
Abstract
Extracellular matrix metalloproteinase inducer (EMMPRIN) had been reported to be involved in the occurrence and development of coronary heart disease (CHD) in previous studies. This study aimed to investigate whether single nucleotide polymorphisms of EMMPRIN and matrix metalloproteinase-9 (MMP-9) contributed to the onset and severity of CHD. One thousand seventy patients suspected to have CHD were enrolled into the study. Each patient had undergone coronary angiogram, and the severity of coronary artery stenosis was assessed by Gensini score. Eight hundred twelve patients were confirmed to have CHD, while 258 patients were selected as non-CHD control. All patients were genotyped for five EMMPRIN polymorphisms (rs8259, rs28915400, rs4919859, rs6758, and rs8637) and one MMP-9 polymorphism (rs3918242) by polymerase chain reaction-restriction fragment length polymorphism and confirmed by direct sequencing. EMMPRIN polymorphism rs8259 and MMP-9 polymorphism rs3918242 were found to be associated with CHD (rs8259: AT vs. AA, adjusted odds ratio [OR] = 2.038, adjusted 95% confidence interval [CI] = 1.080–3.847, p = 0.028; rs3918242: CT vs. CC, adjusted OR = 0.607, adjusted 95% CI = 0.403–0.916, p = 0.017, TT vs. CC, adjusted OR = 2.559, adjusted 95% CI = 1.326–4.975, p = 0.006). No crossover effects were observed although a single environmental or genetic factor had an impact on the occurrence of CHD. The value of the Gensini score revealed that severity of CHD decreased in the rs3918242 CT carriers in both the male and female population. Our study suggested that EMMPRIN rs8259 and MMP-9 rs3918242 polymorphisms may contribute to pathological process of CHD. It could play a critical role in the prediction of CHD.
Introduction
Coronary heart disease (CHD), also known as ischemic heart disease, one of the main underlying causes of death worldwide, was defined as a progressive inflammatory disease of the vessel wall. However, the exact pathogenesis of CHD was not fully understood at present. Studies had indicated that interaction between environmental and genetic risk factors may have potential and intricate impact on occurrence of CHD (Zhao et al., 2018; Hu et al., 2019; Zhang et al., 2019). People gradually realized that CHD as a chronic disease was the consequence of long-term interaction of multiple risk factors.
Extracellular matrix metalloproteinase inducer (EMMPRIN), also known as basigin or CD147, was a single pass type 1 transmembrane glycoprotein that belongs to the immunoglobulin superfamily. EMMPRIN was first described in a fibroblasts-tumor cell coculture assay as the stimulating factor of matrix metalloproteinase 1 (MMP-1) (Biswas, 1982), and was found widely expressed in various cells, including platelets, smooth muscle cells, monocytes, and tumor cells (Schmidt et al., 2008; Chen et al., 2009). One of the most critical roles of EMMPRIN was to participate in the synthesis of MMPs, including MMP-1, MMP-2, MMP-3, MMP-9, and MMP-14 (Egawa et al., 2007; von Ungern-Sternberg et al., 2018). Matrix metalloproteinase-9 (MMP-9), known as an enzyme that degrades mainly type IV collagen and elastin (Van Doren, 2015), was one of the most important downstream factors of EMMPRIN. It was well elucidated that EMMPRIN could regulate the expression of MMP-9 mainly through adenosine 5′-monophosphate-activated protein kinase (AMPK)-mitogen-activated protein kinase (MAPK) signaling pathway in macrophages (Kim et al., 2009; Cao et al., 2014; Lin et al., 2018), and eventually played a role in a series of pathophysiological processes such as tumor metastasis, foam cell formation, plaque progression and destabilization, and migration of inflammatory cells (Seizer et al., 2010; Redzic et al., 2013; Cao et al., 2014).
The EMMPRIN/MMP-9 pathway had been extensively studied in the pathogenesis and metastasis of various cancers, including glioma, malignant melanoma, hepatocellular carcinoma, and gastrointestinal cancer (Huang et al., 2016; Hu et al., 2017; Li et al., 2017a; Peng et al., 2017). In addition, EMMPRIN/MMP-9 pathway had been found to play an important role in many inflammation-related diseases, including inflammatory periapical lesion (Wang et al., 2019), arthritis (Luan et al., 2018), chronic airway inflammation (Rossi et al., 2017), and even thromboinflammation-induced acute ischemic stroke (Jin et al., 2017). Asgari et al. (2019) confirmed that EMMPRIN could be an apoptosis regulator in spermatogenesis by promoting the expression of MMPs, which indicated that EMMPRIN may participate in reproduction, cell proliferation, and apoptosis. We can see that the potential value of EMMPRIN/MMP-9 pathway on pathogenesis of diverse diseases was still immeasurable, and further research was required.
The role of EMMPRIN in the occurrence and development of CHD had gradually been explored for decades. Studies indicated that EMMPRIN may play a vital role in coronary artery disease for its function in plaque formation (Wang et al., 2012; Yang et al., 2014), foam cells formation (Seizer et al., 2010), monocyte differentiation and recruitment (Major et al., 2002; Schulz et al., 2011). Inflammatory cytokine and MMP-9 released by EMMPRIN homophilic interactions or binding to a variety of ligands (Grass and Toole, 2015) were relevant to complex process of atherosclerosis, acute atherothrombosis, and other inflammatory or immune diseases (Pennings and Kritharides, 2014).
Also, serum EMMPRIN had been reported to be significantly higher in the myocardial infarction (MI) group (Akkus et al., 2016), which indicated that EMMPRIN may have the potential to predict and diagnose acute coronary syndrome (ACS). The mechanism of the regulation pathway was not clearly elucidated at present. In recent years, researchers had paid a considerable attention to the relationship between EMMPRIN gene polymorphisms and CHD. Several studies that based on the Chinese population indicated that EMMPRIN rs8259 polymorphisms played a vital role in ACS, heart failure by regulating the expression of EMMPRIN itself (Mao et al., 2014; Yan et al., 2015; Li et al., 2017b). However, the role of EMMPRIN gene polymorphisms in CHD was still controversial. In addition, research on the Chinese Han population was still limited at present. In this study we evaluated whether genetic polymorphisms of EMMPRIN pathway compositions, including EMMPRIN and MMP-9, were associated with the susceptibility and severity of CHD in the Chinese Han population.
Materials and Methods
Study population
A total of 1,070 hospitalized patients suspected of having CHD who underwent coronary angiography were enrolled in the study. CHD was diagnosed according to the World Health Organization guidelines. Exclusion criteria for this study were as follows: patients (1) suffering from other cardiovascular diseases other than CHD, including acute or chronic congestive heart failure, stage 2 or 3 hypertension, cardiomyopathy, infective endocarditis, rheumatic heart disease, myocarditis, congenital heart disease; (2) with severe trauma and burns; (3) with malignant tumor; (4) taking hormonal drugs recently; and (5) with connective tissue diseases.
Among them, 812 patients were diagnosed with CHD after examination, while 258 non-CHD patients were chosen as the control group. Two milliliters peripheral venous blood were collected into an ethylene diamine tetraacetic acid (EDTA) tube and stored in −20°C before coronary angiography. Basic information of all patients, including age, sex, body mass index (BMI), blood pressure, diabetes history, smoking history, alcohol drinking history, and a serum biochemistry index, was obtained from the hospital medical records. All subjects were unrelated ethnic Han Chinese with no mental disorders and everyone signed informed consent. The design of this study was in line with the Helsinki Declaration and ethics document, which was approved by the Ethics Review Committee of Hospital. Ethical approval was obtained before the start of the study.
Assessment of the severity of coronary artery stenosis
We used improved Gensini score to assess the severity of coronary stenosis. Coronary angiography images of all participants were evaluated by at least two attending physicians. The Gensini score was dependent on the location and severity of the stenosis. The scores of different coronary artery parts were as follows: left main coronary artery [5.0], proximal-middle segments of left anterior descending (LAD) coronary artery [2.5], distal segment and embranchments of LAD [0.5], proximal-middle segments of left circumflex artery (LCX) [2.5], distal segment and embranchments of LCX [0.5], and right coronary artery (RCA) [1.0]. The scores of different artery stenosis severity were as follows: <25% [1.0], 25% to ∼<50% [2.0], 50% to ∼<75% [4.0], 75% to ∼<90% [8.0], 90% to ∼<99% [16.0], and >99% [32.0]. The total Gensini score was the sum of all stenotic coronary site scores (scores of different parts × scores of artery stenosis severity).
Selection and analysis of EMMPRIN and MMP-9 gene polymorphisms
Sequences of all single nucleotide polymorphisms (SNPs) were obtained from National Center of Biotechnology Information (NCBI,
Biochemical indicator test
The serum levels of total cholesterol, glucose, triglyceride, high-density lipoprotein cholesterol (HDL-C), low-density lipoprotein cholesterol (LDL-C), troponin, and lipoprotein A were measured by using standard enzymatic methods. The concentrations of apolipoprotein AI and apolipoprotein B were measured by immune turbidimetry kits (Beijing Leadman Biochemical Company, Beijing, China). The blood pressure of each participant was measured in the hospital. In addition, carotid artery ultrasound and left ventricular ejection fractions were measured by B-mode ultrasonic apparatus (GE Healthcare, Little Chalfont, Buckinghamshire, United Kingdom).
Statistical analysis
All data were analyzed by SPSS 19.0 software (IBM Corp., Armonk, NY). Hardy–Weinberg equilibrium (HWE) for all SNPs was evaluated by goodness-of-fit χ2-test. Enumeration data that obeyed normal distribution were expressed as means ± standard deviation, while those not obeying were expressed as median (interquartile range). Categorical variables were expressed as frequency and constituent ratio. One-way analysis of variance and least significant difference tests were performed to compare the difference of enumeration data that were normally distributed, while the Mann–Whitney U tests were performed in non-normal distributed counting variables. The associations between all SNPs and CHD were assessed by binary logistic regression analysis. The unadjusted odds ratios (ORs) and corresponding 95% confidence intervals (95% CIs) were brought out by goodness-of-fit χ2-test. The adjusted ORs and 95% CIs were calculated in the regression model. Crossover analysis was used to evaluate the multiplicative interaction effect of gene polymorphisms and environmental factors (smoking, hypertension, and diabetes), gene polymorphisms only and environmental factors only on CHD. p < 0.05 was considered to be statistically significant.
Results
Basic information of the study population
The distribution of total six genotypes and allele frequencies is represented in Table 1. The sequences of the primers used in this study are listed in Supplementary Table S1. Except for MMP-9 SNP rs3918242 and EMMPRIN rs8259, the rest of candidate SNPs were in accordance with genetic balance indicated by HWE test in whole study population, while all six SNPs were in line with HWE test in non-CHD control group.
Basic Information of All Six Single Nucleotide Polymorphisms
CHD, coronary heart disease; EMMPRIN, extracellular matrix metalloproteinase inducer; HWE-p-value (non-CHD), Hardy–Weinberg equilibrium in non-CHD control population; HWE-p-value (total), Hardy–Weinberg equilibrium in whole study population; MAF (non-CHD), minor allele frequency in non-CHD control population; MAF (total), minor allele frequency in whole study population; MMP, matrix metalloproteinase; SNP, single nucleotide polymorphism; UTR, untranslated region.
Total 812 CHD patients including 594 men and 218 women and 258 non-CHD patients containing 120 men and 138 women were definitely diagnosed by coronary angiography. The baseline characteristics of all participants are listed in Table 2. The distribution of alcohol drinking, BMI, triglyceride, HDL-C, LDL-C, and apolipoprotein B had no significant differences between CHD and non-CHD patients. However, CHD patients were more likely to have a history of smoking, diabetes, and hypertension, with higher levels of blood glucose, apolipoprotein AI, lipoprotein a, troponin compared with control group (p < 0.01).
Distribution of Basic Information Among Patients with Coronary Heart Disease and Noncoronary Heart Disease Groups
Case group: patients diagnosed as CHD. Control group: patients diagnosed as non-CHD.
p < 0.05; ** p < 0.01.
BMI, body mass index; HDL-C, high-density lipoprotein cholesterol; LDL-C, low-density lipoprotein cholesterol; LVEF, left ventricular ejection fraction.
Associations of EMMPRIN and MMP-9 gene polymorphisms with the presence of CHD
No significant differences of all SNPs were observed in the CHD group compared with non-CHD group except for EMMPRIN gene polymorphism (rs8259) and MMP-9 polymorphism (rs3918242). We found a significant correlation between all these mutations and CHD, and after adjusting for confounding factors such as age, sex, BMI, history of hypertension, smoking, and alcohol drinking, the trend of results was similar. As we discovered, rs8259 heterozygous mutation AT had an adjusted OR of 2.038 (adjusted 95% CI: 1.080–3.847, p = 0.028), while rs8259 homozygous mutation TT had an adjusted OR of 1.499 (adjusted 95% CI: 0.786–2.859, p = 0.219) compared with wild-type rs8259 AA. Consistently, rs3918242 TT increased the risk of suffering CHD (adjusted OR = 2.559, adjusted 95% CI: 1.316–4.975, p = 0.006). However, rs3918242 CT may have reduced the occurrence of CHD (adjusted OR = 0.607, adjusted 95% CI: 0.403–0.916, p = 0.017) (Table 3). The remaining genetic models further confirmed a part of the results we obtained. By recessive model, rs3918242 TT had an adjusted OR of 2.791 (adjusted 95% CI: 1.442–5.403, p = 0.002) compared with rs3918242 AA+AT. By additive model, rs3918242 TT had an adjusted OR of 1.640 (adjusted 95% CI: 1.172–2.295, p = 0.004) compared with rs3918242 AA. Regrettably, these models reflected that there was no association between rs8259 AT and CHD (Table 4). We further explored the association between allele's frequencies and the occurrence rate of CHD. No significant differences were observed in the distribution of allele's frequencies between CHD and non-CHD groups (Table 5).
Association of Selected Single Nucleotide Polymorphisms with the Occurrence of Coronary Heart Disease
Case group: patients diagnosed as CHD. Control group: patients diagnosed as non-CHD. Two samples failed detecting EMMPRIN gene rs6758 polymorphism, while three samples failed detecting EMMPRIN gene rs8637 polymorphism.
ORs (95% CI)a and p-valuesa were adjusted for age, sex, smoking, BMI, alcohol drinking, hypertension, and diabetes.* p < 0.05; ** p < 0.01.
CI, confidence interval; OR, odds ratio.
Different Genetic Models of Selected Single Nucleotide Polymorphisms
Two samples failed detecting EMMPRIN gene rs6758 polymorphism, while three samples failed detecting EMMPRIN gene rs8637 polymorphism.
ORs (95% CI)a and p-valuesa were adjusted for age, sex, smoking, BMI, alcohol drinking, hypertension, and diabetes.
p < 0.01.
Effect of Alleles Frequencies on the Occurrence of Coronary Heart Disease
Case group: patients diagnosed as CHD. Control group: patients diagnosed as non-CHD. Two samples failed detecting EMMPRIN gene rs6758 polymorphism, while three samples failed detecting EMMPRIN gene rs8637 polymorphism.
The combined effects of environmental factor and gene polymorphism rs8259 and rs3918242 on CHD
Moreover, gene–environment interaction and crossover analysis were conducted for rs8259 and rs3918242. As we know, smoking, hypertension, and diabetes were all critical and typical environmental factors for CHD, and the history of these factors was significantly different between control and CHD groups in the current results (Table 2). We investigated collective effects of these environmental factors and gene mutations on the presence of CHD. The results are shown in Tables 6 and 7. We chose nonsmokers with polymorphism rs8259 AA as the reference group, and found that only the smoking had the single influence on CHD risk (OR: 4.444, 95% CI: 1.252–15.783) and no combined effect was observed (p interaction a = 0.774). After adjusting for age, sex, BMI, history of alcohol drinking, hypertension, and diabetes, the results were similar (adjusted OR: 4.071, adjusted 95% CI: 1.033–16.046, adjusted p interaction a = 0.79). We chose nonhypertension patients with rs8259 AA for the reference group, and no single and combined effects were observed between hypertension and genetic mutations of rs8259 whether confounding factors were adjusted. At last we chose nondiabetes patients with rs8259 AA for the reference group, and found that rs8258 AT+TT had the single influence on CHD (OR: 1.926, adjusted 95% CI: 1.029–3.638, p interaction c = 0.371). However, after adjusting for confounding factors except diabetes, there was no significant difference between mutants and wild type (adjusted OR: 1.715, adjusted 95% CI: 0.866–3.399, adjusted p interaction c = 0.762). No combined effect was observed whether confounding factors were adjusted (Table 6).
Gene–Environment Interactions Analysis Between rs8259 and Smoking, Hypertension, and Diabetes on Coronary Heart Disease
Case group: patients diagnosed as CHD. Control group: patients diagnosed as non-CHD. p interaction: the p-value of gene–environment interaction term in the binary logistic regression model.
Adjusted ORs (95% CI)a and adjusted p interaction a were adjusted for age, sex, BMI, alcohol drinking, hypertension, and diabetes.
Adjusted ORs (95% CI)b and adjusted p interaction b were adjusted for age, sex, smoking, BMI, alcohol drinking, and diabetes.
Adjusted ORs (95% CI)c and adjusted p interaction c were adjusted for age, sex, smoking, BMI, alcohol drinking, and hypertension.
p < 0.05; ** p < 0.01.
Gene–Environment Interactions Analysis Between rs3918242 and Smoking, Hypertension and Diabetes on Coronary Heart Disease
Case group: patients diagnosed as CHD. Control group: patients diagnosed as non-CHD. p interaction: the p-value of gene–environment interaction term in the binary logistic regression model.
Adjusted ORs (95% CI)a and adjusted p interaction a were adjusted for age, sex, BMI, alcohol drinking, hypertension, and diabetes.
Adjusted ORs (95% CI)b and adjusted p interaction b were adjusted for age, sex, smoking, BMI, alcohol drinking, and diabetes.
Adjusted ORs (95% CI)c and adjusted p interaction c were adjusted for age, sex, smoking, BMI, alcohol drinking, and hypertension.
p < 0.01.
As for rs3918242, the results were slightly different from rs8259. Taking nonsmokers with rs3918242 CC as the reference group, only smoking had a single influence on CHD risk whether confounding factors were adjusted (adjusted OR: 3.265, adjusted 95% CI: 2.060–5.176, adjusted p interaction a = 0.675). Taking nonhypertension patients with rs3918242 AA as the reference group, only hypertension had the single influence on CHD (OR: 1.734, 95% CI: 1.251–2.405) and no combined effect was observed (p interaction b = 0.732). After adjusting for confounding factors except hypertension, however, no single or combined effects were observed. Taking nondiabetes patients with rs3918242 AA as the reference group, only diabetes had single effect on CHD risk (OR: 2.343, 95% CI: 1.460–3.760, p interaction c = 0.619), and similar results were obtained after adjusting for confounding factors or not (adjusted OR: 2.571, adjusted 95% CI: 1.542–4.284, adjusted p interaction c = 0.447) (Table 7).
rs3918242 CT had a protective effect on CHD in both male and female populations
To further assess the impact of gene polymorphisms rs8259 and rs3918242 on the severity of CHD, the Gensini score system was applied to evaluate according to location and severity of the stenosis. Due to the severe skewed distribution of the data, the scores were represented as median (quarterback spacing) (Fig. 1). The specific data are listed in Supplementary Table S2. We chose rs8259 AA as the control group and found that rs8259 had no significant effect on the severity of the CHD (Fig. 1A). Then, we analyzed the male and female populations separately. Consistently, the significance had no change by analyzing the male and female populations separately (Fig. 1B, C).
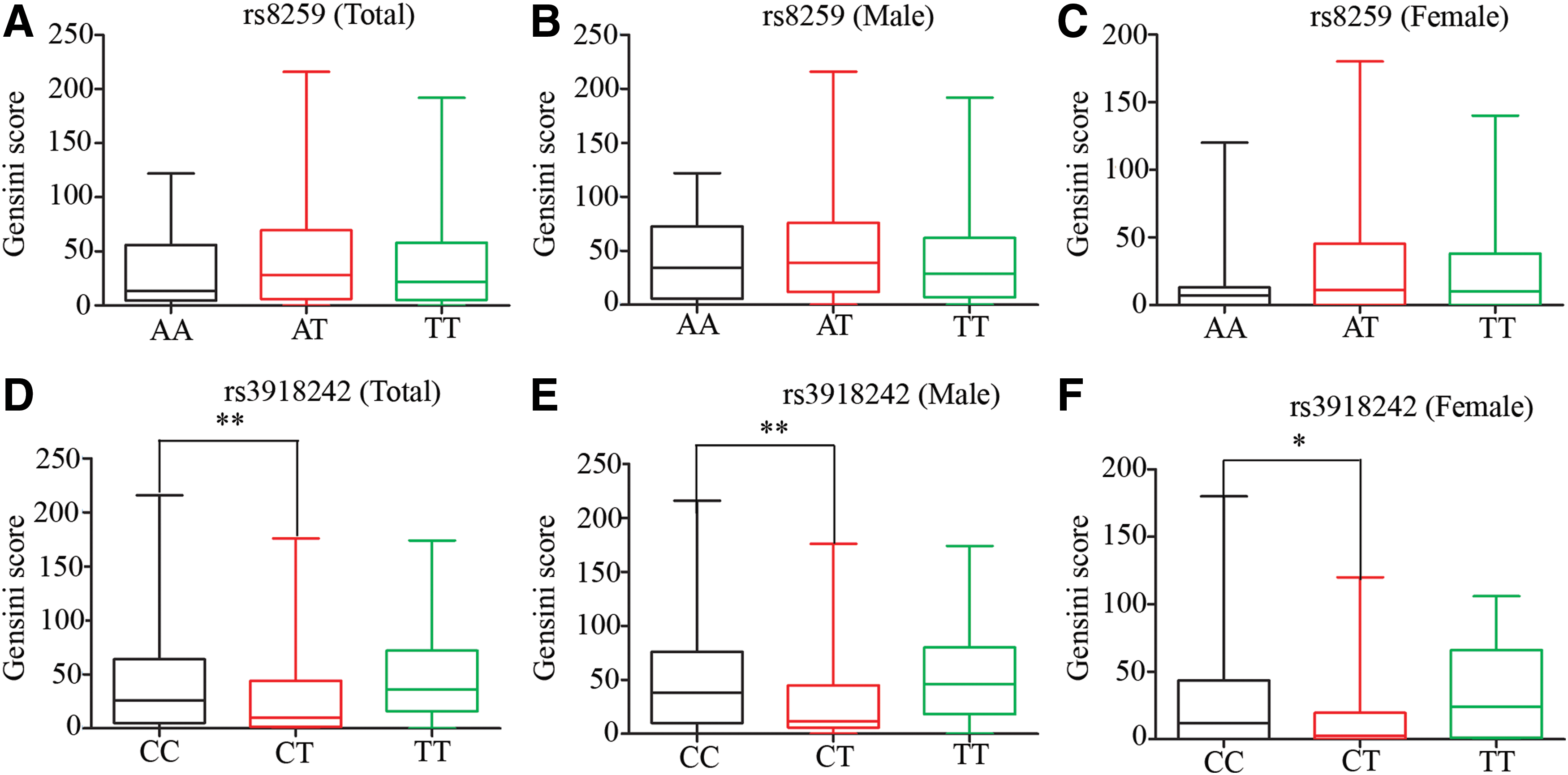
Gensini score evaluating analysis between rs8259, rs3918242, and severity of CHD.
When we chose rs3918242 CC as the control group the results were slightly different. The results exhibited that gene polymorphism rs3918242 CT significantly decreased the severity of CHD (p < 0.0001), while rs3918242 TT had no significant effect (Fig. 1D). Similar results were obtained in both male and female populations separately (Fig. 1E, F).
Discussion
Abundant evidence supported that CHD was a complex pathophysiological process in which an intrinsic gene interacted with the external environment (Zhao et al., 2018; Hu et al., 2019; Zhang et al., 2019). We investigated the potential association of EMMPRIN and MMP-9 gene polymorphisms and CHD susceptibility based on 812 CHD patients and 258 non-CHD controls. The results demonstrated that EMMPRIN gene polymorphism rs8259 and MMP-9 gene polymorphism rs3918242 were significantly related to the morbidity of CHD. Stratified analyses exhibited that rs3918242 CT decreased the severity of CHD in the male and female population. However, gene frequency had nothing to do with the occurrence of CHD. In addition, no gene–environment crossover effect was observed in this study.
Although the mechanisms of EMMPRIN remain obscure, it had been identified as a marker of inflammation for its contribution to inflammatory disease in recent years (Gwinn et al., 2006; Kosugi et al., 2015; Luan et al., 2018). EMMPRIN and cyclophilin A (CyPA) interaction regulated membrane type-1 (MT1)-MMP, MMP-9, and macrophage colony-stimulating factor (M-CSF) during foam cell formation and may promote the vulnerability of atherosclerotic plaques (Seizer et al., 2010). Yuan et al. (2010) found that EMMPRIN–CyPA binding enhanced the inflammatory activity of monocytes/macrophages in atherosclerotic plaque, a cascade of downstream inflammatory responses were inhibited significantly by blocking EMMPRIN in monocytes. Also, disruption of EMMPRIN interaction with CyPA could reduce infarct size and preserve systolic function after myocardial ischemia-reperfusion (Seizer et al., 2011). Highly expressed EMMPRIN in activated platelets, which was demonstrated by its upregulation in the coronary circulation in patients with stable coronary disease (Yong et al., 2013), promoted platelets–monocytes interaction by EMMPRIN homophilic binding to medicate regulation of MMP-9 and inflammatory factors such as interleukin-6 (IL-6) in monocytes through nuclear factor-kappaB (NF-κB) signaling (Schmidt et al., 2008).
In addition to high expression in circulation cells, serum EMMPRIN had been reported to be significantly upregulated in the MI group (Akkus et al., 2016), which indicated that EMMPRIN may have the potential to predict and diagnose ACS. Although one large sample research drew the opposite conclusion (Ueland et al., 2017), these results proved the undeniable connection between CHD and EMMPRIN. In this study, the conclusions we reached about the relationship between EMMPRIN SNPs and MMP-9 SNPs and CHD were not convincing.
To further explore the association between genetic factors and CHD, genetic analyses were conducted by several genetic models. In additive model, only rs3918242 had statistical significance, which suggested that rs3918242 TT significantly improved the risk of CHD (adjusted OR: 1.64, 95% CI: 1.172–2.295, p = 0.004) (Table 4). The results of the recessive model were similar to the additive model and further confirmed the pathogenetic role of rs3918242 TT in CHD (adjusted OR: 2.791, 95% CI: 1.442–5.403, p = 0.002). Contrarily, it was interesting that rs3918242 CT significantly decreased the risk of CHD (adjusted OR: 0.607, 95% CI: 0.403–0.916, p = 0.017*).
Inconsistently, previous studies found that rs3918242 T allele carriers (CT+TT) had significantly increased occurrence of MI compared with the CC carriers (Wang et al., 2011; Rodríguez-Pérez et al., 2016). However, the MMP-9 rs3918242 polymorphism was found to be unrelated to CHD in patients with type 2 diabetes (Klimontov et al., 2017). To solve the inconsistent situation, a meta-analysis (Zhang et al., 2018) was conducted and found a significant association between rs3918242 polymorphism and coronary artery disease (CAD), and MMP-9 T allele may be a risk factor for CAD and MI. In addition, rs3918242 polymorphism tended to be associated with new clinical composite events in CHD population (Opstad et al., 2013).
Although T allele and TT genotypes were confirmed to be risk factors for predicting CHD in previous studies, the association between rs3918242 CT and CHD remained controversial. No evidence was found that rs3918242 CT was associated with the occurrence of CHD in previous research studies (Mahmoodi et al., 2017; Xu et al., 2017). Haberbosch and Gardemann (2005) found that CHD score in the population with apolipoprotein B > 1.27 g/L was lower in rs3918242 CT compared with rs3918242 CC. Ghaderian et al. (2010) found that the proportion of CT carriers was lower in acute myocardial infraction (AMI) patients, and the concentration of MMP-9 was lower in AMI patients carrying CT genotype accordingly. Unfortunately, none of these results were statistically significant. We inferred that the concentration of MMP-9 was lower in CT genotype carriers compared with CC carriers in a certain population. Different characteristics of the study population, including gender composition, lifestyle, the severity of the disease, and underlying disease status, might together contribute to the result and further subgroup analysis was required to confirm our hypothesis.
Limited studies pointed out the function of rs8259 genotype to risk of cardiovascular diseases in Chinese population. Inconsistent with what we obtained in the study, Yan et al. (2015) found that EMMPRIN rs8259 TT genotype and allele T gene may decrease the relative risk of suffering from ACS through downregulating the expression of EMMPRIN. Similarly, Li et al. (2017b) found that rs8259 TT genotype was associated with a decreased risk of chronic heart failure.
The inconsistent results reminded us that the effects of genes on CHD were not single and absolute. Further gene–environment analysis was required. Except for a few diseases caused solely by genes (like congenital genetic disease) or the environment (like traffic accidents), most diseases were the results of a combination of genes and the environment, especially in complicated chronic diseases, in which interaction between gene and environment exerted an important role (Sumi et al., 2017; Zhao et al., 2017b).
Currently, many studies have explored the effects of gene–environment interaction, especially smoking-gene combined impact on CHD. Saleheen et al. (2017) proved that the risk of CHD was reduced in nonsmokers combined with ADAM metallopeptidase with thrombospondin type 1 motif 7 (ADAMTS7) rs7178051 polymorphisms. Chi et al. (2018) found that interaction between phospholipase A2 group VII (PLA2G7) rs1805017 and smoking was associated with increased risk of CHD. As we know, smoking, hypertension, and diabetes were all critical risk factors for CHD, so we focused on these CHD-related lifestyles and risk factors in our research. Although part of single gene or environment effect was observed to be associated with CHD, unfortunately, we found no crossover effect between selected SNPs and environmental factors in our study. The interaction effects and mechanisms needed to be further explored in large sample studies. Perhaps more confounding factors, including diet, race, and others, need to be adjusted to increase the credibility of the results.
To assess the severity of CHD, Gensini score, a currently recognized indicator in CHD research, was applied to assess the severity of CHD in each patient according to the location and extent of coronary stenosis (Zhu et al., 2017; Rampidis et al., 2019). We found that the genetic polymorphism rs8259 did not affect the severity of CHD, and there were no significant statistical differences in the subgroup analysis of gender. In contrast, we found that the genetic polymorphism rs3918242 CT significantly reduced the severity of CHD in both males and females, suggesting that rs3918242 CT may be a protective factor for CHD.
As mentioned before, Haberbosch and Gardemann (2005) found that Gensini score of total studied population whose apolipoprotein B > 1.27 g/L was lower in genotype CT compared with CC although the difference was not statistically significant. Another study based on Chinese population found that the polymorphism of MMP-9 rs3918242 gene was not correlated with the degree of stenosis or the number of coronary arteries, and had no predictive value for the degree of CHD (Li, 2004); however, the simple capacity of the study was limited (<150) and may not reflect the authentic situation. Further studies were required, and different confounding factors, including lifestyle, drug usage, age, gender, and race, should be taken into account.
Our research results were not sufficient to show gender difference in the effects of selected gene sites SNPs on the severity of CHD. However, Savji et al. (2013) investigated that the relationship between age and disease in the male group of peripheral arterial disease and carotid stenosis was more pronounced, and more apparent for women with abdominal aortic aneurysm in a large database of >3.6 million self-referred individuals. Females suffering from ST-elevation MI were associated with a lower mortality in long term (Kanic et al., 2019). In addition to the physical differences, gender difference may be responsible for the differences in lifestyles between male and female in this study (Zhao et al., 2017a).
Further, our experiment still had some limitations. First, subjects in the control group and case group had not been matched strictly. Since all patients required coronary angiography to determine the severity of coronary stenosis, it was difficult to equally and randomly distribute patients in both groups, which may result in partial of SNPs not meeting the Hardy–Weinberg equilibrium test. Second, due to compliance with doctor's advice, the lifestyle of some patients might have changed, which could not be reflected in our research, further molecular-level research associated with selected SNPs should be conducted in the near future. Basic information of all patients is listed in Supplementary Table S3.
In conclusion, we confirmed that EMMPRIN gene polymorphism rs8259 and MMP-9 polymorphism rs3918242 were significantly associated with the risk of CHD. Gensini score suggested that the rs3918242 CT reduced the severity of coronary stenosis. The findings may provide a novel biomarker for the risk prediction, prevention, and early diagnosis of CHD. Pathological mechanisms related research studies were needed to be carried out further to disclose the role of EMMPRIN and MMP-9 in the development and progression of cardiovascular disease.
Authors' Contributions
Y.W. and T.C. performed statistical analysis and wrote the article. J.R., D.L., and X.L. obtained venous blood samples. S.L., C.X., and J.L. were responsible for data entry. X.C. and L.T. led the experiments of this study and provided technological assistance.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants obtained from the National Natural Science Foundation of China (Grant No. 81770475) and the Major Research and Development Project for the Zhejiang Science and Technology Agency (Grant No. 2017C03034).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.