
Abstract
Lung adenocarcinoma (LUAD) is the most common subtype of nonsmall cell lung cancer, and 5-year survival rate is only 15% in recent years. This study aimed to explore the FAM83A expression and its potential functions in LUAD. Data of LUAD were downloaded from The Cancer Genome Atlas database. Expression level of FAM83A was compared between LUAD samples and adjacent normal samples. The association between FAM83A expression and clinic-pathological parameters was analyzed, as well as copy number variation and methylation status. Kaplan–Meier curve was used to visualize the relationship of FAM83A expression with survival outcomes. Finally, gene set enrichment analysis was used to identify potential signaling pathways in LUAD specimens. FAM83A expression was significantly correlated with four clinical factors in LUAD specimens, age, gender, smoking, and overall survival status (all p < 0.05). High expression level of FAM83A was negatively correlated with methylation level. Moreover, patients in low expression groups exhibited a better prognosis than those in high expressed groups, which was independent of gender (p < 0.001). Histidine metabolism pathway was significantly upregulated in FAM83A-high expressed samples than FAM83A-low expressed samples according to functional enrichment analysis. High expression of FAM83A predicted a poor prognosis in LUAD patients. Our study demonstrated that FAM83A might be a potential biomarker and meaningful therapeutic target in LUAD.
Introduction
Lung cancer is still a leading cause of cancer-related deaths, and there is a globally increasing incidence and mortality in recent years (Chen et al., 2017; Barta et al., 2019). Approximately 228,150 new cases are diagnosed as lung cancer in United States, and 142,670 individuals die of this disease (Siegel et al., 2019). The main types of lung cancer include small cell lung carcinoma and nonsmall-cell lung carcinoma (NSCLC). Lung adenocarcinoma (LUAD) is the most common subtype of NSCLC characterized by distinct cellular and molecular features (Travis et al., 2015).
Various factors result in pathogenesis of LUAD, including genetic and environmental effects, whereas tobacco smoking acts as a major risk factor and is responsible for over 80% of all lung cancer cases globally (Testa et al., 2018). Although tremendous progresses have been made in cancer therapy, the 5-year survival rate of LUAD is still unsatisfied and many individuals are diagnosed at advanced stages, which lead to a heavy disease burden in public (Gettinger et al., 2018). Therefore, it is an emerging task to identify novel biomarkers for early detection and prognosis prediction in LUADs.
Substantial efforts have been devoted to the diagnosis of LUAD. It was reported that seven microRNAs, including chr17_10932, has-miR-210-3p, has-miR-199a-3p, has-miR-138-5p, chr1_1402, has-miR-148a-3p, and has-miR-378d, were differentially expressed between LUAD tumor and paracancerous tissues, which could be adopted for the miRNA panel for LUAD diagnosis (He et al., 2017). Circular RNA hsa_circ_0013958 was found to be overexpressed in LUAD tissues, which played a key role in enhancing cell proliferation and invasion, suppressing cell apoptosis, and could be used as a potential screening tool of LUAD (Zhu et al., 2017).
In addition, several researches have attempted to search for the prognosis-related biomarkers of LUAD. Shi et al. found that the upregulation of cell division cycle 20, a critical element of spindle assembly checkpoint, was in association with inferior prognosis of LUAD patients and could be applied to the identification of cases with high risk (Shi et al., 2017). A previously reported study showed that long noncoding RNA SOX21-AS1 was implicated in the development of LUAD by influencing the tumor size and tumor-node-metastasis stage and considered as a potential predictor of prognosis (Lu et al., 2017). Despite the significant advances in the diagnostic and prognostic biomarkers of LUAD, extensive clinical work is needed for their pervasive applications in routine clinical settings.
FAM83A, also known as BJ-TSA-9, is a member of FAM83 family and located on chromosome 8q24 (Chen et al., 2017). FAM83A has been reported to be implicated in the occurrence and development of multiple cancers. For example, Chen et al. (2017) showed that FAM83A overexpression could promote the development of pancreatic cancer (Chen et al., 2017). Cipriano et al., (2014) found that the expression level of FAM83A was significantly elevated in the tumor tissues of ovarian and bladder cancers compared with the corresponding normal tissues (Cipriano et al., 2014). In addition, aberrant expression of FAM83A was suggested to involve in the chemoresistance of breast cancer through interacting with epidermal growth factor receptor/phosphatidylinositol 3 kinase/AKT signaling pathway (Grant, 2012). However, to our knowledge, the association of FAM83A with the carcinogenesis, development, and prognosis of LUAD is rarely studied.
In this study, we downloaded the LUAD datasets from The Cancer Genome Atlas (TCGA) database and conducted a comprehensive analysis for these LUAD samples. We found that the expression level of FAM83A was generally higher in LUAD samples compared with adjacent normal samples. Several clinic-pathologic factors were correlated with up-expression of FAM83A, such as age, gender, and OS status. According to Kaplan–Meier analysis, high expression level of FAM83A was associated with poor prognosis in LUAD patients, and histidine metabolism pathway was significantly activated in FAM83A-high expression samples. Thus, FAM83A might serve as a promising target with prognostic values in LUAD patients. Our study might promote a further understanding for LUAD development.
Materials and Methods
Data resource
The data associated with LUAD were downloaded from TCGA database, including 592 tumor samples and 59 adjacent normal tissue samples. Moreover, 575 tumor samples were labeled with tumor stage, while 524 samples were accompanied by survival outcomes and survival status. Two cohorts from the Gene Expression Omnibus (GEO, Access numbers: GSE31210, GSE50081) were used as the validation sets for survival analysis, which included 246 and 181 LUAD samples, respectively.
Differential expression analysis
Genes with an average count of less than 10 were removed. The edgeR function package (Robinson et al., 2010) in R software was used to screen differentially expressed genes (DEGs). The false discovery rate <0.05 and |log2FC (fold change) | > 1 were considered as thresholds.
Correlation analysis
Copy number variation (CNV) and aberrant methylation level of genes are involved in pathogenesis and development of various cancers. In this study, we analyzed the correlation between FAM83A expression and CNVs, as well as the methylation status in LUAD. First, the copy number alterations were evaluated using Genomics Identification of Significant Targets in Cancer 2.0 (GISTIC 2.0) (Mermel et al., 2011). In this study, we calculated the segment score of each gene and defined it as −1, 0, 1, which represented deletion, normal copy number, and amplification, respectively. The probes of 450 k methylation array data were distributed in different regions of the genome, such as the promoter, 5′UTR, 3′UTR, and Body. The DNA methylation data were analyzed by Illumina GenomeStudio Software, and the β values were obtained. The β > 0.6 indicated fully methylated status, β < 0.2 indicated fully unmethylated status, and 0.2 < β < 0.6 indicated partially methylated status. For the genes that might have multiple probes of methylation, the mean values were used. The correlation between FAM83A expression and its methylation status was calculated by Pearson correlation analysis. p < 0.05 was regarded as the cutoff for indicating significance. r < 0 indicated negative correlation, and r > 0 indicated positive correlation.
Gene set enrichment analysis
In this study, gene set enrichment analysis (GSEA) was used to perform function analysis for cohorts of genes. The profiles c2.cp.kegg.v6.2.symbols.gmt, which were downloaded from the website of GSEA and included 5501 genes and their function information, were considered as gene set, and normal value p < 0.05 was set as a threshold. The p values were adjusted by multiple hypothesis test of Benjamini and Hochberg method.
Quantitative real-time PCR
Total RNA was extracted using TRIzol (Invitrogen). Reverse transcription was done using cDNA Reverse Transcription Kit for mRNAs. The expression of mRNAs was measured using TaqMan Gene Assay specific for FAM83A and Formimidoyltransferase Cyclodeaminase (FTCD). FAM83A-siRNA and negative control siRNA were purchased from Qiagen. The experiments were performed in triplicate.
Proliferation assay by lactate dehydrogenase activity measurement
Cells were quantified by measuring the lactate dehydrogenase activity with a Cytotoxicity Detection Kit according to the manufacturer's instructions (Roche, Germany). Briefly, the cells that were treated with FAM83A-siRNA were lysed and centrifuged for 3 min at 12,000 rpm. The supernatant was then further processed. The assay was repeated thrice.
Statistical analysis
The chi-square test and two-sided t test were used to analyze the correlation between corresponding clinical characteristics and FAM83A expression in LUAD patients. Besides, two-sided t test and analysis of variance tested were used to evaluate the expression of FAM83A in LUAD and adjacent tumor samples. Kaplan–Meier analysis was performed to estimate the overall survival (OS) (Goel et al., 2010). The log-rank test was used to explore OS differences among multiple groups of samples. The correlation between FAM83A expression and methylation was calculated by Pearson correlation method (Sedgwick, 2012). p < 0.05 was considered as a threshold.
Results
Relationship analysis between FAM83A expression and clinicopathologic features
First, the LUAD data were normalized, and expression values of FAM83A were preprocessed. The LUAD samples were divided into two groups after setting median expressed value of FAM83A as cutoff point. The results are shown in Table 1, and the p values in Table 1 were analyzed by chi-square test, which were not adjusted by multiple hypothesis tests. FAM83A expression was significantly correlated with four clinical factors, including age, gender, smoking, and OS status (all p < 0.05). In addition, there was no significant difference in pathologic stage of patients between FAM83A high expression group and FAM83A low expression group.
The Correlation Analysis Results Between FAM83A Expression and Clinic-Pathological Factors in Lung Adenocarcinoma Patients
OS, overall survival.
FAM83A was overexpressed in LUAD samples
We compared the expression value of FAM83A in LUAD tissues and adjacent tissue samples and found that the expression level of FAM83A was significantly higher in LUAD tissue samples compared with that in adjacent tissue samples (Fig. 1A, p < 0.001). However, there was no significant difference in FAM83A expression among patients with different tumor stages (Fig. 1B). Besides, FAM83A expression levels in LUAD tissue samples and adjacent tissue samples from validation dataset GSE31210 were analyzed. As shown in Supplementary Figure S1, the expression level of FAM83A in LUAD tissue samples was significantly higher than that in adjacent tissue samples. The results preliminarily indicated that FAM83A played a major role in LUAD development and might be highly expressed in the early stage of cancer, as well as the whole process of disease.
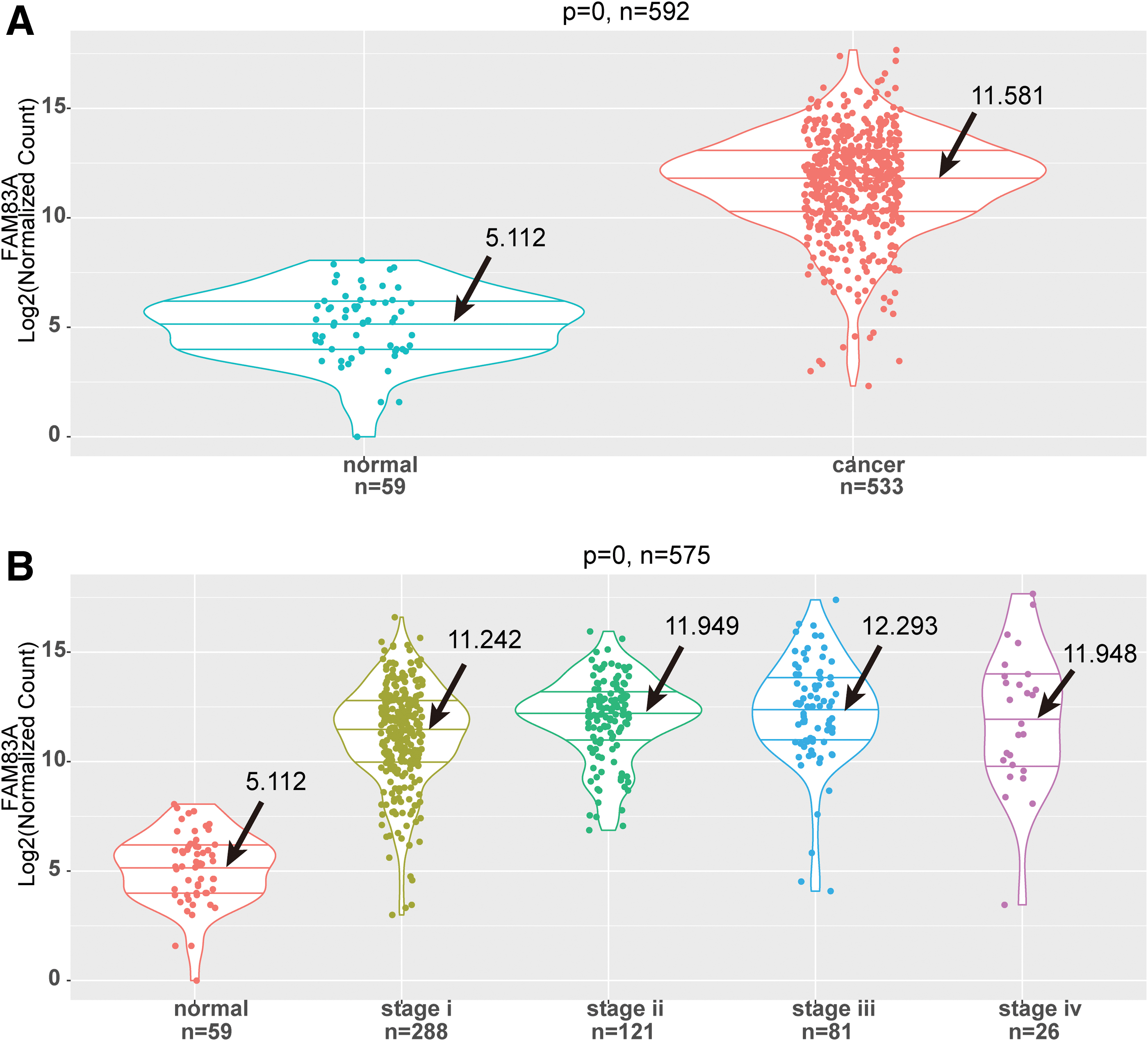
High expression of FAM83A in LUAD specimens compared with adjacent normal specimens.
FAM83A expression was negatively correlated with methylation level
On the whole, the FAM83A expression in LUAD samples was negatively correlated with its methylation level according to Pearson correlation analysis (Fig. 2A, r < 0, p < 0.01). There were a total of three CpGs located at the promoter region of FAM83A, and we also explored correlations between methylation level of every of those three CpGs and FAM83A expression. As a result, all the three CpGs were negatively correlated with FAM83A (p < 0.01), and the correlation coefficient of cg02715629 achieved −0.53 (Supplementary Fig. S2). As for the LUAD samples with amplified copy number, there was no significant correlation between FAM83A expression and CNV (Fig. 2B). The results suggested that aberrant expression of FAM83A might play a major role in pathogenesis of LUAD which involved in multiomics.
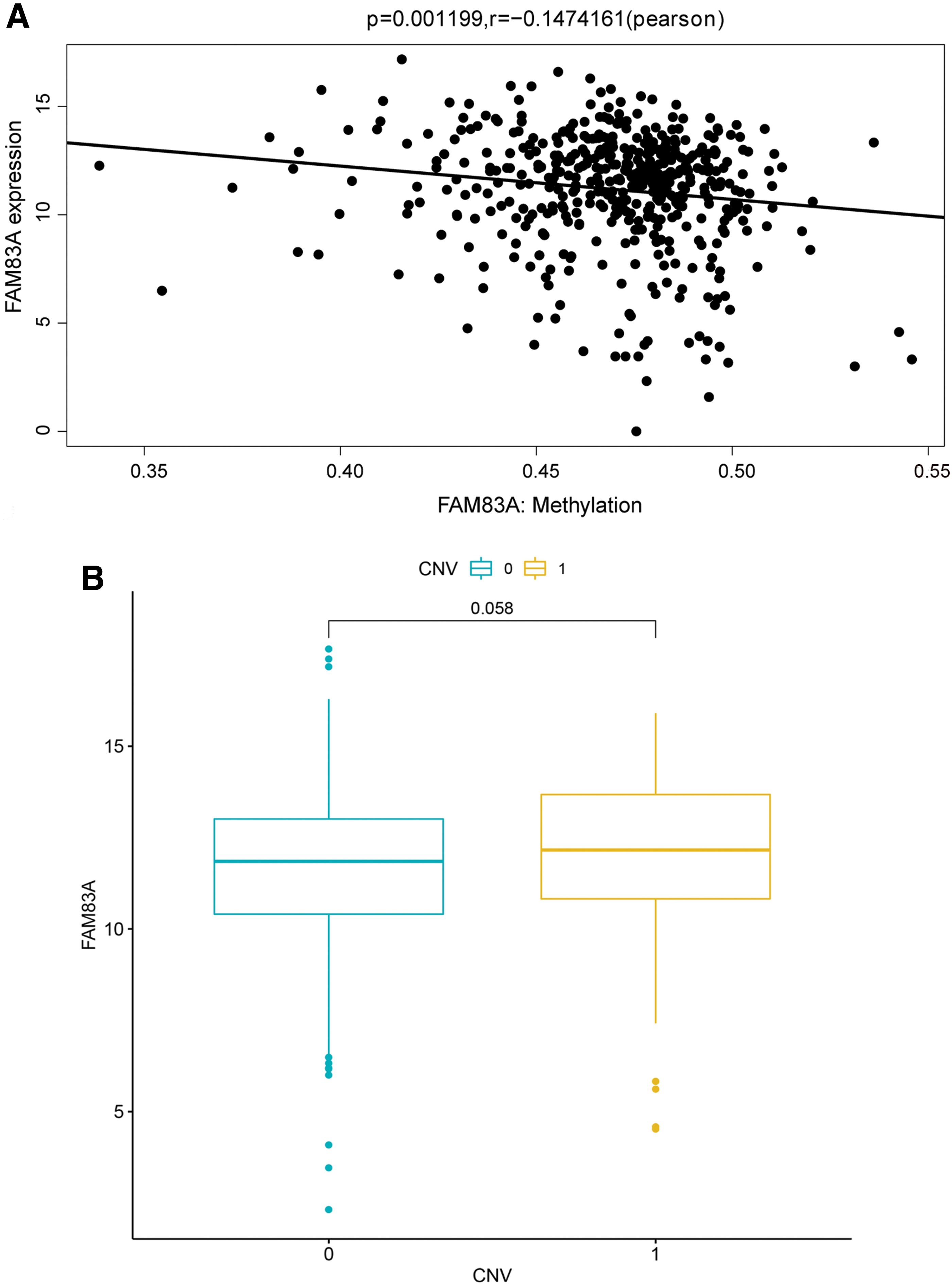
Correlation analysis of FAM83A expression with methylation level and CNV.
High expression level of FAM83A was associated with poor prognosis in LUAD patients
After setting the median expression value of FAM83A as a threshold, LUAD samples from TCGA were divided into two groups. Kaplan–Meier analysis showed that patients in low expression groups presented a better prognosis than those in high expressed groups (Fig. 3A, p < 0.001). Then Cox regression analysis was performed, which showed that FAM83A expression could affect the prognosis of LUAD patients independent of multiple confounding variables, including age, gender, tumor stage, TP53 mutation, and smoking status (Fig. 3B). The prognostic function of FAM83A was further validated in datasets GSE31210 and GSE50081 from GEO, which also showed that LUAD patients with high FAM83A expression level had worse OS than those with low FAM83A expression level (p = 0.0047, p = 0.0094, respectively, Fig. 4). These results indicated that abnormal expression of FAM83A might be related to prognosis in LUAD patients regardless of several confounding factors such as age, gender, tumor stage, TP53 mutation, and smoking status.
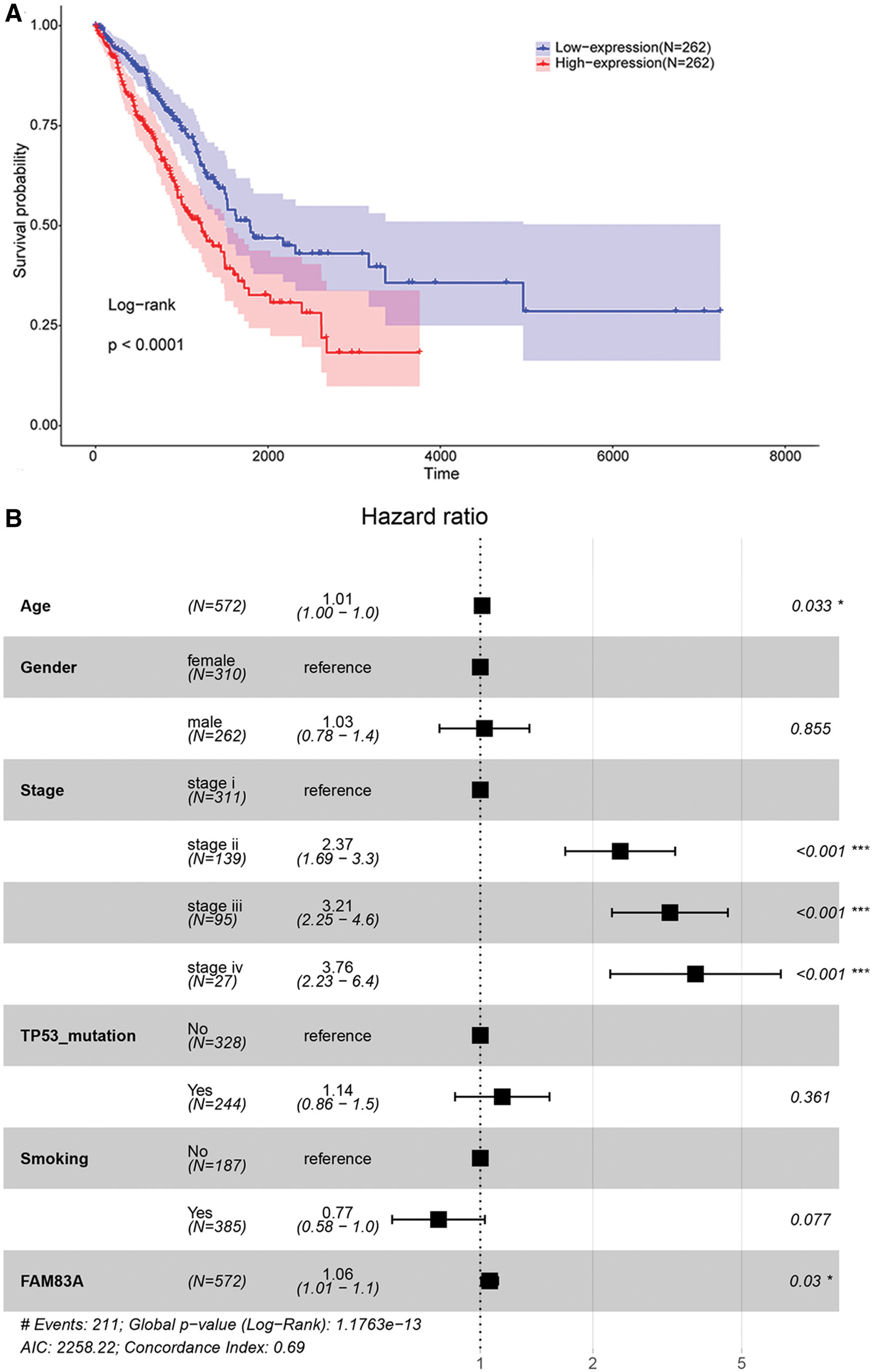
Survival analysis of LUAD patients from TCGA.
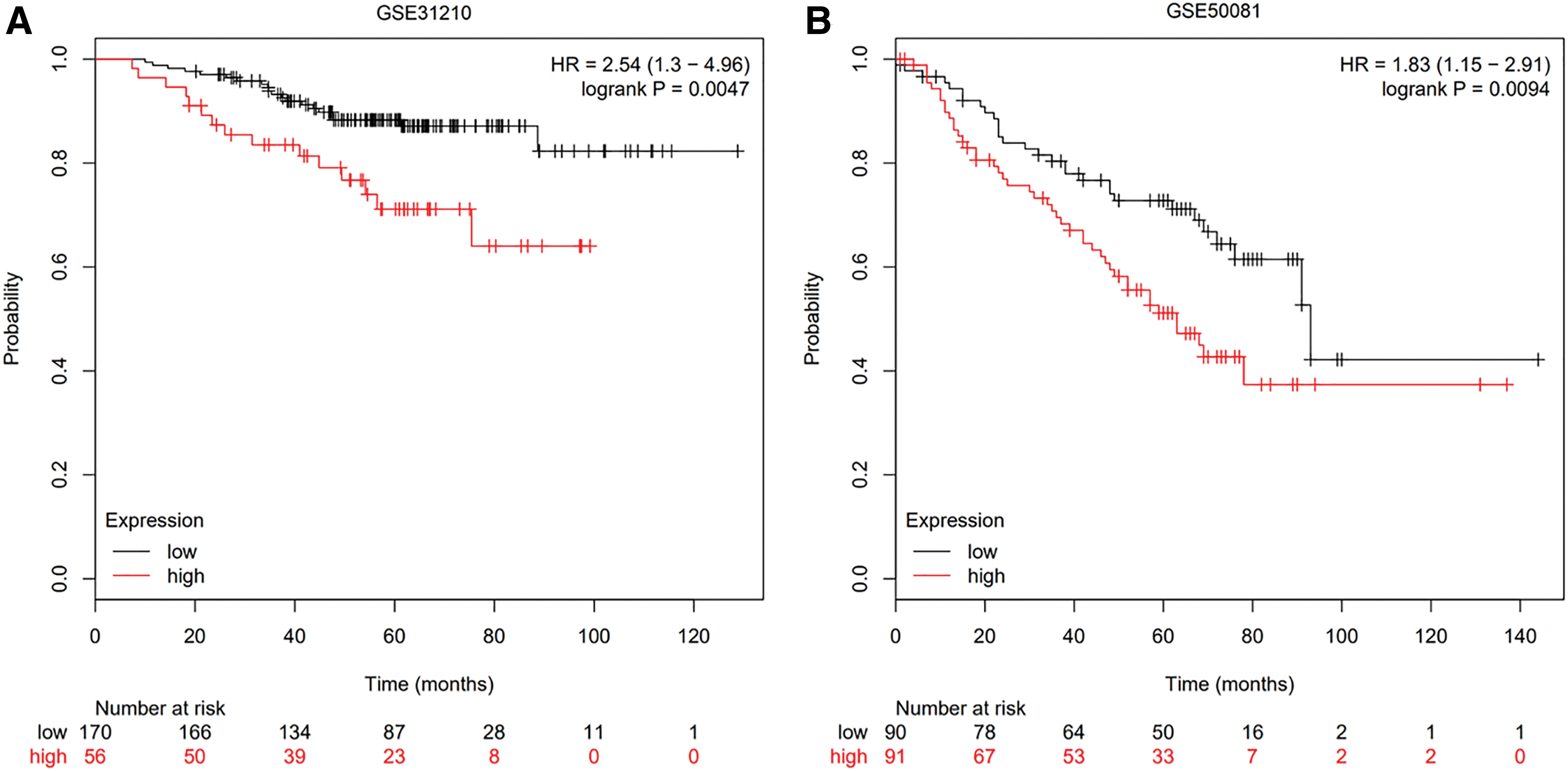
Survival analysis of LUAD patients from GEO.
Elevated FAM83A expression may correlate with poor prognosis in LUAD patients by activating histidine metabolism pathway
To explore the potential mechanism of FAM83A overexpression in LUAD patients, the DEGs were identified from top 10 samples with higher expressed FAM83A (FAM83A_High) compared with 10 samples with lower expressed FAM83A (FAM83A_Low). A total of 1030 DEGs were identified between these 20 samples. The heatmap of these DEGs is displayed in Figure 5A. GSEA was further conducted to investigate the signaling pathways of these DEGs involved. Based on the criteria of p < 0.05, we found that histidine metabolism was significantly activated in FAM83A_High group samples (Fig. 5B). Thus, we speculated that FAM83A might be involved in pathogenesis and development of LUAD through the upregulation of histidine metabolism pathway. The top 10 most significant KEGG pathways are provided in Supplementary Table S1.
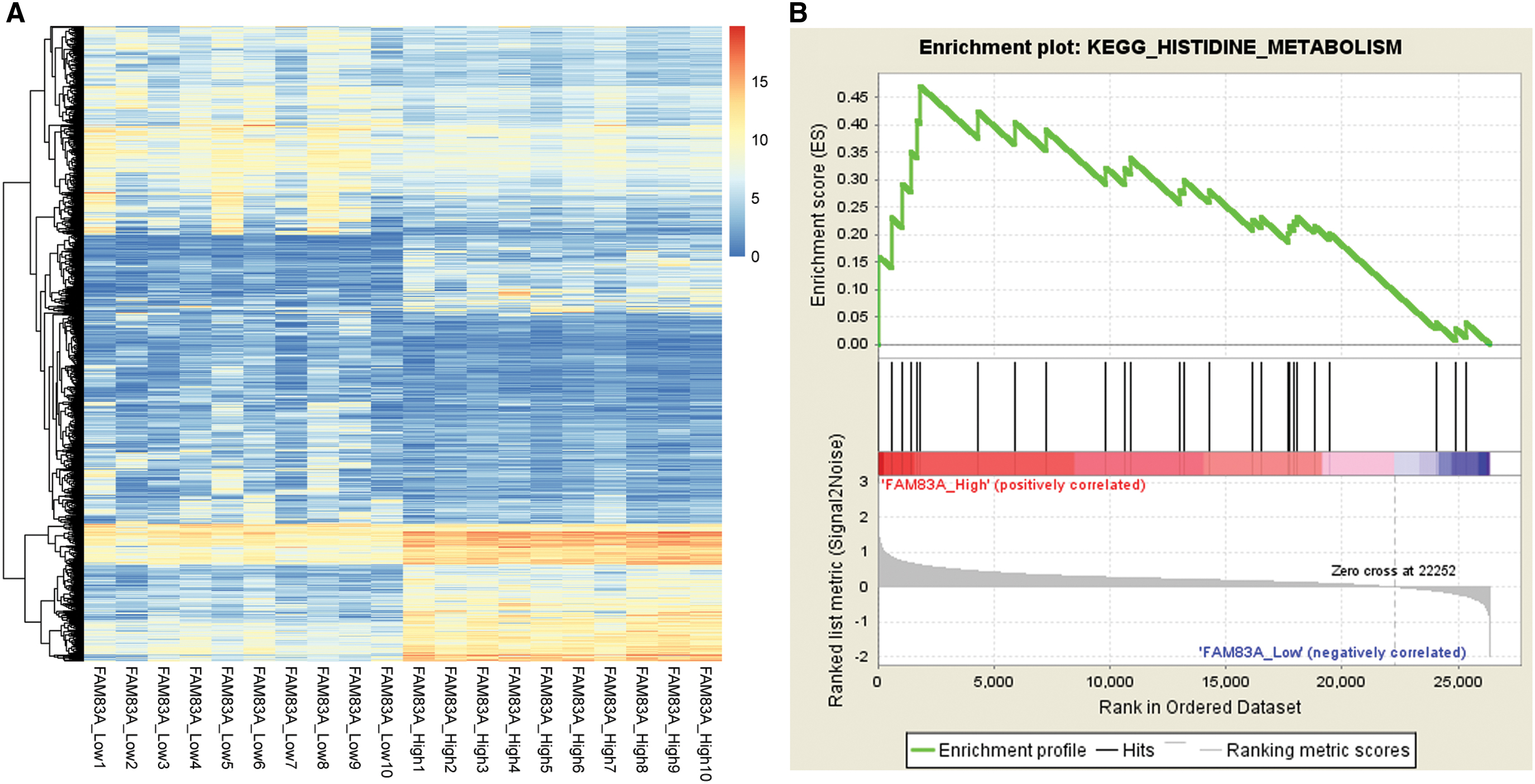
FAM83A expression was correlated with upregulation of histidine metabolism pathways.
FAM83A was involved in cell proliferation in LUAD through histidine metabolism pathway
To investigate the function of FAM83A and histidine metabolism pathway, quantitative real-time PCR and proliferation were carried out in LUAD transferred with FAM83A-siRNA by liposome 2000. As shown in Figure 6A and B, declined FAM83A expression by siRNA significantly reduced the expression of FTCD, which led to obvious inhibition of cell proliferation (Fig. 6C). This result demonstrated that FAM83A could inhibit the cell proliferation through histidine metabolism pathway downregulation.

Investigation of the role of FAM83A and histidine metabolism pathway in LUAD.
Discussion
In this study, we identified that overexpression of FAM83A was prevalent in LUAD samples compared with adjacent normal samples. According to correlation analysis, high expression of FAM83A was significantly correlated with three clinic-pathologic factors, age, gender, and OS status. Furthermore, the FAM83A expression in LUAD samples was negatively correlated with methylation level. Histidine metabolism pathway was significantly activated in FAM83A high expressed samples based on functional enrichment analysis.
FAM83A belongs to the FAM83 family. The proteins in FAM83 (family with sequence similarity 83) family were characterized by conserved DUF1669 domain in N-termini, and this domain consisted of a conserved phospholipase D-like catalytic motif (Bozatzi and Sapkota, 2018). Previous studies have reported that FAM83 family oncogenes were broadly involved in human cancers. For example, FAM83B was upregulated in breast cancer, lung carcinoma, ovary cancer, and lymphoid cancer (Okabe et al., 2015). Overexpression of FAM83B was involved in EGFR- and RAS- driven oncogenic transformation in breast cancer (Cipriano et al., 2012).
FAM83D, acting as a member of FAM83 family, could promote cell proliferation and migration in MCF10A breast cells (Walian et al., 2016). FAM83D could also activate MEK/ERK signaling pathway in hepatocellular cell lines (Wang et al., 2015). Indeed, BJ-TSA-9 or FAM83A was originally identified as a tumor-specific biomarker and was highly expressed in nearly half of lung cancer tissue samples (21/40) (Li et al., 2005). FAM83A presented diagnostic value in differentiating lung cancer patients compared with healthy controls (Liu et al., 2008). Moreover, a clinical meta-analysis research revealed that high FAM83A expression was related to poor prognosis in breast cancer patients (Lee et al., 2012).
Based on these findings, FAM83A acted as a candidate tumor gene with prognostic value in some cancer types, whereas the biological function of FAM83A in cancer progression was still unclear. Lee et al., (2012) reported that overexpression of FAM83A could promote proliferation and invasion in breast cancer cells and resulted in EGFR-TKI resistance both in culture and in vivo. The molecular mechanism might be associated with phosphorylation of c-RAF and PI3K, members of RAS/ERK and PI3K/mTOR pathway (Lee et al., 2012). Our study had first identified that high expression level of FAM83A predicted a poor prognosis in LUAD patients. Together with previous studies, we suggested that abnormal expression of FAM83A might be involved in RAS or mTOR pathways in LUAD progression.
In this study, high expression of FAM83A was positively correlated with four clinical factors (age, gender, smoking, and OS status). On the contrary, there was no significant correlation-ship among FAM83A expression and tumor stage. The results indicated that overexpression of FAM83A occurred in the early stage of LUAD and exhibited a higher expressed status throughout the whole tumor stages. Thus, high expression of FAM83A might serve as a potential biomarker for early detection of disease. Furthermore, Kaplan–Meier analysis results showed that patients in low expression groups presented a better prognosis than those in high expressed groups regardless of the gender (Fig. 3, p < 0.001).
After literature retrieval, we found that association of FAM83A expression and clinic-pathogenic factors has not been reported in lung cancer. Interestingly, Shen et al. (2017) showed that higher expression of FAM83B, another member of FAM83 family, predicted a poor prognosis in pancreatic ductal adenocarcinoma patients. The upregulation of FAM83B was positively associated with tumor stage, regardless of lymphatic metastasis and histological features.
In addition, aberrant amino acid metabolism was an emerging hallmark of cancer, whereas the molecular mechanisms were poorly explored (Hanahan and Weinberg, 2011; Pavlova and Thompson, 2016). Generally, the mammalian cells increased the import of nutrients from environment to fulfill the biosynthetic demands for cell proliferation. Glucose and glutamine act as two principal nutrients that support survival and biosynthesis in normal mammalian cells. However, aberrant activation of oncogene and loss of tumor suppressor could deregulate the uptake of glucose and amino acids into cancer cells (Warburg et al., 1927; Sauer et al., 1982).
Tsuber et al., (2017) analyzed large cohorts of tumor tissue samples and found that mutations in cancer can cause gain of several amino acids, including histidine (Tsuber et al., 2017). Increasing level of histidine, glucose, and lipids in serum profiles was also observed in breast cancer patients and strongly correlated with disease relapse (Tenori et al., 2015). Recent study showed that histidine degradation can boost cancer therapy, and diet rich in the amino acid histidine increases the effectiveness of methotrexate treatment and lowers toxicity in mice (Naama et al., 2018). Although the abnormal uptake or gains of histidine were universally described in tumor cells, the role of histidine metabolism in cancer progression was rarely reported.
In this study, the functional analysis in our study revealed that histidine metabolism pathway was significantly upregulated in FAM83A-High group specimens (Fig. 5). A recent study identified that mTORC2 can regulate amino acid metabolism in cancer cell by phosphorylating cystine-glutamate antiporter xCT (Gu et al., 2017), and the glutamate secretion was subsequently suppressed. Considering this, whether the FAM83A expression regulated histidine metabolism through mTOR pathway should be verified in experiments.
However, there are still limitations in our study. The LUAD sample size in microarray datasets is small, and more tumor samples are needed to verify our results. In conclusion, this present study showed that high expression level of FAM83A might predict a poor prognosis in LUAD patients and might be involved in progression of LUAD through upregulation of histidine metabolism pathway. Our findings revealed that FAM83A might act as a potential prognostic biomarker and therapeutic target in LUAD.
Footnotes
Authors' Contributions
J.Y. made substantial contributions to conception and design, acquisition of data, and analysis and interpretation of data. M.H. performed the experiments and was involved in drafting the article or revising it critically for important intellectual content; T.P. had given final approval of the version to be published. T.P. agreed to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Disclosure Statement
No competing financial interests exist.
Funding Information
Funding was not received.
Supplementary Material
Supplementary Table S1
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.