
Abstract
Although long noncoding RNAs (lncRNAs) and circular RNAs (circRNAs) have been suggested to play important roles in the pathogenesis of diseases, atherosclerosis-related lncRNAs and circRNAs remain rarely reported. This study aimed to explore the underlying molecular mechanisms of atherosclerosis based on the competing endogenous RNA (ceRNA) regulatory hypothesis of lncRNAs and circRNAs. The expression profiles of circRNAs, lncRNAs, and mRNAs in human THP-1 macrophages treated with oxidized low-density lipoprotein (an in vitro atherosclerosis model), or not, were obtained from the Gene Expression Omnibus database under accession numbers GSE107522, GSE54666, and GSE54039, respectively. The present study identified 29 differentially expressed circRNAs in GSE107522, 544 differentially expressed genes (DEGs) in GSE54666, and 502 DEGs and 231 differentially expressed lncRNAs in GSE54039 datasets by using the Linear Models for Microarray Data method. Eight DEGs were found to be shared and expressed with the consistent trend in GSE54666 and GSE54039 datasets. Two of them (ASPH, aspartate beta-hydroxylase; and PDE3B, phosphodiesterase 3B) were suggested to be crucial based on functional enrichment, protein–protein interaction, and ceRNA network analyses. ASPH, through interaction with CACNA2D4 (calcium voltage-gated channel auxiliary subunit alpha2delta 4), may be associated with atherosclerosis by regulating the cellular response to calcium ion; and PDE3B may exert roles in negative regulation of angiogenesis through cross talk with ELMO1 (engulfment and cell motility 1). Furthermore, the expression of ASPH and PDE3B may be regulated by hsa_circ_0028198/hsa_circ_0092317/XIST-miR-543; PDE3B expression may be also modulated by hsa_circ_0092317/hsa_circ_0003546/H19/XIST-miR-326. In conclusion, our identified ceRNA interaction axes may possibly be important targets for treatment of atherosclerosis.
Introduction
Atherosclerosis is an important risk factor to induce ischemic cardiovascular diseases, including myocardial infarction and stroke, which are two leading causes for mortality and morbidity worldwide (Mathers and Loncar, 2006; Barquera et al., 2015). The hallmark of early atherosclerosis is the formation of foam cells that are resulted from uptake and accumulation of oxidized low-density lipoprotein (ox-LDL) in arterial macrophages (Yu et al., 2013; Delgadomaroto et al., 2017). Thus, how to prevent foam cell formation and slow the progression of arteriosclerotic processes has been a rising public concern.
Recently, emerging evidence has suggested that noncoding RNAs (ncRNAs), including microRNAs (miRNAs), long noncoding RNAs (lncRNAs), and circular RNAs (circRNAs), play crucial roles in development of various diseases (Manel, 2011; Beermann et al., 2016). miRNAs negatively regulate the expression of mRNAs of protein-coding genes by binding to the complementary sequences located in the 3′-untranslated region (UTR) (Hausser and Zavolan, 2014); lncRNAs/circRNAs could function as competing endogenous RNAs (ceRNAs) to competitively bind with miRNAs through their miRNA response elements and then indirectly regulate the expression of miRNA-targeted mRNAs (Salmena et al., 2011). Therefore, lncRNA/circRNA-miRNA-mRNA interaction may be an important mechanism for initiation and development of atherosclerosis. This hypothesis has been preliminarily demonstrated by the following studies: Ye et al. (2019) identified that upregulated lncRNA myocardial infarction associated transcript (MIAT) could sponge miR-149-5p and prevent the inhibition effects of miR-149-5p on CD47, leading to upregulation of the antiphagocytic molecule, CD47, in advanced atherosclerosis. Knockdown of MIAT attenuated the progression of atherosclerosis, reduced the necrotic core size, increased plaque stability in vivo, and promoted the clearance of apoptotic cells by macrophages in vitro (Ye et al., 2019). Yan et al. (2017) found that ox-LDL increased MEG3 expression in Raw264.7 macrophage cells. MEG3 could sponge miR-204, while miR-204 could regulate the expression of cyclin-dependent kinase inhibitor 2A in Raw264.7 cells treated with ox-LDL. By RNA-Seq analysis and ceRNA network construction, Zhang et al. (2018a) identified ocu-cirR-novel-18038, -18298, -15993, -17934, -17879, -18036, and -14389 as crucial circRNAs related to atherosclerosis. However, atherosclerosis-related lncRNA/circRNA-miRNA-mRNA ceRNA regulatory mechanisms remain rarely reported until now, and no lncRNAs and circRNAs that interact with the same miRNAs and targets were investigated.
The goal of this study was to further screen crucial lncRNA/circRNA-miRNA-mRNA ceRNA axes in ox-LDL-induced foam cells by using microarray data collected from a public database and a series of bioinformatic analysis. The results of the present study may deepen the understanding of the molecular mechanisms of atherosclerosis and provide some targets for developing therapeutic approaches.
Materials and Methods
Downloading public microarray datasets
By searching the National Center for Biotechnology Information (NCBI) Gene Expression Omnibus repository (GEO) database with the key words, ox-LDL and foam cells, three microarray datasets involving human THP-1 macrophages were obtained, including GSE107522, GSE54666 (Reschen et al., 2015), and GSE54039 (Hu et al., 2014). In the GSE107522 dataset, the authors analyzed the circRNA expression profiles in three THP-1 macrophages and three foam cells induced by stimulation of THP-1 macrophages with ox-LDL (50 mg/mL) for 24 h using Agilent-069978 Arraystar Human CircRNA microarray, V1 (platforms: GPL19978). In the GSE54666 dataset, six independent biological replicates, including half of the THP1 macrophages treated with ox-LDL (50 μg/mL) for 48 h to form foam cells and the other half treated with the control buffer, were obtained to detect the mRNA expression profiles using Illumina HumanHT-12, V4.0, Expression BeadChip (platforms: GPL10558). In the GSE54039 dataset, lncRNA/mRNA expression profiling analysis for three THP-1 macrophages and three THP-1 macrophage-derived foam cells (50 μg/mL of ox-LDL treatment for 48 h) was performed using Arraystar Human LncRNA microarray, V2.0 (Agilent-033010 Feature Number version) (platforms: GPL13825).
Differential analysis
The preprocessed expression matrix of each dataset was downloaded from GEO, with the quantile normalization method using the GeneSpring GX, v11.5.1, software package for GSE107522 and GSE54039, while robust spline normalization was performed for GSE54666. The gene differential expression analysis was carried out with the Linear Models for Microarray Data (LIMMA) method software (version 3.34.0) (Ritchie et al., 2015). The differentially expressed genes (DEGs), differentially expressed circRNAs (DECs), and differentially expressed lncRNAs (DELs) were annotated according to the corresponding platform information. A p-value <0.05 and |logFC (fold change)| > 0.5 were defined as statistical threshold values due to the fact that results of the false discovery rate were not perfect. Furthermore, this cutoff point was selected to prevent the rare interaction pairs screened between RNAs. A heatmap was rendered using the R package pheatmap (version: 1.0.8) based on Euclidean distance to visualize the distinguishing ability of DEGs, DECs, and DELs for samples.
Protein–protein interaction network analysis
The Search Tool for the Retrieval of Interacting Genes/Proteins (STRING; version 10.0) database (Szklarczyk et al., 2015) was used to retrieve the protein interactions between DEGs identified in GSE54666 and GSE54039 datasets. Only STRING-derived interactors with high confidence (≥0.700) were imported into the Cytoscape software (version 3.6.1) (Kohl et al., 2011) to construct the protein–protein interaction (PPI) network.
circRNA/lncRNA-miRNA-mRNA ceRNA network construction
The interaction relationships between miRNAs and DEGs were predicted using the miRWalk database (version 2.0), which contained 12 algorithms (Dweep and Gretz, 2015). miRNAs that interacted with DECs were predicted using the CircInteractome database (version 1.0) (Dudekula et al., 2016). miRNAs that interacted with DELs were predicted using the miRcode database (version 1.0) (Jeggari et al., 2012). DEC and DEL-related ceRNA networks were constructed based on the commonly interactive miRNAs with DEGs, DECs, and DELs. The circRNA/lncRNA-miRNA-mRNA ceRNA network was visualized using Cytoscape software (version 3.6.1) (Kohl et al., 2011).
Functional enrichment analysis
Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway and gene ontology (GO) enrichment analyses were done using the Database for Annotation, Visualization and Integrated Discovery (DAVID) online tool (version 6.8) (Huang et al., 2009). Pathways or GO biological process terms with p-values of less than 0.05 were considered to be significantly enriched.
Results
Differential expression analysis
According to the cutoff criteria (p < 0.05 and |logFC| > 0.5), a total of 29 DECs (21 upregulated and 8 downregulated) and 544 DEGs (236 upregulated and 308 downregulated) were, respectively, identified between macrophages treated with and without ox-LDL in GSE107522 and GSE54666 datasets. The GSE54039 datasets included mRNA and lncRNA data, in which 502 DEGs (264 upregulated and 238 downregulated) and 231 DELs (63 upregulated and 168 downregulated) were, respectively, examined when comparing macrophages undergoing ox-LDL treatment with ox-LDL nonstimulation. Furthermore, 12 DEGs were found to be shared in GSE54666 and GSE54039 datasets, among which 8 (FARP1, FERM, ARH/RhoGEF, and pleckstrin domain protein 1; FCGR2B, Fc fragment of IgG receptor IIb; ASPH, aspartate beta-hydroxylase; NAV2, neuron navigator 2; OGFRL1, opioid growth factor receptor-like 1; ST14, suppression of tumorigenicity 14; HMGCS1, 3-hydroxy-3-methylglutaryl-CoA synthase 1; and PDE3B, phosphodiesterase 3B) were expressed with the consistent trend, while 4 (IGF1, insulin-like growth factor 1; DUSP1, dual-specificity phosphatase 1; ATP2B4, ATPase plasma membrane Ca2+ transporting 4; and HS3ST2, heparan sulfate-glucosamine 3-sulfotransferase 2) were opposite in expression trend in 2 databases (Table 1). The hierarchical clustering heatmap visually revealed that THP-1 macrophages and foam cells displayed distinguishably characteristic circRNA (Fig. 1A), mRNA (Fig. 1B, C), and lncRNA (Fig. 1D) expression profiles.

Hierarchical clustering and heatmap analysis of differentially expressed circRNAs
Crucial Differentially Expressed Genes, lncRNAs, and circRNAs Between Macrophages Treated With and Without Oxidized Low-Density Lipoprotein
circRNA, circular RNA; FC, fold change; lncRNA, long noncoding RNA.
Functional enrichment for DEGs
Considering that common DEGs were less in GSE54666 and GSE54039 datasets, KEGG pathway enrichment was performed for DEGs identified in these two datasets, respectively. The results showed that DEGs in GSE54666 were enriched into 20 KEGG pathways, such as hsa04923:Regulation of lipolysis in adipocytes (PDE3B); hsa04921:Oxytocin signaling pathway (CAMK2B, CACNA2D4); and hsa01212:Fatty acid metabolism (ACAT2); while 5 KEGG pathways were obtained for the DEGs in GSE54039 such as hsa05205:Proteoglycans in cancer (CAMK2D) (Table 2).
Kyoto Encyclopedia of Genes and Genomes Pathway Enrichment Genes for Differentially Expressed Genes
IBD, inflammatory bowel disease.
In addition, the GO enrichment study was also performed. The results indicated that DEGs in GSE54666 were enriched into 215 GO biological process terms, such as GO:0071277∼cellular response to calcium ion (ASPH), GO:0010524∼positive regulation of calcium ion transport into cytosol (ASPH), GO:0016525∼negative regulation of angiogenesis (PDE3B), and GO:0006915∼apoptotic process (ELMO1), while 75 GO biological process terms were enriched for the DEGs in GSE54039, also including GO:0016525∼negative regulation of angiogenesis (PDE3B), GO:0071277∼cellular response to calcium ion (ASPH), and GO:0070588∼calcium ion transmembrane transport (ASPH) (Table 3).
Gene Ontology Term Enrichment Analysis for Differentially Expressed Genes
Only top 10 are listed.
GO, gene ontology.
PPI network
When uploading the DEGs identified in both GSE54666 and GSE54039 datasets to STRING, 1217 high-confidence interaction relationships were found to be present in 448 DEGs, which were used to construct the PPI network. In this network, ASPH could interact with CACNA2D4; PDE3B could interact with ELMO1, CAMK2D, and CAMK2B; and HMGCS1 and HMGCR were interacted (Fig. 2), further revealing the underlying upstream or downstream mechanisms for crucial genes.

Protein and protein interaction network for differentially expressed genes. Color images are available online.
ceRNA network construction
There were 585 interaction pairs predicted between 29 DECs and 230 miRNAs by the CircInteractome database; there were 3995 interaction pairs predicted between 56 DELs and 153 miRNAs by the miRcode database; and there were 3208 interaction pairs predicted between 8 common DEGs in 2 datasets and 931 miRNAs by the miRWalk 2.0 database based on at least 6 algorithms. By integrating with the common miRNAs, circRNA-miRNA-mRNA (including 94 miRNAs, 8 DEGs, and 29 DECs) (Fig. 3) and lncRNA-miRNA-mRNA (including 54 miRNAs, 8 DEGs, and 29 DELs) (Fig. 4) regulatory networks were constructed.

ceRNA interaction network of circRNA-miRNA-mRNA. Circular nodes represent circRNAs; regular triangle nodes represent mRNAs; and inverted triangle nodes represent miRNAs. ceRNAs, competing endogenous RNAs. miRNA, microRNA. Color images are available online.

ceRNA interaction network of lncRNA-miRNA-mRNA. Square nodes represent lncRNAs; regular triangle nodes represent mRNAs; and inverted triangle nodes represent miRNAs. Color images are available online.
In the circRNA-related ceRNA network (Fig. 3), ASPH can be targeted by 43 miRNAs, while 41 of them could also interact with 18 upregulated DECs (such as hsa_circ_0028198 and hsa_circ_0092317), resulting in the ceRNA relationship pairs of hsa_circ_0028198/hsa_circ_0092317-miR-543-ASPH; PDE3B can be regulated by 15 miRNAs, all of which could also interact with 17 upregulated DECs (such as hsa_circ_0092317, hsa_circ_0028198, and hsa_circ_0003546), forming the following ceRNA relationships of hsa_circ_0092317/hsa_circ_0003546-miR-326-PDE3B and hsa_circ_0028198/hsa_circ_0092317-miR-543-PDE3B.
In the lncRNA-related ceRNA network (Fig. 4), ASPH can be regulated by 30 miRNAs, while these 30 miRNAs could also interact with 18 upregulated DELs (such as XIST), which constituted the ceRNA axis of XIST-miR-543-ASPH. PDE3B can be regulated by 11 miRNAs; meanwhile, all these miRNAs could also interact with 17 upregulated DELs (such as H19 and XIST), with the ceRNA relationships of MALAT1/XIST-miR-543-PDE3B and H19/XIST-miR-326-PDE3B. These crucial ceRNA axes are shown in Figure 5 after integration of lncRNA and circRNA ceRNA networks.
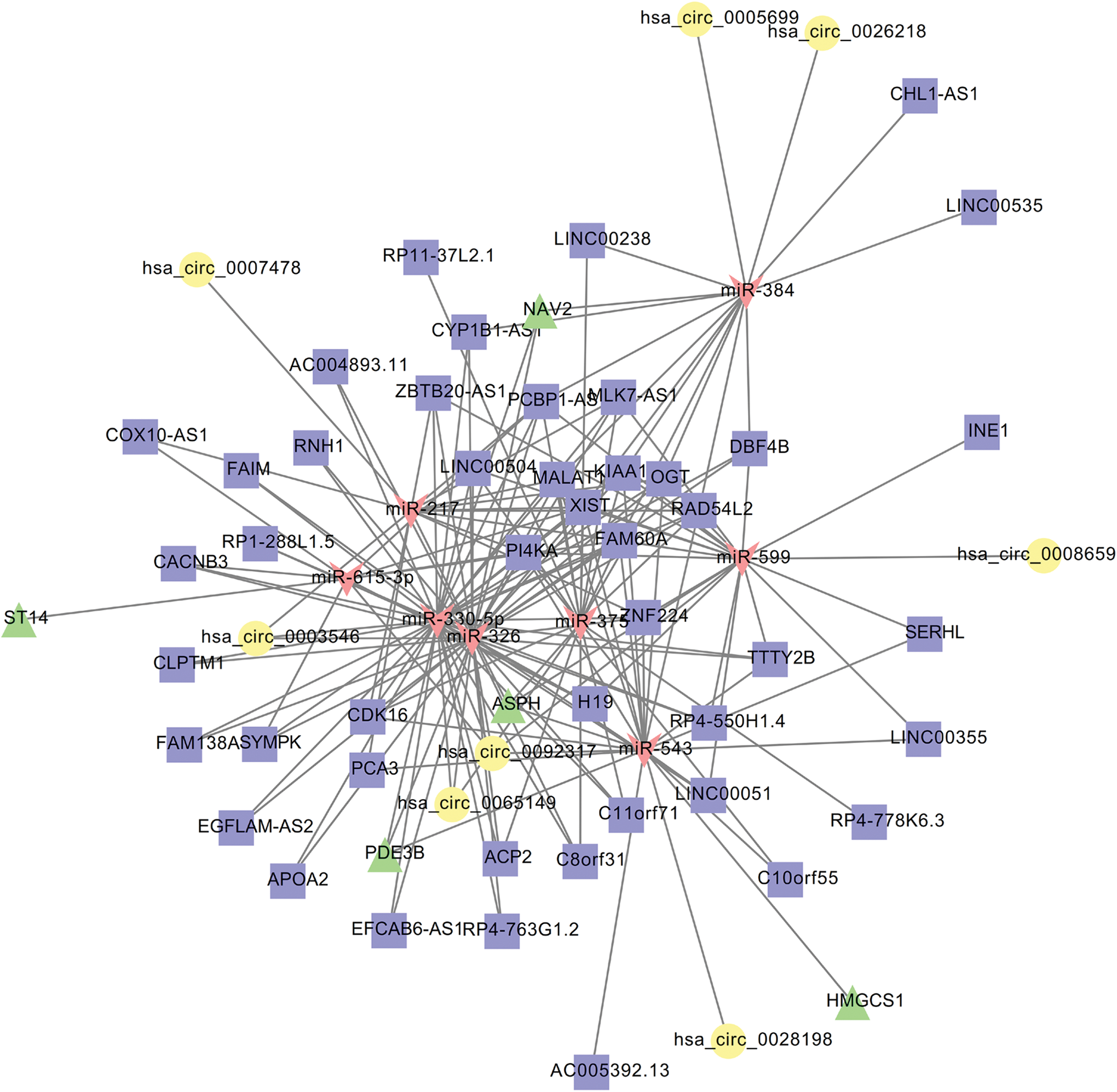
Integrated competing endogenous RNA (ceRNA) interaction network of circRNA/lncRNA-miRNA-mRNA based on the common miRNAs. Square nodes represent lncRNAs; circular nodes represent circRNAs; regular triangle nodes represent mRNAs; and inverted triangle nodes represent miRNAs. Color images are available online.
Discussion
Although eight common DEGs were identified in two microarray datasets, two DEGs (ASPH and PDE3B) may be particularly crucial based on functional enrichment and ceRNA network analyses. ASPH, through interaction with CACNA2D4, may be associated with atherosclerosis by regulating the cellular response to calcium ion; and PDE3B may exert roles in negative regulation of angiogenesis through cross talk with ELMO1. Furthermore, the expression of ASPH and PDE3B may be regulated by hsa_circ_0028198/hsa_circ_0092317/XIST-miR-543; PDE3B expression may also be modulated by hsa_circ_0092317/hsa_circ_0003546/H19/XIST-miR-326.
Phosphodiesterase (PDE) is an enzyme that is responsible for hydrolysis of cyclic adenosine monophosphate (cAMP) (Soderling and Beavo, 2000). cAMP is an important second messenger that activates cAMP-dependent protein kinases (such as PKA) and then contributes to phosphorylation of ABCA1 (ATP-binding cassette transporter A1) to mediate the cholesterol transport process and prevent the formation of foam cells (Liao et al., 2017). Thus, the upregulation of phosphodiesterase genes may be an underlying mechanism to promote the development of atherosclerosis. This hypothesis has been demonstrated previously (Nagaoka et al., 1998; Priksz et al., 2018). For example, Priksz et al. (2018) observed that the level of PDE9A enzyme was significantly elevated in left ventricle samples of animals that received atherogenic rabbit chow supplemented with 1% cholesterol and 1% fat compared with control. Nagaoka et al. (1998) found that the expression and activity of the PDE3A gene were significantly increased in the aortas of atherosclerosis-prone insulin-resistant rats. In addition, Kilanowska and Szkudelski (2019) also proved that inhibition of PDE3B promoted insulin release from isolated pancreatic islets and possibly decreased the risk of diabetes-induced arteriosclerosis (Muhammed et al., 2012; Kilanowska and Szkudelski, 2019). In line with these studies, we also identified that PDE3B was an upregulated gene in macrophages treated with ox-LDL. However, the mechanisms of PDE3B in atherosclerosis remain rarely investigated. In this study, we found that PDE3B could interact with ELMO1. It has been reported that ELMO1/Dock180 proteins form an evolutionarily conserved signaling module to function as a bipartite guanine nucleotide exchange factor for the small GTPase Rac that mediates physical engulfment of apoptotic cells by macrophages (Elliott and Ravichandran, 2010; Schäker et al., 2015). The reduced macrophages due to transformation into foam cells may lead to accumulation of apoptosis, which was also an important pathogenesis for atherosclerosis (Li et al., 2017). Thus, ELMO1 may be downregulated in foam cells, which was also demonstrated in our study. Furthermore, deregulation of PDE3B expression might promote pancreatic islet cell apoptosis (Muhammed et al., 2012). These findings indicate that upregulated PDE3B may be involved in atherosclerosis by downregulation of ELMO1 and then promoting cell apoptosis.
Considering the important roles of PDE in atherosclerosis, PDE inhibitors are hypothesized to be potential drugs for alleviation of atherosclerosis. This hypothesis has been confirmed in current studies by several scholars: clinical trials showed that use of the PDE III selective inhibitor, cilostazol (100–200 mg/day), significantly reduced the mean intima-media thickening of both common carotid arteries and internal carotid arteries, total plaque area (TPA) in patients with a TPA less than 0.5 cm2, and the noncalcified plaque component (Katakami et al., 2010; Kim and Cho, 2013; Lee et al., 2019). In vivo experiments showed that cilostazol decreased triglycerides, increased HDL cholesterol levels, and inhibited foam cell formation (Tani et al., 2000; Okutsu et al., 2009; Park and Heo, 2018). However, attention was also paid to drug complications (myocardial infarction, stent thrombosis, bleeding, rash, and gastrointestinal side effects) and long-term use-induced drug resistance (Sakurai et al., 2013). Therefore, it is necessary to further develop therapeutic approaches that targeted suppression of PDE3B. miRNAs are recently extensively studied ncRNAs that inhibit the expression of target genes by binding to their 3′ UTR (Hausser and Zavolan, 2014). Due to endogenous characteristics, miRNAs have potential for development of novel drugs without major complications. In this study, we also predicted the upstream miRNAs for PDE3B, such as miR-543 and miR-326. Although there was no direct evidence to show the roles of miR-543 and miR-326 in atherosclerosis, they may be speculated to be upregulated according to reports on diabetes (Santovito et al., 2014; Azhir et al., 2018) and inflammatory diseases (Navarro-Quiroz et al., 2017; Wu et al., 2018), which are risk factors for atherosclerosis (Pleskovič et al., 2017). Furthermore, lncRNAs/circRNAs could function as ceRNAs to influence the inhibition effects of miRNAs on target genes (Qu et al., 2015). Hereby, the investigation of lncRNAs/circNAs may also provide targets for development of novel drugs. In this study, we predicted that hsa_circ_0003546/hsa_circ_0092317 and H19/XIST could interact with miR-326, while hsa_circ_0028198/hsa_circ_0092317 and XIST could interact with miR-543. There were studies to report that lncRNA H19 promoted atherosclerosis by regulating MAPK and NF-κB inflammatory signaling pathways (Pan, 2017); knockdown of XIST and H19 may suppress development of coronary atherosclerotic heart disease by suppressing cell apoptosis induced by ox-LDL (Xu et al., 2018; Zhang et al., 2018b). The roles of these circRNAs have not been explored previously, revealing that they may be novel targets for atherosclerosis. The hsa_circ_0003546 parental gene is DENND5A, and the study by Zhang et al. (2018a) showed that ocu-cirR-novel-10230 (corresponding to DENND5A) was significantly upregulated in high-fat diet-induced atherosclerosis rabbits (logFC = 2.59, p = 0.0347), indirectly proving that hsa_circ_0003546 was highly expressed in foam cells, which was in line with our study. No studies were performed to validate the association of hsa_circ_0028198/hsa_circ_0092317 with atherosclerosis and thus they may be novel targets.
ASPH encodes aspartate beta-hydroxylase, which is a type II transmembrane protein and a kind of α-ketoglutarate-dependent dioxygenase. Recently, most of the studies focused on the cancer-promoting effects of ASPH (Sturla et al., 2016; Huang et al., 2018) and only a small portion of studies indicated that recombinant ASPH reduced the viability of natural killer (NK) cells and influenced its cytotoxicity (Huyan et al., 2014). Decreased NK cell activity was reported to be associated with atherosclerosis in elderly humans (Bruunsgaard et al., 2002). Thus, ASPH may be involved in atherosclerosis by decreasing the numbers of NK cells. However, the roles of ASPH remain rarely investigated. In our study, we predicted that ASPH may interact with CACNA2D4 to participate in foam cell formation. CACNA2D4 is a part of the CACN gene family responsible for encoding voltage-dependent calcium channels; its deletion was implicated in bipolar disorder and schizophrenia (Van Den Bossche et al., 2012). The youth with major bipolar disorder are predisposed to develop accelerated atherosclerosis and early cardiovascular diseases (Goldstein et al., 2016). In line with these studies, we also found that CACNA2D4 was downregulated in foam cells. Therefore, targeted inhibition of ASPH may be an underlying approach for treatment of atherosclerosis. Furthermore, we also demonstrated that hsa_circ_0028198/hsa_circ_0092317/XIST-miR-543 may be involved in regulating ASPH through the ceRNA mechanism, indicating that hsa_circ_0028198/hsa_circ_0092317/XIST-miR-543 may be also potential targets for inhibition of ASPH and alleviation of atherosclerosis, as described above.
However, there were some limitations in this study. First, although there is evidence to reveal the associations of H19/XIST/hsa_circ_0003546/miR-326/PDE3B with atherosclerosis, no experiments (dual-luciferase reporter assay, overexpression, or knockout of genes) were performed to validate the ceRNA relationships among them. Second, although our study preliminarily screened ASPH, which may be involved in atherosclerosis, further clinical in vitro and in vivo experiments are necessary to confirm its expression and function mechanisms.
Availability of Data and Materials
The sequencing datasets, GSE107522, GSE54666, and GSE54039, were downloaded from the GEO database in NCBI.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.