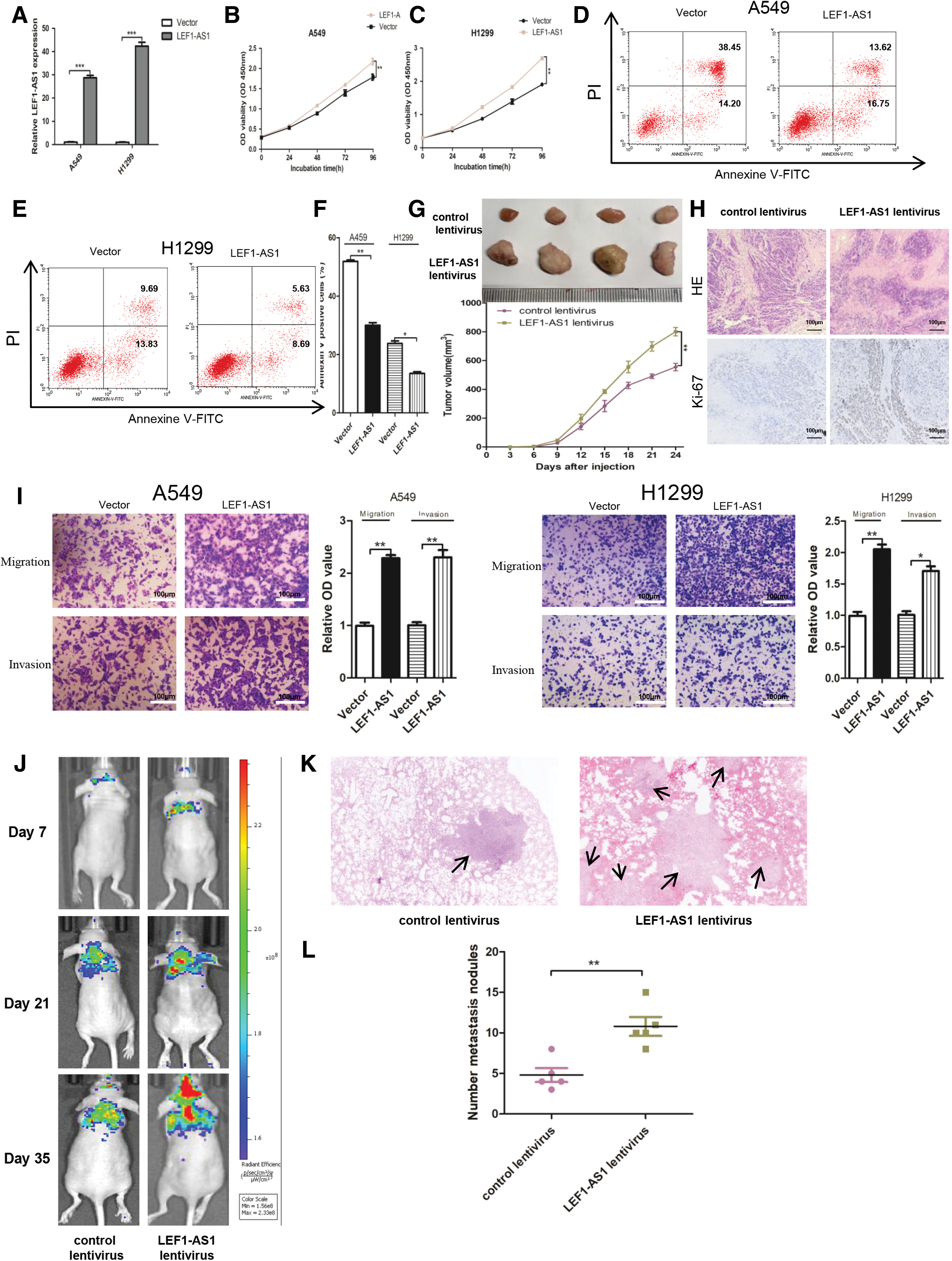
LEF1-AS1 promoted NSCLC cell proliferation and migration in vivo and in vitro.
Moreover, we found that LEF1-AS1 overexpression markedly promoted cell migration and invasion in both the A549 and H1299 cells (Fig. 2I). Then mouse models were adopted to evaluate the in vivo metastasis efficacy of LEF1-AS1. As shown in Fig. 2J, BLI scans were taken at intervals from day 7 to 35 after tail vein injection. The analysis of the BLI scans revealed that LEF1-AS1 in A549 cells promoted lung metastasis. More metastatic nodules were detected in the mice injected with LEF1-AS1-overexpressed A549 cells group compared with those of the control group, which were confirmed using pulmonary HE staining (Fig. 2K, L). All the data revealed that LEF1-AS1 served as an oncogenic driver in the pathogenesis of NSCLC.
LEF1-AS1 facilitated SOX4 expression by sponging miR-489
As the dominant function of lncRNA is related to cellular localization, we analyzed its cellular distribution by qRT-PCR and found that LEF1-AS1 was mainly localized in the cytoplasm of A549 and H1299 cells through a cellular fractionation assay (Fig. 3A). We then hypothesized that LEF1-AS1 may function as ceRNA by competitively binding miRNAs. Through bioinformatics analysis, we screened miR-489 as a potential target of LEF1-AS1. To verify the binding site of LEF1-AS1 and miR-489, we cloned the LEF1-AS1 wild or mutant fragment into luciferase reporter plasmids and cotransfected these plasmids with miR-NC or miR-489 mimics into A549 cells. Results found that the luciferase intensity was decreased by the cotransfected miR-489 mimics and LEF1-AS1 wild plasmid but not the mutant reporter vector, which lacked the putative binding site (Fig. 3B).
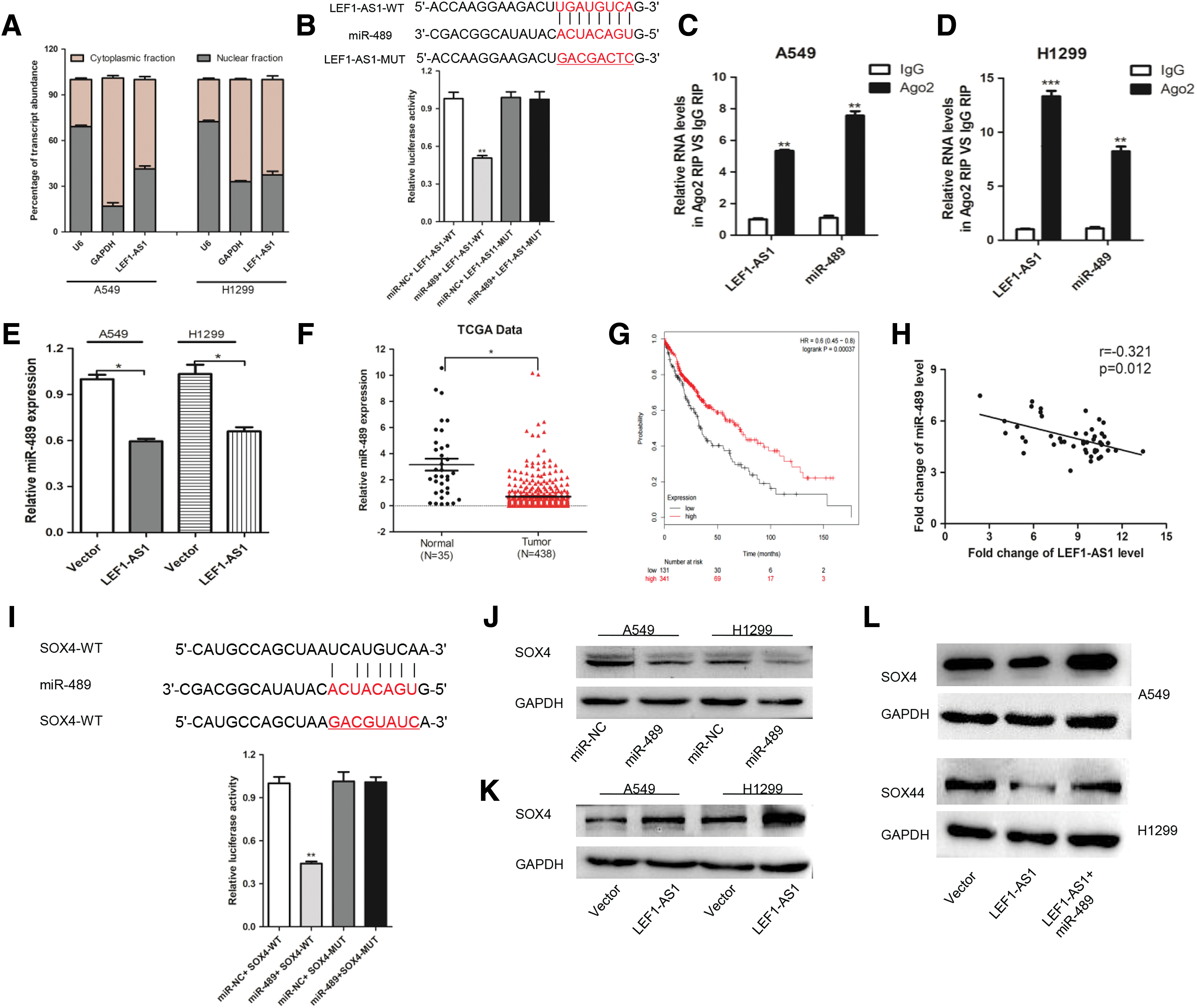
Next, to further confirm the interaction between LEF1-AS1 and miR-489, RNA immunoprecipitation (RIP) assays were conducted on A549 and H1299 cells using an antibody against Ago2, the core component of the RNA-induced silencing complex. The results showed that LEF1-AS1 and miR-489 were both enriched in Ago2 immunoprecipitation relative to the control IgG (Fig. 3C, D). Intriguingly, miR-489 was downregulated by LEF1-AS1 (Fig. 3E). Data analysis found that miR-489 was also downregulated in NSCLC tissues compared with the adjacent normal tissues (Fig. 3F). Moreover, Kaplan–Meier curve showed that a low miR-489 expression in NSCLC tissues was significantly associated with worse overall survival (Fig. 3G). As shown in Figure 3H, a negative expression correlation between LEF1-AS1 and miR-489 was detected in the NSCLC samples. All of the abovementioned data hinted us that miR-489 may serve as a potential target for LEF1-AS1 exerting its carcinogenic effect.
To further elucidate the mechanism underlying the contribution of the LEF1-AS1-miR-489 axis to NSCLC progression, TargetScan was used to predict the potential target genes of miR-489 and found that SOX4 was a putative target mRNA of miR-489 (Fig. 3I). Then, we transfected the miR-489 mimic into A549 and H1299 cells and found that the SOX4 protein level was significantly decreased (Fig. 3J). Correspondingly, the protein levels of SOX4 were proportional to the expression levels of LEF1-AS1 (Fig. 3K). Finally, a western blot assay was conducted to determine whether LEF1-AS1 could regulate SOX4 expression through miR-489. As shown in Figure 3L, the protein level of SOX4 was increased upon LEF1-AS1 transfection, whereas miR-489 could counteract the stimulating effect of LEF1-AS1. Collectively, these data demonstrated that LEF1-AS1 could function as a molecular sponge for miR-489 to regulate SOX4 expression.
LEF1-AS1 targeted the miR-489/SOX4 axis to promote NSCLC tumorigenesis
The abovementioned results demonstrated that LEF1-AS1 could compete with miR-489 and increase the expression of SOX4, which prompted us to verify whether LEF1-AS1 exerted its oncogenic function through the miR-489/SOX4 axis. The results showed that LEF1-AS1-induced proliferation and antiapoptosis effects were compromised by the transfection of miR-489 mimics or SOX4 siRNA (Fig. 4A–C). Next, migration and invasion abilities were also measured in A549 and H1299 cells, and the results further confirmed that either overexpressing miR-489 or inhibiting SOX4 could repress the increasing cellular migration and invasion in NSCLC cells transfected with LEF1-AS1 (Fig. 4D, E). Consistent with the abovementioned results, a western blot assay further demonstrated that LEF1-AS1 induced EMT progression through the miR-489/SOX4 axis (Fig. 4F). In summary, we concluded that LEF1-AS1 exerted its oncogenic ability dependent on the miR-489/SOX4 axis.
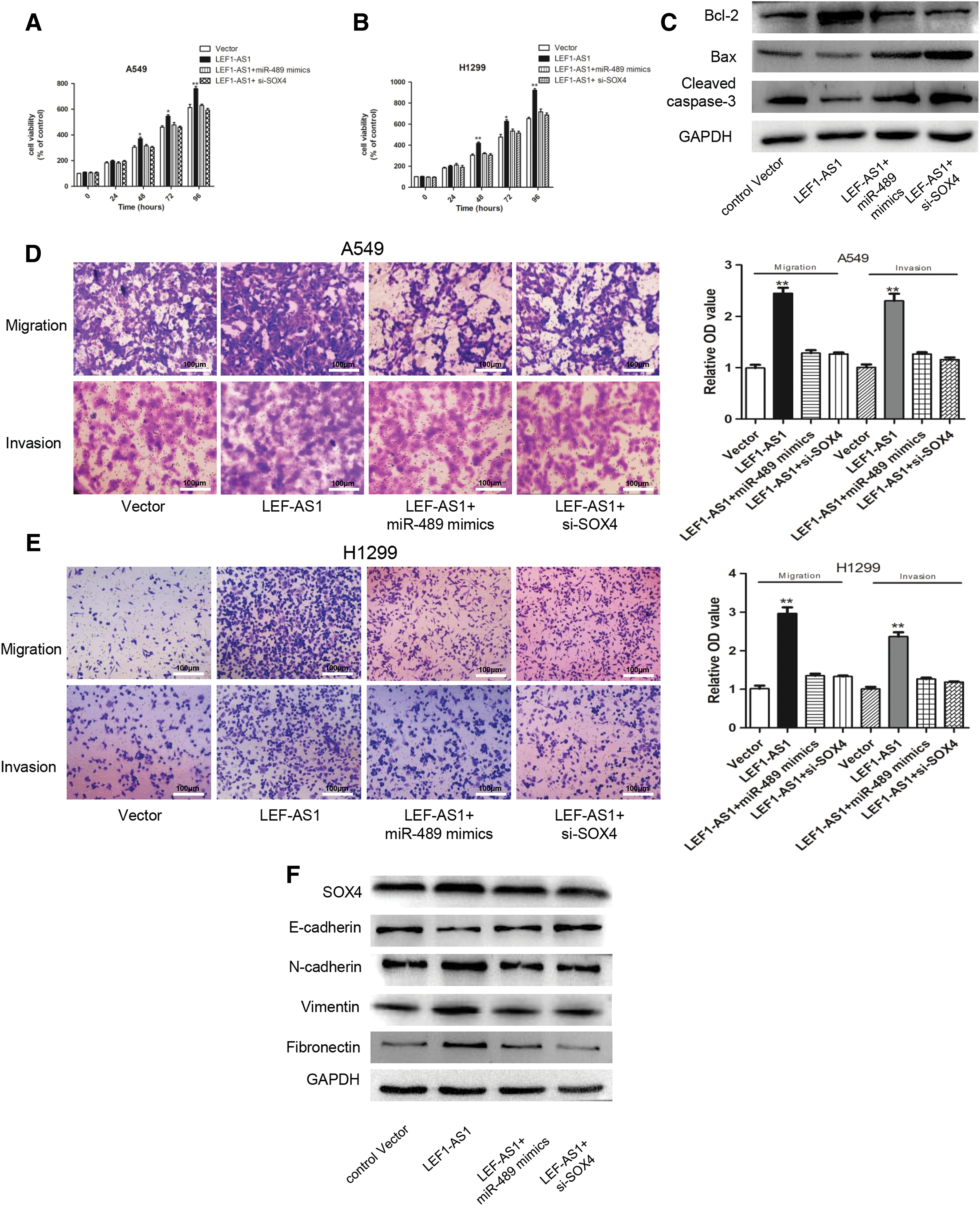
LEF1-AS1 targeted the miR-489/SOX4 axis to promote NSCLC tumorigenesis.
Discussion
Over the past decade, despite tremendous advances in tumor diagnosis and the treatment of NSCLC, the prognosis for broader-scale NSCLC patients is still short of effective therapies. Hence, it is important to illustrate the underlying mechanisms of NSCLC development and find new potential drug targets. Emerging evidence has indicated the regulatory role of lncRNAs in many cancers, including NSCLC (Li and Chen, 2013; Tian et al., 2017). The dysregulated lncRNAs participated in tumor progression and metastasis (Feng-Qi et al., 2015; Zhang et al., 2015b) and have acted as biomarkers in cancer diagnosis and prognosis (Qiu et al., 2015).
lncRNAs can exert their functions through diverse modes, among which acting as ceRNAs to abrogate the inhibitory effect of miRNAs on their target genes has been reported by a growing number of reports. For example, TGF-β-induced lnc-ATB could directly bind with miR-200s and regulate ZEB1 and ZEB2 to induce EMT and promote invasive phenotype in hepatocellular carcinoma (Fan et al., 2017). Another lncRNA, UCA1, was found to be significantly upregulated in NSCLC tissues, and the overexpression of UCA1 remarkably facilitated cell proliferation and colony formation through the UCA1/miR-193a-3p/ERBB4 axis regulatory network in NSCLC (Nie et al., 2016).
With the help of substantial advances in microarray and sequencing techniques, many dysregulated lncRNAs have been identified and proven to play profound roles on multiple cancers. Among those newfound lncRNAs, LEF1-AS1 was first identified to be upregulated in glioblastoma (GBM) patients and studies found that lncRNA LEF1-AS1 could act as an oncogene to promote GBM cell proliferation, migration, and invasion partially through regulating the ERK and Akt/mTOR pathways (Wang et al., 2017). Moreover, a growing number of studies showed that LEF1-AS1 promoted prostate cancer progression by competing with miR-330-5p and increased LEF1 expression. In another study, LEF1-AS1 contributed to progression through regulating miR-544a/FOXP1 axis in lung cancer, which was consistent with our study that LEF1-AS1 acted as an oncogenic driver of NSCLC, but in a different manner (Wang et al., 2017).
Considering the known findings as mentioned above, we then speculated whether LEF1-AS1 could also exert similar functions in NSCLC. In the present study, we identified that LEF1-AS1 was upregulated in NSCLC tissues and was associated with poor patient survival. The overexpression of LEF1-AS1 remarkably promoted NSCLC cell proliferation and migration in vitro and in vivo. As LEF1-AS1 mainly localized in the cytoplasm, we tried to explore potential binding miRNAs for LEF1-AS1. A low expression of miR-489 was observed in many cancers, and an miR-489 loss promoted proliferation, migration, and invasion in lung cancer cells (Xie et al., 2015). Consistent with previous reports, we found that the expression of miR-489 was both downregulated in NSCLC tumor tissues from the TCGA database and in clinical specimens. Moreover, the expression of miR-489 was inversely proportional to the expression of LEF1-AS1.
Next, we confirmed that SOX4 was a new target of miR-489. The high expression of SOX4 was reported in many cancers, and SOX4 activation boosts tumor development and progression, such as the inhibition of apoptosis, the induction of cell migration, and metastasis (Vervoort et al., 2013; Lourenço and Coffer, 2017). Moreover, it was demonstrated that SOX4 contributes to tumorigenesis by controlling EMT, which was consistent with the function of LEF1-AS1. Further studies demonstrated that the promotion of proliferation and metastasis of LEF1-AS1 activation was restrained by overexpressing miR-489 or downregulating SOX4.
However, there are still limitations in our present study. The expression of LEF1-AS1 was negative related to lymph node metastasis in our clinical data, which was not in accordance with promoting metastasis ability of LEF1-AS1. We think this contradiction was partly due to the heterogeneity of tumor tissues and the small sample size (Zhang et al., 2016). The prospective, multicenter cohort study with large samples is needed for us to better understand the role of LEF1-AS1 in NSCLC.
Conclusion
In summary, we found that LEF1-AS1 was overexpressed in NSCLC tissues compared with matched normal tissues and was inversely correlated with miR-489 expression in NSCLC tissues. Next, we demonstrated that LEF1-AS1 negatively regulated miR-489 to promote SOX4 expression in NSCLC cells. Taken together, the LEF1-AS1/miR-489/SOX4 signaling axis likely exerted key functions in the proliferation and metastasis of NSCLC. Furthermore, our study provided mechanistic insight into the oncogenic roles of LEF1-AS1 and the pivotal effects of LEF1-AS1 as a promising therapeutic target for NSCLC patients.
Footnotes
Acknowledgments
The work was funded by the Quanzhou Science and Technology Plan Project (Grant No. 2018Z101).
Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.