
Abstract
Our previous genome-wide association study has identified a suggestive association at rs11264799 within FCRL3 (Fc receptor-like 3) locus on 1q23.1 for IgA nephropathy (IgAN) in a Chinese Han population. This study aims to investigate the association of FCRL3 variants with the susceptibility, clinicopathological phenotypes and prognosis of IgAN. Eleven FCRL3 single-nucleotide polymorphisms (SNPs) were selected and analyzed in this two-stage case/control study with a total of 1750 IgAN cases and 2500 healthy controls in a Chinese Han population. Unconditional logistic regression models were used to estimate odds ratios and 95% confidence intervals (CIs) as implemented in the PLINK software. Luciferase assays were applied to detect the allelic effect of rs11264794 on gene expression regulation. We found that four SNPs (rs11264794, rs7865684, rs11264799, and rs6691569) were significantly associated with IgAN susceptibility after Bonferroni correction in the combined samples. Genotype/phenotype association analysis observed that two SNPs (rs11264794 and rs11264793) were associated with less disease severity. After adjusting for confounders, rs11264794 was independently correlated with renal outcome in IgAN patients (hazard ratio = 0.64, 95% CI = 0.43–0.97, p = 0.033). In addition, the protective allele A of rs11264794 was significantly associated with higher FCRL3 gene expression. Furthermore, luciferase reporter gene assays demonstrated that the minor allele of rs11264794 obviously reduced the specific binding between miR-183-5p.1 and FCRL3 3′-untranslated region. Our results indicate that FCRL3 gene polymorphisms are associated with the development and progression of IgAN, and the rs11264794-A allele showed a protective role for IgAN.
Introduction
IgA nephropathy (IgAN) is the most common glomerular disease, accounting for 45.3–54.3% of all primary glomerulonephritis and representing a major cause of end-stage renal disease (ESRD) in China (Liu, 2013). IgAN is considered an autoimmune disease with a multihit pathogenetic process regulated by both genetic and environmental factors (Knoppova et al., 2016; Li and Yu, 2018). However, the exact etiology of IgAN remains unknown, and no specific treatment has been established.
Genome-wide association studies (GWAS) and candidate gene-based studies have identified multiple genes that confer susceptibility to IgAN (Gharavi et al., 2011; Yu et al., 2011a, 2011b; Kiryluk et al., 2014; Li et al., 2015). It has been suggested that inherited defects of IgAN are associated with innate, adaptive, and mucosal immunity, which are all involved in the pathogenesis of IgAN (Rauen and Floege, 2017). Nonetheless, the identified loci to date can only explain a small fraction of the genetic risk of IgAN, suggesting that there are additional genetic factors remained to be uncovered.
We previously performed a GWAS for IgAN in a Chinese population and identified a suggestive signal within the FCRL3 (FC receptor-like 3) locus at rs11264799 on chromosome 1q23.1 (Li et al., 2015). FCRL3 is one of the members of several Fc receptor-like glycoproteins, and shares a high structural homology with classical Fc receptors (Davis, 2007). The protein encoded by FCRL3 contains immunoreceptor tyrosine activation motif (ITAM) and immunoreceptor tyrosine inhibitory motif (ITIM) in its cytoplasmic domain, which are thought to play a bidirectional role in regulating the immune system (Davis, 2007). FCRL3 is primarily expressed in lymphoid organs, particularly on the surface of B cells (Li et al., 2013), as well as T cells (Swainson et al., 2010) and natural killer cells (Parkes et al., 2018).
FCRL3 was demonstrated to augment the inhibitory signal of the B cell receptor (BCR) and affect the generation and differentiation of B cells, which can lead to a change in antibody production (Li et al., 2013). Moreover, Swainson et al. (2010) found that FCRL3 gene expression was associated with regulatory T cell (Treg) dysfunction, which may contribute to the loss of self tolerance. In addition, accumulating evidences have shown that genetic polymorphisms of FCRL3 are associated with several immune-related diseases, including rheumatoid arthritis (Kochi et al., 2005), multiple sclerosis (Yuan et al., 2016), and Graves' disease (Fang et al., 2016). Based on these findings, the FCRL3 gene is likely to modulate immune cell functions and contribute to the pathogenesis of autoimmune diseases.
It is speculated that FCRL3 may play an important part in the development of IgAN. However, until now, there has been no systematic investigation about the role of FCRL3 in IgAN. The purpose of this study was to determine the relationships between single-nucleotide polymorphisms (SNPs) within the FCRL3 gene and the susceptibility, clinical phenotypes and disease progression of IgAN in a Chinese Han population.
Materials and Methods
Subjects
In this study, we performed a two-stage case/control study to evaluate the association between FCRL3 candidate SNPs and the susceptibility to IgAN. A total of 1750 cases and 2500 healthy controls were recruited from The First Affiliated Hospital of Sun Yat-sen University, consisting of 1000 cases and 1000 controls for the discovery stage (stage 1) and 750 cases and 1500 controls for the validation stage (stage 2). All the subjects were of self-reported Chinese Han ancestry and were unrelated.
All the cases were biopsy proven, with no evidence of secondary IgAN or other autoimmune diseases. The age- and gender-matched healthy controls were recruited from the same hospital and verified with normal urinalysis and no history of renal diseases. The detailed clinical information of cases at the time of diagnosis was collected from patients' files and questionnaires. The study was performed in compliance with the tenets of the Declaration of Helsinki and approved by the Research Ethics Committee of The First Affiliated Hospital, Sun Yat-sen University (No. 201037). All participants provided written informed consent.
SNP selection and genotyping
SNP genotyping information was retrieved from the HapMap (release 27, phase I + II+III,
Besides, three potential functional SNPs (rs7522061, rs7528684, and rs945635) were chosen after screening the online web server SNPinfo. In addition, two previously reported SNPs (rs2282284 and rs3761959) were also included due to their significant associations with other autoimmune diseases (Kochi et al., 2005; Lan et al., 2015; Yuan et al., 2016). Among these 12 SNPs, rs11264798 was excluded for further analysis owing to complete linkage disequilibrium (LD) (r 2 = 1) with the potential functional SNP rs7522061. Finally, a total of 11 SNPs (rs11264793, rs6691569, rs11264799, rs10489678, rs12030807, rs11264794, rs7522061, rs7528684, rs945635, rs2282284, and rs3761959) were selected for association analysis.
Genotyping of the 11 investigated SNPs in stage 1 was performed using the Sequenom MassARRAY platform (Sequenom, San Diego, CA). In stage 2, six candidate SNPs were genotyped by LGC Genomics (London, United Kingdom), using the Kompetitive Allele-Specific PCR genotyping platform. Specific primers were designed for each variant (Supplementary Tables S1 and S2).
Bioinformatic analysis of SNP rs11264794
HaploReg version 4.1* and RegulomeDB database** were used to determine the potential functional role for SNP rs11264794. TargetScan Human 7.1*** PolymiRTS 3.0 † and miRBase 18 databases ‡ were used to predict whether any microRNAs (miRNAs) can bind to the region around SNP rs11264794.
Transient transfection and luciferase reporter assays
Reporter assays were used to detect the allelic effect of rs11264794 on FCRL3 gene expression. FCRL3 3′-untranslated region (3′-UTR) with the rs11264794-T or -G allele was amplified by PCR and cloned into the psiCHECK™-2 vector (Promega, Madison, WI) at NotI and XhoI restriction sites (3′-UTR-G-psiCHECK-2 and 3′-UTR-T-psiCHECK-2). The blank plasmid of psiCHECK-2 was used as control. All plasmids were verified by sequencing before use. MiR-183-5p.1 (UAUGGCACUGGUAGAAUUCACU) and miR-4470 (UGGCAAACGUGGAAGCCGAGA) mimics were chemically synthesized by the RiboBio Company (Guangzhou, China).
Human embryonic kidney 293 (ATCC, Manassas, VA) cells were maintained in antibiotic-free Dulbecco's modified Eagle's medium (Gibco) supplemented with 10% fetal bovine serum (Gibco). Cells were seeded into 24-well plates and transfected with 500 ng of each plasmid DNA (psiCHECK-2-control, 3′-UTR-G-psiCHECK-2, and 3′-UTR-T-psiCHECK-2) with or without miR-183-5p.1 or miR-4470 mimics (100 nM) using Lipofectamine 3000 (Life Technologies, Carlsbad, CA).
After 48 h of transfection, the cells were harvested, lysed, and analyzed the firefly and Renilla luciferase activities according to the protocol of dual-luciferase reporter assay system (Promega). The ratio of Renilla to firefly luciferase activities was calculated as the relative luciferase activity for each sample. Each experiment was repeated three times, and each transfection experiment included three biological replicates for each allele.
Measurement of serum IgA1 and galactose-deficient IgA1 levels
To evaluate the correlations between FCRL3 SNPs and serum IgA1 and galactose--deficient IgA1 (Gd-IgA1) levels, we have rerecruited 96 IgAN cases to detect their serum IgA1 and Gd-IgA1 levels using enzyme-linked immunosorbent assay as previously reported (Moldoveanu et al., 2007).
Quantitative real-time PCR and flow cytometry analysis
Peripheral blood mononuclear cells (PBMCs) were isolated by Ficoll/Hypaque density gradients (Sigma Chemical Co., St Louis, MO). Total RNAs from PBMCs of 68 healthy controls and 46 IgAN patients were purified with TRIzol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. Eluted RNAs were reverse transcribed to generate cDNA using the Transcriptor First Strand cDNA Synthesis Kit (Roche, Switzerland). FCRL3 mRNA expression levels were quantified by quantitative real-time PCR with TaqMan® (Hs00364720_m1), using GAPDH (Hs03929097_m1) as endogenous control. All reactions were analyzed in duplicate on an ABI 7900HT Fast Real-Time PCR System (Applied Biosystems). The relative expression levels of FCRL3 were described using the equation 2−ΔCt, where ΔCt (delta Ct) = Ct candidate (FCRL3) − Ct GAPDH.
PBMCs were washed with phosphate-buffered saline (PBS) and then stained with Alexa Fluor® 647 mouse anti-human FCRL3 (BD Pharmingen, San Diego, CA) or Alexa Fluor 647 mouse IgG2b isotype control (BD Pharmingen) for 30 min at 4°C. After staining, the cells were washed twice with PBS and analyzed by flow cytometry (Beckman Coulter, Miami, FL). The results were expressed as the percentage of cells positive for FCRL3.
Statistical analysis
Quantitative data were expressed as mean ± standard deviation for normally distributed variables and median (interquartile range) for skewed distributions. The Hardy/Weinberg equilibrium (HWE) of selected SNPs in control samples was assessed by the chi-square test.
Associations between FCRL3 SNPs and IgAN susceptibility were evaluated with unconditional multivariate logistic regression analysis under three models (additive, dominant, and recessive genetic models) as previously described (Xu et al., 2014). For individual SNP analysis, Bonferroni's correction was carried out to correct the p-values for multiple testing. Haploview version 4.2 was used to obtain the LD patterns (Barrett et al., 2005). Association analysis between FCRL3 haplotypes and IgAN risk was performed using 1000 permutation tests.
Genotype/phenotype analysis was conducted under an additive model. Multiple linear regression analysis was used for quantitative traits, and logistic regression was applied for binary and ordinal variables. In addition, a retrospective cohort study was conducted to evaluate the effect of FCRL3 SNPs on the progression of IgAN. The endpoint was defined as doubling of serum creatinine level or progress to ESRD (estimated glomerular filtration rate [eGFR] <15 mL/min/1.73 m2 or need renal replacement therapy). Kaplan/Meier method and Cox proportional hazard model were used to evaluate the survival of IgAN patients with available clinical follow-up data.
PLINK software (version 1.07) (Purcell et al., 2007) was used to evaluate the genetic association of FCRL3 gene polymorphisms and haplotypes with IgAN. Other analyses were performed with SPSS 19.0 software (SPSS, Inc., Chicago, IL) and GraphPad Prism 5.0 software (GraphPad Software, Inc., San Diego, CA).
Results
Clinical features of the study participants
A total of 971 cases and 970 controls remained in stage 1, and 707 cases and 1459 controls remained in stage 2 for further analysis after quality control, where 59 individuals (29 cases and 30 controls) in stage 1 and 84 individuals (43 cases and 41 controls) in stage 2 were excluded for poor genotyping results. No significant differences in terms of age (p = 0.055) and gender (p = 0.616) were noted among the two groups (Supplementary Table S3). The complete clinical information of IgAN patients from stage 1 is shown in Supplementary Table S4.
Association between FCRL3 polymorphisms and IgAN susceptibility
The detailed information of the 11 candidate SNPs is shown in Supplementary Table S5. All the SNPs were successfully genotyped with call rates >95%, and the genotype counts were consistent with the HWE (p > 0.05) in two stages.
In stage 1, nine SNPs (rs11264794, rs7528684, rs11264799, rs6691569, rs11264793, rs12030807, rs7522061, rs3761959, and rs945635) showed nominal associations (p < 0.05) with IgAN susceptibility (Table 1). Then, we evaluated the LD pattern among the 11 candidate SNPs in stage 1 and found that 4 SNPs (rs7528684, rs7522061, rs3761959, and rs945635) were in high LD (r 2 = 0.99) (Fig. 1). Then, six SNPs (rs11264794, rs7528684, rs11264799, rs6691569, rs11264793, and rs12030807) were selected for validation in stage 2, and four SNPs were revealed to be more strongly associated with IgAN under additive models (rs11264794, odds ratio [OR] = 0.85, 95% confidence interval [CI] = 0.75–0.97, p = 0.015; rs7528684, OR = 0.85, 95% CI = 0.74–0.97, p = 0.014; rs11264799, OR = 0.79, 95% CI = 0.67–0.93, p = 0.005; and rs6691569, OR = 0.81, 95% CI = 0.67–0.97, p = 0.019) (Table 1).
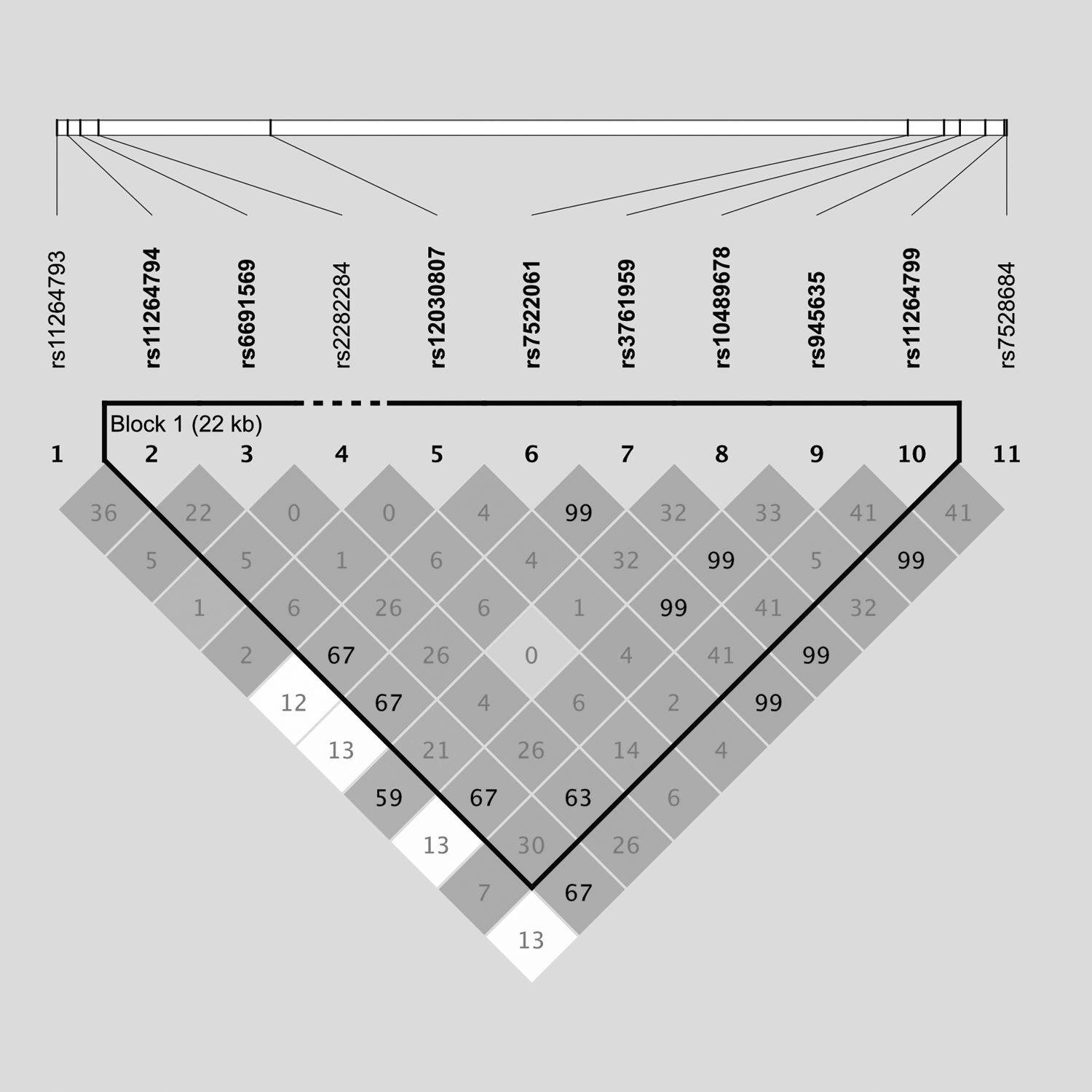
The LD plot for SNPs of FCRL3 measured by r 2. The block structure was estimated using Haploview 4.2, and one LD block has been identified (black solid line). FCRL3, Fc receptor-like 3; LD, linkage disequilibrium; SNP, single-nucleotide polymorphism.
Association Analysis of Fc Receptor-Like 3 Polymorphisms with Susceptibility to IgA Nephropathy
The p-values were obtained by logistic regression analysis with adjustment of age and gender.
CI, confidence interval; IgAN, IgA nephropathy; NA, not available; OR, odds ratio; SNP, single-nucleotide polymorphism.
In the combined samples, three SNPs were more significantly associated with reduced IgAN risk after Bonferroni correction under additive models (rs11264794, OR = 0.83, 95% CI = 0.75–0.90, p = 2.20 × 10−5; rs7528684, OR = 0.85, 95% CI = 0.78–0.93, p = 5.61 × 10−4; and rs6691569, OR = 0.83, 95% CI = 0.73–0.93, p = 0.002), while the rs11264799 was more significantly associated under a recessive model (OR = 0.59, 95% CI = 0.43–0.80, p = 9.71 × 10−4) (Table 1).
Furthermore, the haplotype structures of the four significant SNPs in the combined samples were further investigated (Supplementary Fig. S1). The results of haplotype analysis are shown in Table 2. Notably, the haplotype carrying protective alleles of A of rs11264794, G of rs7528684, T of rs11264799, and A of rs6691569 showed a protective effect on the susceptibility to IgAN (AGTA; OR = 0.83, 95% CI = 0.73–0.94, p = 0.019), while the haplotype carrying risk alleles of these four SNPs showed a significantly increased risk for IgAN (CACG; OR = 1.22, 95% CI = 1.12–1.33, p = 9.00 × 10−4).
Haplotype Analysis of Fc Receptor-Like 3 Polymorphisms with the Risk of IgA Nephropathy
Haplotypes with the frequency <0.01 were ignored in the analysis. The p-value <0.05 shown in bold.
Haplotypes consisted of SNPs rs11264794 (A/C), rs7528684 (G/A), rs11264799 (T/C), and rs6691569 (A/G).
The p-value was from logistic regression analysis adjusted for age and gender.
One thousand-fold permutation test.
Association of FCRL3 variants with clinical phenotypes and prognosis of IgAN
The associations between FCRL3 gene polymorphisms and clinical phenotypes as well as progression were further investigated in IgAN patients (Tables 3 and 4). The protective allele A of rs11264794 was associated with increased eGFR levels (β = 0.06, p = 0.046), low frequency of hypertension (OR = 0.80, p = 0.026) and hyperlipidemia (OR = 0.80, p = 0.043), and the reduced occurrence of tubular atrophy and interstitial fibrosis (OR = 0.79, p = 0.018). Similarly, the rs11264793-T allele was associated with a low frequency of hyperuricemia (OR = 0.74, p = 0.009) and the reduced occurrence of tubular atrophy and interstitial fibrosis (OR = 0.74, p = 0.011). Moreover, those subjects with the AA genotype of rs11264794 had the lowest Gd-IgA1 levels in IgAN patients (p = 0.026) (Supplementary Table S6).
Association Analysis of Fc Receptor-Like 3 Polymorphisms with Clinicopathological Parameters in IgA Nephropathy Patients
All the analyses adjusted for age and gender. Bold indicates statistical significance.
eGFR, estimated glomerular filtration rate.
Cox regression Analysis Between Fc Receptor-Like 3 Polymorphisms and the Risk of Composite Endpoint of IgA Nephropathy
Bold indicates statistical significance.
Model 1: Unadjusted.
Model 2: Adjusted for age, gender, hypertension, proteinuria, and eGFR.
HR, hazard ratio.
Of 971 IgAN patients, longitudinal retrospective progression data were available for 729 individuals with a median follow-up time of 27.8 (25th–75th percentile, 18.2–40.5) months. During the follow-up, 110 patients reached the endpoint. The Cox regression analysis showed that three FCRL3 variants (rs11264794, rs7522061, and rs7528684) were associated with favorable prognosis in IgAN patients, and rs11264794 was an independent protective factor for IgAN progression after controlling for the effects of age, gender, hypertension, proteinuria, and eGFR (adjusted hazard ratio = 0.64, 95% CI = 0.43–0.97, p = 0.033; under the dominant model) (Table 4). The renal survival rate in IgAN patients with the CC genotype of rs11264794 was lower than that in patients with the CA or AA genotypes (log-rank p = 0.032) (Fig. 2).
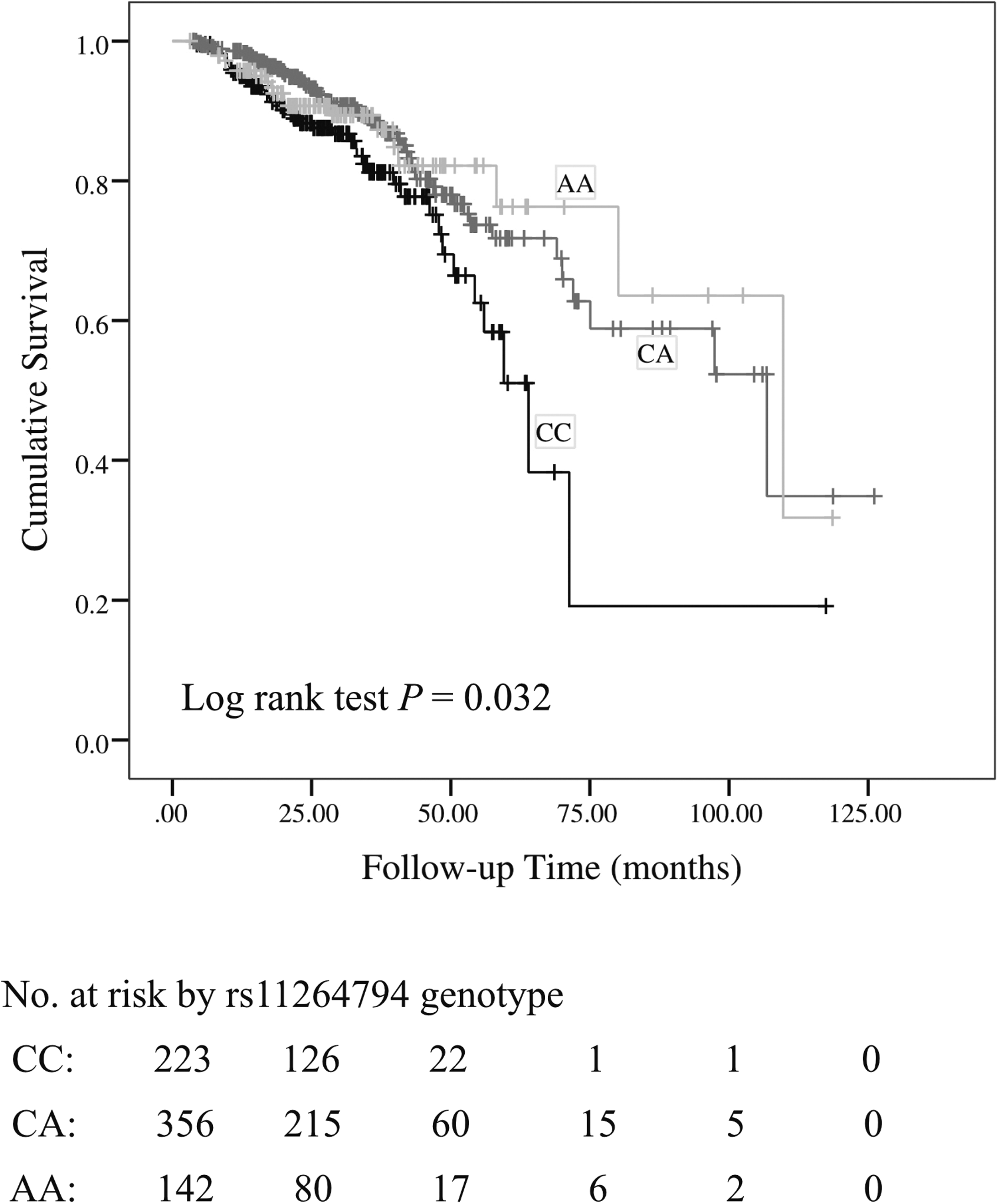
Influence of FCRL3 rs11264794 genotypes on renal survival in IgAN patients. The number of patients at risk at selected time points is shown below the plot (SNP rs11264794 AA genotype vs. CA genotype vs. CC genotype, p = 0.032, log-rank test). IgAN, IgA nephropathy.
Functional annotation of rs11264794
As SNP rs11264794 was both associated with the development and prognosis of IgAN, multiple bioinformatic databases were used to further get the functional annotation of this SNP. SNP rs11264794 was located in the 3′-UTR of FCRL3 (Supplementary Table S7), and two miRNAs (miR-183-5p.1 and miR-4470) were predicted to bind at the rs11264794 region according to TargetScan.
Comparison of FCRL3 gene expression levels in PBMCs among SNP rs11264794 genotypes
To investigate the possible effect of SNP rs11264794 genotypes on gene expression, the FCRL3 expression levels in PBMCs from 46 IgAN patients and 68 healthy controls with different genotypes were analyzed. The mRNA and protein expression levels of FCRL3 in IgAN patients were both significantly lower than those in healthy controls (p < 0.05) (Supplementary Fig. S2). When stratified by the rs11264794 genotypes, we found that heterozygous and homozygous genotypes of SNP rs11264794 had higher FCRL3 gene expression levels than those with wild-type genotypes (Supplementary Table S8).
Effect of rs11264794 on the binding of miR-183-5p.1 or miR-4470 to FCRL3 3′-UTR
Luciferase reporter assays were used to elucidate the mechanism underlying the effect of rs11264794 on the expression of FCRL3. As shown in Figure 3, we found that the 3′-UTR of FCRL3 containing the minor allele of rs11264794 combined with miR-183-5p.1 could significantly increase the luciferase gene expression.
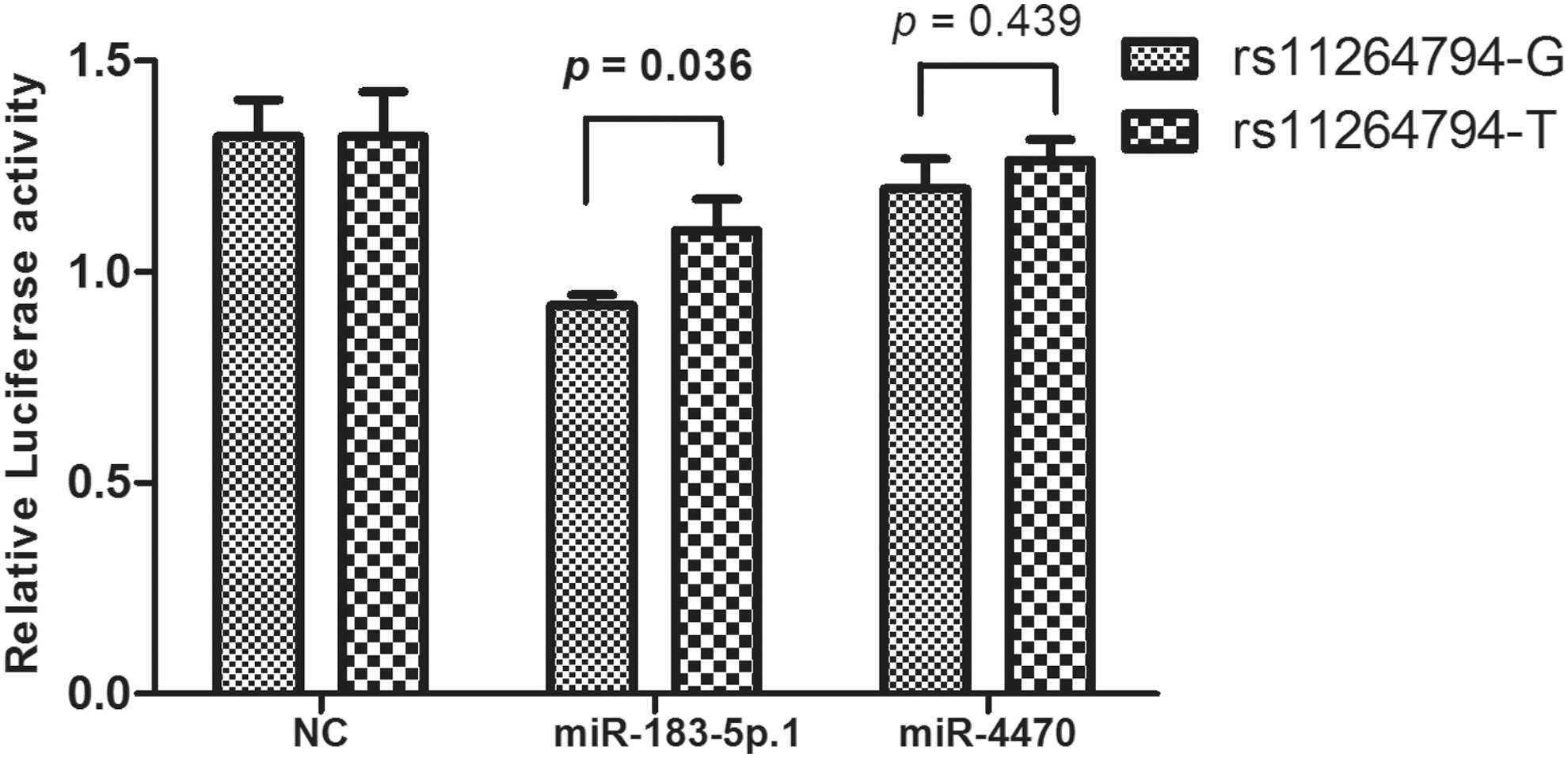
The effect of rs11264794 on miR-183-5p.1 and miR-4470: FCRL3 binding. Luciferase expression vectors containing the FCRL3 3′UTR with the rs11264794-G or -T allele were constructed and cotransfected with miRNA mimics (miR-183-5p.1 or miR-4470) in HEK293 cells. Data are presented as mean (standard deviation) of different groups. HEK293, human embryonic kidney 293; miRNA, microRNA; UTR, untranslated region.
Discussion
IgAN is a common primary glomerulonephritis and has a complex genetic etiology. Multiple gene polymorphisms have been reported to be associated with the development and/or progression of IgAN (Feehally and Barratt, 2015). In this two-stage case/control study, we found that four genetic variants (rs11264794, rs7528684, rs11264799, and rs6691569) of FCRL3 were remarkably associated with IgAN susceptibility. In addition, FCRL3 gene polymorphisms were also associated with the decreased severity and better prognosis of IgAN. SNP rs11264794 located in the 3′-UTR of FCRL3 had a potential effect on the regulation of FCRL3 gene expression through binding with miR-183-5p.1. To our knowledge, we have reported for the first time that FCRL3 gene polymorphisms are associated with the development and progression of IgAN.
Emerging lines of evidence have demonstrated that genetic variants of FCRL3 can influence the susceptibility to several autoimmune disorders (Rostamzadeh et al., 2018). Interestingly, the FCRL3 gene polymorphism exhibited a complex pattern of associations with immune diseases because it showed as a risk factor for rheumatoid arthritis (Kochi et al., 2005) and Graves' disease (Fang et al., 2016), while as a protective factor for IgAN, confirmed in our study, autoimmune Addison disease (Owen et al., 2007), and multiple sclerosis (Matesanz et al., 2008). The inconsistent associations of the same alleles further support the notion of pleiotropy that a single gene has an effect on multiple phenotypes (Zhou et al., 2014). Moreover, FCRL3 contains two motifs, ITAM and ITIM, that can activate or inhibit tyrosine protein kinase, resulting in activating or inhibiting cell signaling and causing bidirectional immune response (Davis, 2007).
FCRL3 gene polymorphisms were previously found to play important roles in the development of certain clinical phenotypes of autoimmune diseases (Chen et al., 2011; Brorsson et al., 2015). In this study, our results revealed that SNPs (rs11264794 and rs11264793) of FCRL3 were associated with increased eGFR level, less frequency of hypertension and hyperuricemia, lower occurrence of tubular atrophy and interstitial fibrosis, and decreased serum Gd-IgA1 level, which are all the clinicopathologic predictors for less disease severity or improved renal outcome of IgAN (Syrjanen et al., 2000; Maixnerova et al., 2016). Moreover, Cox regression further showed that the AA+CA genotype of rs11264794 was an independent protective factor for decline in renal function. Taken together, these findings indicated the involvement of FCRL3 gene variants in IgAN deterioration and progression.
SNP rs7528684 has been identified as a potential functional genetic candidate factor for a variety of autoimmune diseases (Kochi et al., 2005; Fang et al., 2016; Yuan et al., 2016) and demonstrated to modify a binding site for the nuclear factor-κB (NF-κB) as well as associated with increased FCRL3 mRNA expression (Kochi et al., 2005). The noncanonical NF-κB pathway has been implicated in the regulation of IgA class switching and developing IgAN (Zhang and Sun, 2015). It was possible that FCRL3 can stimulate immunoglobulin production through activating the downstream NF-κB. The most strongly associated SNP rs11264794 in this study was demonstrated to alter the binding ability of hsa-miR-183-5p.1 to 3′-UTR of FCRL3 and regulate the transcriptional level of FCRL3 gene. Moreover, our data revealed that the rs11264794-A allele was associated with increased expression levels of FCRL3 in PBMCs of IgAN patients and control subjects. These findings collectively highlight the regulatory effects of rs11264794 and rs7528684, as well as an important role of FCRL3 gene in IgAN.
FCRL3 belongs to the immunoglobulin receptor superfamily and plays a key role in the development, maturation, and function of B cells (Kochi et al., 2009). In in vitro studies, FCRL3 was demonstrated to act as a negative regulator of BCR signaling, and inhibit the production of immunoglobulin (Li et al., 2013). Besides, FCRL3 is reported to be expressed by Tregs, and FCRL3+ Treg can suppress the proliferation of effector T cells (Swainson et al., 2010). Therefore, FCRL3 has a functional role in the immune system and potentially implicated in the pathology of immune-related diseases. At present, IgAN is considered an autoimmune disease, and polymeric IgA1 or Gd-IgA1 produced from plasma cells are commonly found in IgAN patients (Al Hussain et al., 2017). Lower expression levels of FCRL3 in IgAN patients indicated that it may augment the production of autoantibodies through regulating the BCR signaling and function of Tregs.
Conclusion
We identified that the variants of FCRL3, particularly rs11264794, were significantly associated with the development and progression of IgAN in a Chinese Han population. In vitro studies demonstrated that rs11264794 showed a protective role for IgAN through transcriptional modulation of FCRL3. Our findings can provide more evidences of the involvement of FCRL3 in the pathogenesis of IgAN. However, the present work was only studied in the Chinese Han population, and further studies with larger sample sizes in different ethnicities will be needed to validate our results.
Footnotes
Acknowledgments
We are grateful to all the subjects and healthy volunteers who participated in this work. We thank Professor Jianjun Liu for reviewing the article and providing constructive suggestions.
Authors' Contributions
M.L., S.F., and Z.Z. designed the study and were responsible for the project management; Z.Z., D.S., R.X., P.Y., and M.W. collected the clinical data of patients, performed experiments, and analyzed the data; Z.Z., S.F., and D.S. wrote the article; and M.L., X.Y., H.M., F.H., and Z.L. provided critical revision of the article. All authors read and approved the final article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was funded, in part, by the National Key Research and Development Program, China (No. 2016YFC0906100), Operational Grant of Guangdong Provincial Key Laboratory, China (No. 2017B030314019), Guangdong Provincial Programme of Science and Technology, China (No. 2017A050503003; No. 2017B020227006), Guangzhou Municipal Programme of Science and Technology, China (No. 201704020167; No. 2016201604030005; and No. 201807010002), National Natural Science Foundation of China (No. 81770661; No. 81570599; and No. 81570750), and the Young and Middle-aged Talents Program of The First Affiliated Hospital, Sun Yat-sen University.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Supplementary Table S7
Supplementary Table S8
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.