
Abstract
Osteosarcoma (OS), a highly aggressive bone tumor, mainly occurs in young patients and always presents abnormalities in molecular biology, such as microRNAs (miRNAs). However, the characteristic and underlying mechanism of miR-671-5p in OS are still unclear. In this study, we certify that miR-671-5p is remarkably downregulated in OS tissues and cells. Overexpressed miR-671-5p can suppress OS cell proliferation in vivo and in vitro, by the way of arresting cell-cycle progression. The overexpression of cyclin D1 (CCND1) and CDC34 promotes cell proliferation and cell-cycle promotion, whose functions are contrary to miR-671-5p. miR-671-5p directly binds to CCND1 and CDC34, which are thought as the key factors in regulating cell cycle. Taken together, our results suggest that by targeting CCND1 and CDC34, miR-671-5p plays a tumor suppressor in OS to inhibit the development of OS, implicating it as a novel target for therapeutic intervention in OS.
Introduction
As one of the most malignant bone tumors, osteosarcoma (OS), with about 66% cases of bone cancers in the world, has a high mortality rate in children and adolescents (Wang et al., 2019a). Cell proliferation is considered a characteristic of cancer development and associates with poor prognosis of patients due to high cancer cellular proliferation rate (He et al., 2017). So far, radical surgical resection combined with neoadjuvant chemotherapy is still the major treatment for OS (Kager et al., 2017; Wang et al., 2019b). Great efforts have been made to improve OS treatment, but the efficacy seems to be very limited. Thus, some patients, diagnosed at the advanced stage, often have poor prognosis, and the growth of the OS is thought as the main reason. Therefore, searching novel mechanisms underlying OS proliferation is extremely urgent, which will make contributions to seeking for new therapeutic targets for OS treatment.
MicroRNAs (miRNAs), a family of ncRNAs with the length of 22–25 nucleotides, could cause degradation or translation repression of target mRNAs by binding to the three prime untranslated regions (3′-UTR) of these genes (Tutar, 2014). Numerous studies showed that miRNAs function in various biological processes, such as cell proliferation (Cantini et al., 2019; Luo et al., 2019). Aberrantly expressed miRNA has been validated to cause dysregulated gene expression in multifarious diseases, especially in cancer (Li et al., 2018; Slattery et al., 2018; Wilk and Braun, 2018). The roles of miRNAs have been widely explored. miRNA-1269a plays a role in promoting progression of OS by inhibiting TGF-β1 expression (Yu et al., 2019), while miRNA-185 inhibits malignant phenotype of MG-63 cells by targeting vesicle-associated membrane protein 2 (Li et al., 2019). By targeting CKAP4, miR-671-3p promotes proliferation of glioma cells (Lu et al., 2018). However, no studies about the regulatory mechanisms of miR-671-5p in OS have been investigated so far.
In this study, we demonstrate that miR-671-5p is downregulated in OS cells and tissues and indicated a poor prognostic features of OS patients, and miR-671-5p could suppress the growth of OS cells in vitro and in vivo. Furthermore, we confirm that miR-671-5p inhibits OS proliferation by directly targeting 3′-UTR of cyclin D1 (CCND1) and CDC34. In addition, we show that miR-671-5p plays a critical role in regulating OS proliferation and might be a potential target of treatment for OS.
Materials and Methods
Identification of the differentially expressed miRNAs in OS
The miRNA data of OS, GSE28425, were downloaded from the Gene Expression Omnibus (GEO). GSE28425 (based on GPL8227 platform) enrolled 19 OS cell lines and 4 human bone tissues. Subsequently, the “edgeR” package was used to identify the differentially expressed miRNAs by fold change (FC) and false discovery rate (FDR) (Robinson et al., 2010). miRNAs with FC ≥2 and FDR <0.05 were considered significantly different.
Cell culture and clinical specimens
The OS cell lines, including U2OS, HOS, Saos-2, MNNG/HOS CI #5, and MG-63, and the normal cell line (hFOB1.19) were cultivated in DMEM, MEM, or RPMI-1640 medium supplemented by 10% fetal bovine serum, 1% streptomycin, and penicillin (100 U/mL; all from Gibco) in 5% CO2 at 37°C. We collected 20 pairs of OS tissues and matched paracancerous tissues, all of which were from the first affiliated hospital of Zhengzhou University, between March 2015 and August 2018. The experiments were approved by the Ethics Committee of Zhengzhou University.
Cell transfection
The miR-671-5p mimics, negative controls, and the vector of overexpressed CCND1 and CDC34 were synthesized and purchased from GenePharma (Shanghai, China). After these plasmids were transfected into MG-63 and U2OS cells through Lipofectamine® 2000 (Invitrogen–Thermo Fisher Scientific) for 48 h, cells were collected for other analysis.
Cell proliferation assay
The viability of cells was tested by Cell Counting Kit-8 (CCK8) assay kit (Qihai, Shanghai, China). After transfection with plasmid, the absorbance at 450 nm was measured with SUNRISE Microplate Reader (Tecan Group Ltd., Mannedorf, Switzerland) at 0, 24, 48, 72, and 96 h. For colony formation assay, cells were cultured in six-well plates for 14 days. After cells were fixed and stained, the counts of colonies were then evaluated.
Flow cytometry analysis of the cell cycle and apoptosis
MG-63 and U2OS cells transfected with miR-671-5p mimics, negative controls or CCND1 and CDC34 plasmid, were stained with cell cycle assay kit (PI; Qihai) and Annexin V-fluorescein isothiocyanate (FITC) and 7-AAD using an FITC Annexin V Apoptosis Detection Kit. Then, cells were analyzed by flow cytometry (FACScan; BD Biosciences) according to the manufacturer's recommendations.
Quantitative RT-PCR and western blotting
RNA was extracted from tissues and cells using TRIzol Kit (Takara), and using SYBR® Fast qPCR Mix (Takara), quantitative RT-PCR (qRT-PCR) was performed through the Real-Time PCR System (Bio-Rad). For miRNA-671-5p, U6 was used as normalizer. And for CDC34 and CCND1 mRNA, β-actin was used as normalizer. The specific primers for CCND1 and CDC34 were used; the sequences were shown as follows: miRNA-671-5p: the forward primer, 5′GCCCGCAGGAAGCCCUGGAGGGGC3′; the reverse primer, 5′GTGCAGGGTCCGAGGT3′. CCND1: the forward primer, 5′CTAAGATGAAGGAGACCATCCC3′; the reverse primer, 5′AAGGTCTGCGCGTGTTTGCGGAT3′. CDC34: the forward primer, 5′ACAGAAACAGGTGCGCTTACC3′; the reverse primer, 5′CAGCCGGTCACGTTCTTCTTT3′.
For western blotting, cells were lysed using RIPA buffer (HAT Biotechnology, Xi'an, China). Then, the lysates were separated by 12% SDS-PAGE, and transferred to PVDF membranes (Millipore). The primary antibodies against CDC34 (1:1000; Proteintech Biotechnology, China), CCND1 (1:1000; Proteintech Biotechnology), Bcl-2 (1:1000; Proteintech Biotechnology), P21 (1:1000; Proteintech Biotechnology), BAX (1:1000; Proteintech Biotechnology), and β-actin (1:5000; Proteintech Biotechnology) were purchased, and β-actin was used as the loading control.
Dual-luciferase reporter assays
The MG-63 and U2OS cells were cultivated overnight in 24-well plates and then transfected, 0.5 mg of the overexpression of the miR-671-5p vector and 0.5 mg of the wild and mutant CCND1 or CDC34 luciferase reporter plasmid (Genechem, Shanghai, China), per well with Lipofectamine 2000. After transfection for 48 h, luciferase activity analysis was performed by a Dual-Luciferase Reporter Assay System (Promega).
In vivo experiments
Four-week-old male BALB/c nude mice (Centre of Laboratory Animals, the Medical College of Zhengzhou University, Zhengzhou, China) were purchased. To develop a nude mouse xenograft model, MNNG/HOS CI #5 (1 × 107) cells that were transfected with miR-671-5p or miR-control vectors were subcutaneously injected into nude mice. For each group, there are six nude mice involved. After 3 weeks, the mice were sacrificed and the xenograft tumor tissue was saved for further investigation. Animal experiments were approved by the Institutional Animal Care and Use Committee of Zhengzhou University.
Immunohistochemical staining
The xenograft tumor tissue was soaked in 4% paraformaldehyde and embedded in paraffin. After the tissue was sliced, the sections were incubated in the primary antibodies overnight. The primary antibodies against Ki67 (1:200) and proliferating cell nuclear antigen (PCNA) (1:200) were purchased from Servicebio. Following the reaction by diaminobenzidine and counterstain, the results were then evaluated under a microscope.
Statistical analysis
We set up three replicates for each samples and all experiments were repeated three times. The data involved in our experiment are shown in forms of mean ± SD. We plotted the Kaplan–Meier curve to present the survival of OS patients. The expression correlation between CCND1 or CDC34 and miR-671-5p was analyzed by Pearson correlation coefficient. p < 0.05 was considered statistically significant.
Results
Identification of differentially expressed miRNAs
Nineteen OS cell line samples and four human bone tissue samples were enrolled in our study. After differential expression analysis, a total of 209 miRNAs, consisting of 104 upregulated and 105 downregulated miRNAs (Supplementary Table S1), were identified as differentially expressed miRNAs in OS cell lines versus normal bone tissues and we displayed the results in a volcano plot (Fig. 1). Moreover, in the list, we found the expression level of miR-671-5p was obviously low in OS cell lines, while high in human normal bone tissues.

Volcano plot of differentially expressed miRNAs. The red dots stand for upregulated miRNAs, and the blue dots stand for downregulated miRNAs. FDR, false discovery rate; miRNAs, microRNAs. Color images are available online.
miR-671-5p is downregulated in OS and correlates with patients' survival
To further detect the expression of miR-671-5p in OS, qRT-PCR was conducted to test its expression in 20 pairs of OS tissues and adjacent nontumor tissues, both of which were selected randomly. The specific clinical characteristics of these patients such as sex and stage are shown in Supplementary Table S2. Results showed that miR-671-5p was obviously downregulated in OS tissues versus nontumor tissues (Fig. 2A). Also, miR-671-5p was obviously decreased in four common OS cell lines compared with normal bone cell line hFOB 1.19, respectively (Fig. 2B). Next, all OS patients were divided into miR-671-5p high (n = 10) and low (n = 10) group according to median value. It revealed OS patients who have a low miR-671-5p level showed a distinctly short overall survival by Kaplan-Meier analysis (Fig. 2C). These results indicate that miR-671-5p could serve as a prognostic biomarker for OS.
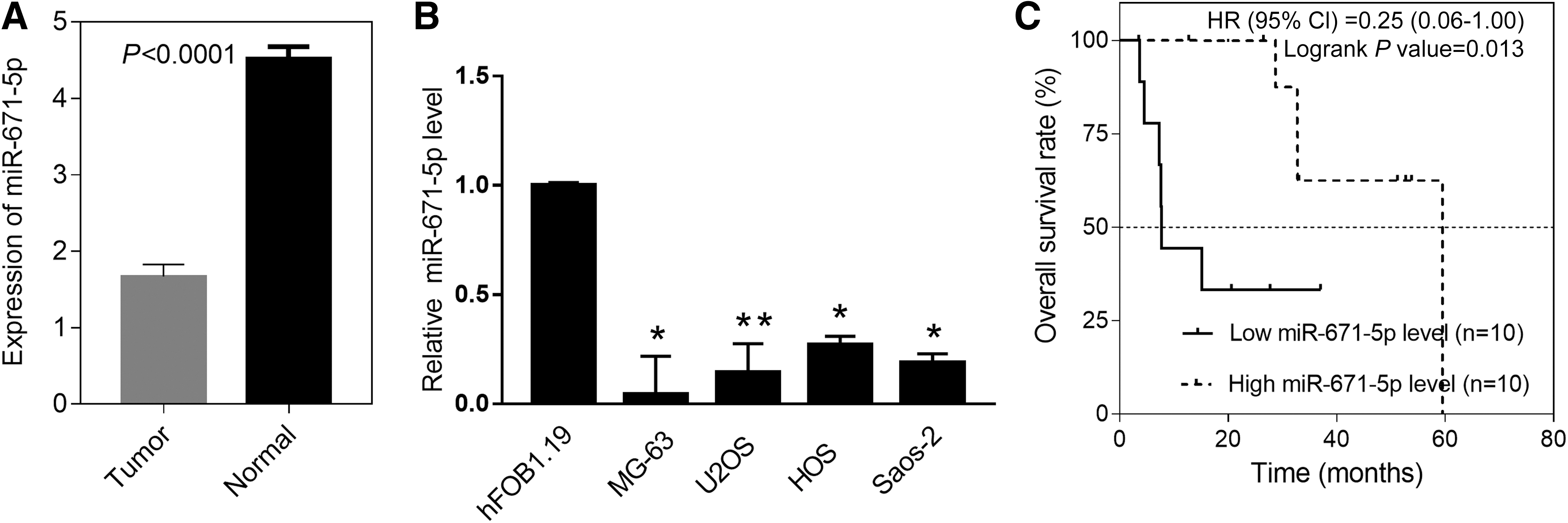
The expression of miR-671-5p in OS and associates with OS survival. The expression level of miR-671-5p in OS and matched nontumor tissues
miR-671-5p inhibits OS cell growth
To explore the biological function of miR-671-5p in OS better, we choose MG-63 and U2OS cells with the lower expression of miR-671-5p for further cell experiment. First, miR-671-5p mimics were transfected both in MG-63 and U2OS cells. Then, we verified miR-671-5p was significantly upregulated in MG-63 and U2OS cells by qRT-PCR (Fig. 3A). In addition, overexpression of miR-671-5p suppressed cell proliferation by CCK8 assays (Fig. 3B), and decreased cell colonies according to the colony formation assay (Fig. 3C). As determined by flow cytometry, cell cycle transition was arrested from G1 to S phase (Fig. 3D) through overexpression of miR-671-5p, which could induce apoptosis (Fig. 3E). Thus, these data demonstrated that it was regulated by miR-671-5p, including OS cell proliferation, cell cycle progression, and apoptosis in vitro. To sum up, our findings show miR-671-5p inhibits the growth of OS cells in vitro.

miR-671-5p inhibits cell proliferation and progression of cell cycle, and promotes apoptosis of OS cells in vitro.
CCND1 and CDC34 are downstream targets of miR-671-5p in OS
To dissect the regulatory mechanism of miR-671-5p in regulating of OS proliferation, miRanda and TargetScan are applied to seek for target genes of miR-671-5p. Intriguingly, the 3′-UTR of CCND1 and CDC34 was the target of miR-671-5p (Fig. 4A). Luciferase reporter assay was performed to investigate whether miR-671-5p could directly target the 3′-UTR of CCND1 and CDC34. The results showed cells transfected with the wild CCND1 or CDC34 3′-UTR showed a lower luciferase activity, however, a higher luciferase activity in cells transfected with the mutant CCND1 or CDC34 3′-UTR (Fig. 4B, C). Overexpressed miR-671-5p could decrease the mRNA and protein level of CCND1 and CDC34 in MG-63 and U2OS cells, respectively (Fig. 4D, E). Our results show miR-671-5p reduces the expression of CCND1 and CDC34 by directly targeting their 3′-UTR.
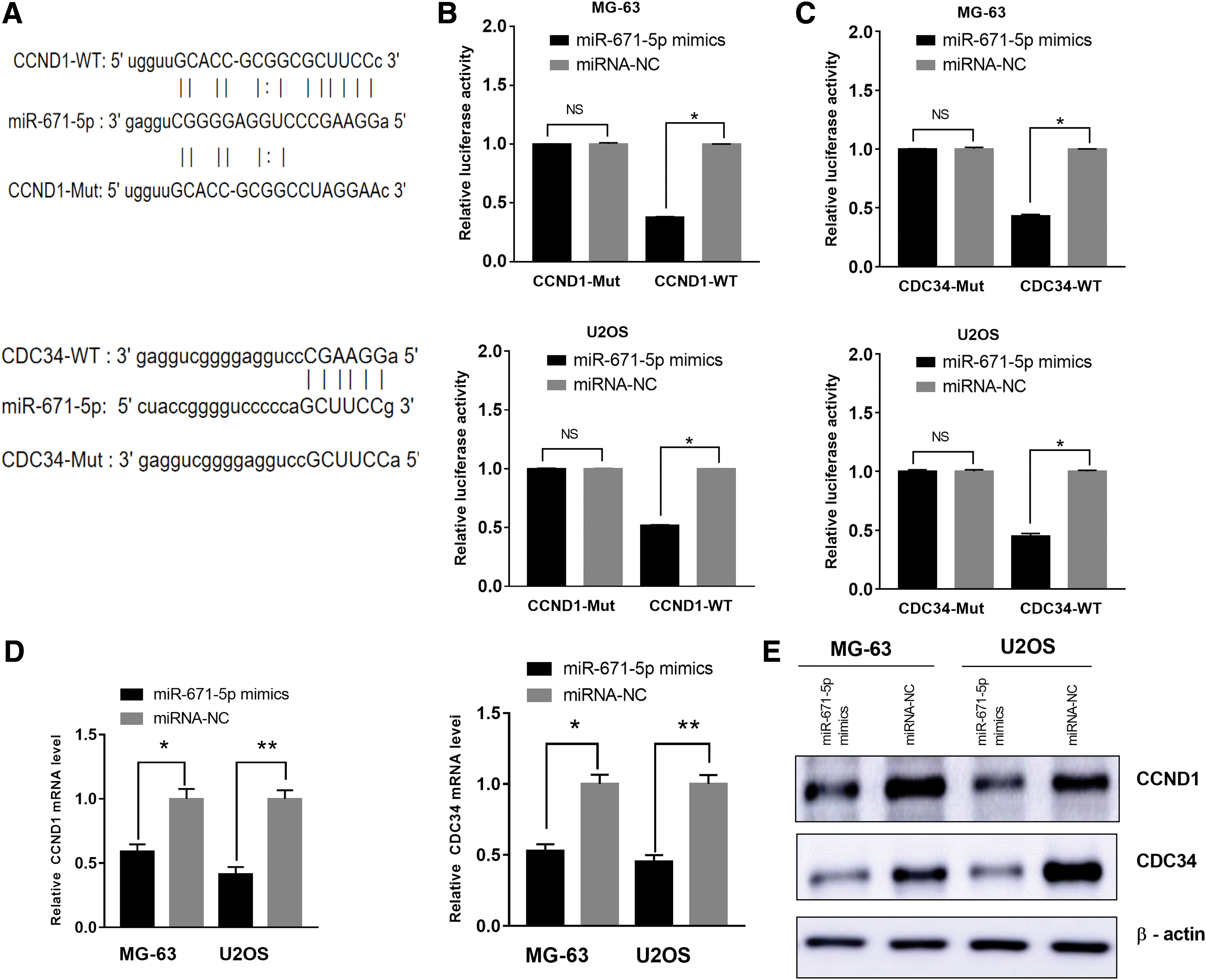
CCND1 and CDC34 are direct target genes of miR-671-5p in OS cells.
miR-671-5p negatively correlated with CCND1 and CDC34 in OS tissues and suppressed the growth of OS in vivo
To figure out the association between expression of miR-671-5p and CCND1 and CDC34, expression of CCND1 and CDC34 in different miR-671-5p level groups of human OS tissues was tested. Results showed the mRNA level of CCND1 (R = 0.372, p = 0.004) and CDC34 (R = 0.613, p < 0.001) has a negative correlation with miR-671-5p expression in all OS tissues (Fig. 5A). Consistently, immunohistochemistry (IHC) results showed that PCNA and Ki67 protein expression in miR-671-5p high-expressing tumors (n = 10) were obviously lower than those in miR-671-5p low-expressing tumors (n = 10) (Fig. 5B).

The relationship between miR-671-5p and CCND1 or CDC34 expression of OS tissues and the role of miR-671-5p in the growth of OS in vivo.
For investigating the role of miR-671-5p in tumorigenicity in vivo, we carried out the subcutaneous tumor formation experiment in nude mice. The picture of tumor size showed that the overexpression of miR-671-5p obviously suppressed the proliferation of MNNG/HOS CI #5 cells in nude mice (Fig. 5C). Also, there is a lower level of PCNA and Ki67 in tumors with high miR-671-5p level, whereas a higher level of PCNA and Ki67 protein expression in tumors with low miR-671-5p level as shown in IHC (Fig. 5D). From the above, our data indicate the level of CCND1 or CDC34 is inversely connected with miR-671-5p in OS tissues and miR-671-5p can inhibit the growth of OS in vivo.
Restoration of CCND1 and CDC34 reverses the biological effects of miR-671-5p
For a better understanding about how CCND1 and CDC34 influence the biological functions of miR-671-5p, CCND1 or CDC34 was restored in MG-63 and U2OS cells, respectively (Fig. 6A). The result was the suppressive functions of miR-671-5p was reversed by way of CCND1 or CDC34 restoration on cell proliferation (Fig. 6B), colony formation (Fig. 6C), cell cycle progression (Fig. 6D), and apoptosis (Fig. 6E). What is more, the overexpressed CCND1 or CDC34 can reverse the impact of miR-671-5p on apoptosis-related protein (Fig. 6F). These results show that both CCND1 and CDC34 can serve as available functional mediators of miR-671-5p.
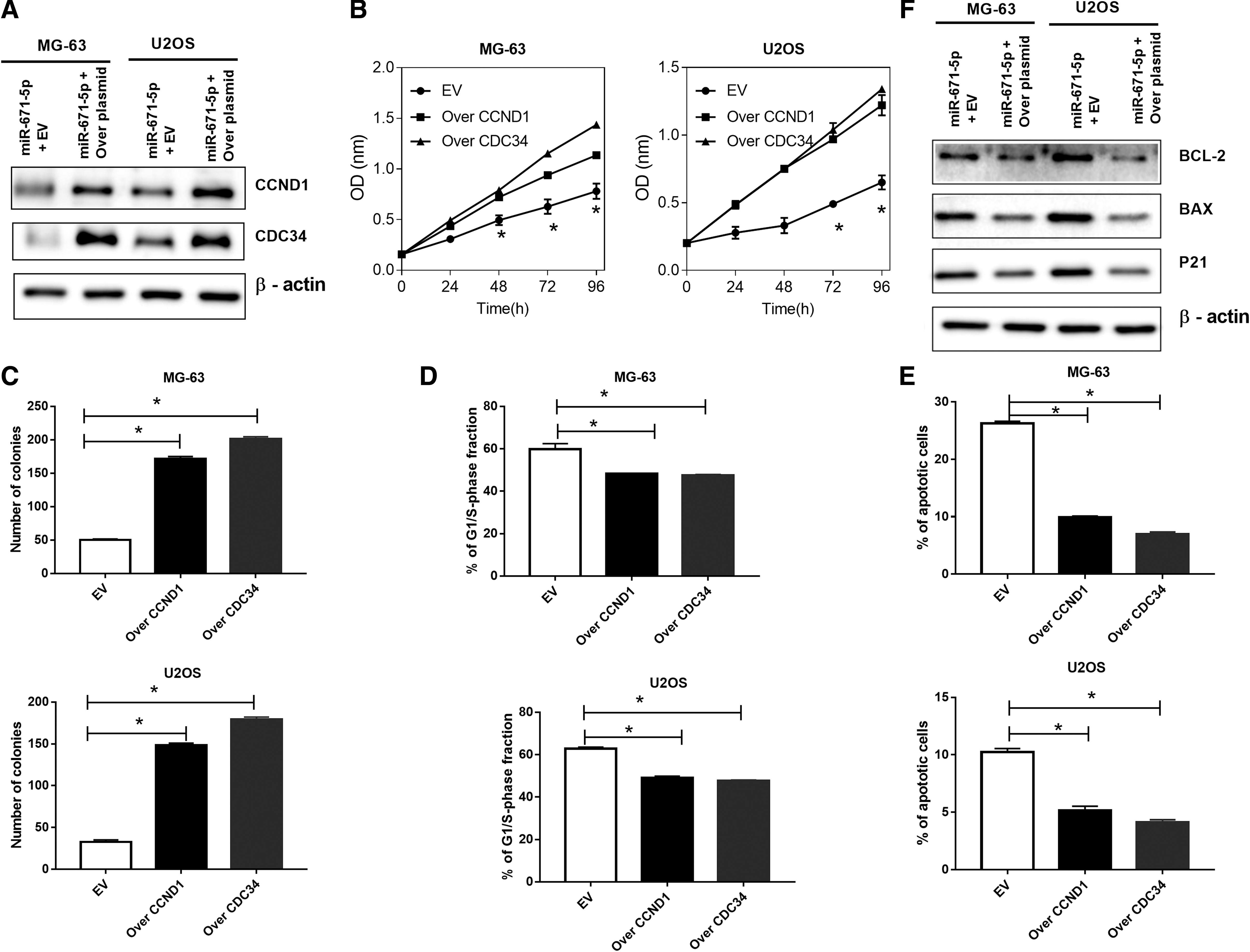
CCND1 or CDC34 partially restores miR-671-5p-mediated biological effects in OS cells.
Discussion
Increasing studies have confirmed that miRNAs involved in OS, such as progression and so on (Zhang et al., 2019). Recently, miRNAs are recognized as the key regulators and predictors in OS (Nakka et al., 2017; Ji et al., 2018). Our study has shown that miR-671-5p was in the low expression in OS tissues and cell lines, consistent with the GEO data. Furthermore, OS patients with low miR-671-5p level are more likely to have poor prognosis. All the above indicate miR-671-5p participated in OS progression and could serve as a potential biomarker for OS treatment.
Researchers have certified that miR-671-5p could inhibit EMT by targeting FoxM1 in breast cancer (Tan et al., 2016), while promoting prostate cancer cell proliferation (Yu et al., 2018). In our research, it was certified miR-671-5p not only inhibited cell proliferation, progression of cell cycle, and colony formation but also promoted apoptosis in vitro and in vivo. Furthermore, we first put forward the evidence that CCND1 and CDC34 were the target genes for miR-671-5p. First, miR-671-5p played a negative regulation role in the expression of CCND1 and CDC34 in OS cells, which was validated in OS cells by luciferase activity analysis and in OS tissues through qRT-PCR, respectively. Furthermore, biological functions of miR-671-5p could be reversed by regulating the expression of CCND1 or CDC34 in OS cells. CCND1 is a key cell cycle regulatory protein and the gate-keeping protein that charges regulating the transition from G1 phase to S phase of the cell cycle. Studies have demonstrated oncogenic capacity of CCND1 in various cancers, indicating that CCND1 may contribute toward therapy (John et al., 2017). For example, Chen et al. (2019) found miRNA-584 inhibits proliferation and invasion of pancreatic cancer cells by targeting CCND1. Belonging to the family of ubiquitin-conjugating enzymes, CDC34, a regulator of the G1 to S phase transition (de Bie and Ciechanover, 2011), can catalyze the addition of ubiquitin to other protein substrates (Craig and Tyers, 1999) and plays a significant role in mediating ubiquitination and degradation of p21 and p27 (Liu et al., 2018a, 2018b). CDC34, highly expressed in human cancers, can induce cell cycle arrest in G1 phase in head and neck squamous cell carcinoma (Han et al., 2018). In this study, we also found CCND1 and CDC34 were highly expressed in OS cell lines. All the above will help to strengthen our understanding of the role of miRNA in cancer.
It is revealed in previous research the target genes of miR-671 are mainly enriched in the regulation of cell proliferation (Tan et al., 2016; Yu et al., 2018). Dysregulation of cell proliferation plays a crucial function in malignant transformation of normal cells (Brandmaier et al., 2017). However, the cell proliferation process is mainly relied on genes involved in the cell cycle, the dysregulation of which could lead to abnormal cell cycle and cell proliferation (Dalton, 2015; Hafner et al., 2019). The dysregulation of cell cycle kinases could give rise to an abnormal growth of OS cells (Cheng et al., 2016). And cell cycle-associated genes can be regulated by all kinds of genes, including miRNAs (Hydbring et al., 2017; Otto et al., 2017). Our research found the effects of miR-671-5p on OS growth could be abolished by alteration of CCND1 and CDC34. In this study, cell cycle plays an important part in biological functions of miR-671-5p in OS. Taken the results above, it can be confirmed miR-671-5p inhibits OS cell growth by the way of blocking cell cycle.
In conclusion, our results suggest there exists the downregulation of miR-671-5p in OS tissues and cell lines, and the low level of miR-671-5p indicates a poor prognosis. Besides, miR-671-5p suppresses OS cell proliferation. Furthermore, CCND1- and CDC34-mediated cell cycle pathway has a direct participation in the oncogenic function of miR-671-5p in OS. The results above define the vital role of miR-671-5p in maintaining cellular proliferation in OS, and miR-671-5p could serve as a possible molecular target for the therapy of OS.
Conclusion
In a word, our study concluded that miR-671-5p could inhibit the proliferation of OS in vivo and in vitro by directly targeting CCND1 and CDC34, which are pivotal in cell cycle procession. Thus, miR-671-5p/CCND1/CDC34 axis might be the potential target for OS treatment.
Footnotes
Acknowledgment
This work was supported by grants from the National Natural Science Foundation of China Nos. 81702663.
Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.