
Abstract
Fibroblast growth factor 21 (FGF21) is a hormone-like member of the FGF family that is associated with cell death in atherosclerosis. However, its underlying mechanisms remain unclear. In this study, the effect of FGF21 on endothelial cell pyroptosis and its potential mechanisms were investigated. Results showed that FGF21 inhibits oxidized low-density lipoprotein (ox-LDL)-induced pyroptosis and related molecular expression in human umbilical vein endothelial cells (HUVECs). Mitochondrial function was damaged by ox-LDL and restored by FGF21. A mechanism proved that ubiquinol cytochrome c reductase core protein I (UQCRC1) was downregulated by ox-LDL and upregulated by FGF21. Further, the silencing of UQCRC1 aggravated HUVEC pyroptosis and impaired mitochondrial function and reactive oxygen species (ROS) production. Moreover, Tet methylcytosine dioxygenase (TET2) was involved in the regulation of UQCRC1 expression and pyroptosis. In summary, FGF21 inhibited ox-LDL-induced HUVEC pyroptosis through the TET2–UQCRC1–ROS pathway.
Introduction
Fibroblast growth factor 21 (FGF21), which belongs to the FGF endocrine subfamily (Kuro-o, 2008; Itoh, 2010), is involved in energy metabolism in many organs (Fisher and Maratos-Flier, 2016). FGF21 is an effective factor that regulates glucolipid metabolism and prevents cardiovascular disease (CVD) development, especially atherosclerosis (AS). In addition, AS is a main cause for CVD (Frohlich and Al-Sarraf, 2013), which is also a chronic inflammatory disease associated with endothelial dysfunction/damage, proinflammatory cytokine release, and foam cell formation (Galkina and Ley, 2009; Back and Hansson, 2015; Tanaka et al., 2016).
Pyroptosis is a programmed cell death that is characterized by plasma membrane rupture and proinflammatory mediator release (Man et al., 2017; Hoseini et al., 2018). Pyroptosis is related to endothelial cell dysfunction and AS development (Li et al., 2018; Wu et al., 2018).
Reactive oxygen species (ROS) can mediate the initiation of pyroptosis in oxidized low-density lipoprotein (ox-LDL)-treated human umbilical vein endothelial cells (HUVECs) (Jiang et al., 2018; Zhaolin et al., 2019b). FGF21 increases the levels of anti-oxidative factors, such as superoxide dismutase (SOD) and glutathione (GSH), to protect against the effect of oxidative stress and ROS (Jiang et al., 2014). In this regard, FGF21 may potentially inhibit pyroptosis.
ROS is mainly produced by mitochondria, ubiquinol cytochrome c reductase core protein I (UQCRC1), a subunit of complex III in the respiratory chain of mitochondria. UQCRC1 knockout has been shown to worsen the outcome in some ischemia/hypoxia diseases (Ellinger et al., 2016). However, the mechanism of expression regulation remains unclear, especially its epigenetic regulation. Could UQCRC1 be regulated by methylation modification? Tet methylcytosine dioxygenase 2 (TET2) is an epigenetic regulatory enzyme that catalyses the oxidation of 5-methylcytosine (5mc) in DNA into 5-hydroxymethylcytosine (5hmc) and also exerts noncatalytic actions (Ko et al., 2010). The abnormal methylation of DNA is related to cell death (Kontaki and Talianidis, 2010) and AS (Hai and Zuo, 2016).
We hypothesize that FGF21 can inhibit ox-LDL-induced endothelial cell pyroptosis by the TET2–UQCRC1–ROS pathway and tested our hypothesis by employing vascular endothelial cells and studies regarding the role as played by TET2, UQCRC1, and ROS.
Materials and Methods
Cell culture
HUVECs were obtained from Science Cell Research Laboratories (Carlsbad, CA) and cultured in endothelial cell medium containing 10% FBS in a humidified atmosphere at 37°C, 5% CO2, and 95% air. First, the cells were passaged three times up to confluence and allocated into three groups with an equal number. The groups were treated with the control (endothelial cell medium). After treatment, the cells were harvested and extracted for analysis. Overexpression/short hairpin RNA lentivirus of TET2 and negative control were synthesized by GeneChem (Shanghai, China).
Small interfering transfection
Small interfering RNA (siRNA) was synthesized by GeneChem (Shanghai, China). HUVECs were transfected with UQCRC1siRNA in accordance with the manufacturer's instructions. Transfection efficiency was evaluated with protein expression by Western blot analysis.
ROS detection
Based on the oxidative conversion of cell-permeable 2′,7′-dichlorofluorescein diacetate (DCFH-DA; KeyGen, Jiangsu, China) into fluorescent dichlorofluorescein, the level of ROS produced by HUVECs was tested by using DCFH-DA reaction with hydroxyl radical, hydrogen peroxide, or peroxynitrite. The different groups of cells were incubated with endothelial cell medium. After different treatment, the cells were washed twice with cold phosphate-buffered saline (PBS; pH 7.4) and incubated with DCFH-DA at room temperature for 30 min in the dark. Dihydroethidine was oxidized in the cytoplasm and entered the DNA of the cell, giving the nucleus a bright red fluorescence, which could be an indicator of peroxide. The fluorescent signal was recorded by using fluorescence microscopy (Olympus Microscope IX3; Olympus, Tokyo, Japan).
Western blot analysis
Cells were washed three times with PBS and lysed with radioimmuno-precipitation assay and phenylmethanesulfonyl fluoride (94:6) on ice for 30 min. The cells were then centrifuged at 9500 g for 15 min and centrifuged again at 12,000 rpm/min for 30 min at 4°C. The supernatant was assayed for protein by using a BCA kit. A total of 20 μg of protein was resolved on a sodium dodecyl sulfate polyacrylamide gel and transferred onto a nitrocellulose filter membrane, which was blocked in Tris-buffered saline with Tween® 20 (TBST) with 5% BSA (CWBio, Peking, China). The membrane was incubated with caspase-1 (1:500; Proteintech, Rosemont), NLRP3 (1:500; ABclonal, MA), IL-1β (1:500; Proteintech), TET2 (1:1000; Abcam, MA), and UQCRC1 (1:4000; Proteintech) and diluted in TBST containing 2.5% skim milk buffer at 4°C overnight. After the membranes were washed five times in PBS with Tween 20, they were incubated with fluorescence-conjugated antirabbit IgG secondary antibody at room temperature for 1.5 h (1:2000). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was selected as an internal control. Protein bands were visualized by using Quantity One Software, and target protein was quantified.
Quantitative PCR
Total RNA was extracted by using Trizol (Roche) and reverse transcribed into cDNA by using RTkit (TOYOBO). Quantitative PCR with SYBR Green Premix Dimer Eraser (Takara) was performed by using an ABI 7000sequence detection system, and mRNA levels were normalized according to the levels of the housekeeping gene GAPDH. Primer sequences for quantitative PCR amplification were as follows: IL-1β forward, 5′-CGATCACTGAACTGCACGCT-3′; IL-1βreverse,5′-AGAACACCACTTGTTGCTCCA-3′; UQCRC1 forward, 5′-AATGTCAGGAA GCTGTCTCGT-3′; UQCRC1 reverse, 5′-TATGTCCATGGGATGCCACC-3′; actin forward, 5′-AAGATCAAGATCATTGCTCCTCCTG-3′; actin reverse, 5′-GCCGGA CTCGTCA TACTCC T-3′.
Cellular immunofluorescence
Immunofluorescence was used to detect the expression of caspase-1 and UQCRC1 in HUVECs. The cells were fixed with 4% paraformaldehyde for 30 min, penetrated with 0.1% Triton X-100 for 30 min, and blocked with goat serum for 1 h. The cells were incubated with anti-caspase-1 (1:250; Abcam) and anti-UQCRC1 (1:100; Proteintech) antibodies at 4°C overnight followed by Alexa fluoroconjugated secondary antibody (Invitrogen) in the dark for 1 h; 4′,6-diamidino-2-phenylindole (Beyotime, Shanghai, China) was used to stain nuclei for 5 min. The cells were observed under a fluorescence microscope (Olympus Microscope IX3; Olympus).
Mitochondrial membrane potential detection
Transmembrane potential (Δψ) collapse occurred during mitochondrial dysfunction, whereas JC-1 (5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethyl benzimidazolcarbocyanine) entered the mitochondria. Flow cytometry was used to detect the intensity of green/red fluorescence, which reflects the mitochondrial membrane potential or mitochondrial function. The cells were washed twice with PBS and centrifuged at 2000 rpm for 5 min. The cells were then suspended in 500 μL of JC-1 solution and incubated at 37°C in 5% CO2 for 15 min. The cells were then centrifuged at 2000 rpm at room temperature for 5 min, harvested, and resuspended in 500 μL of 1 × incubation buffer. Finally, the sample was detected with flow cytometry (E x = 488 nm; E m = 530 nm).
ATP production measurement
Hence, ATP was calculated by colorimetry with phosphomolybdic acid. Intracellular ATP level was measured by using an ATP measurement kit (NanJing Jcbio, Co., Ltd., Nanjing, China). The cells (1.0 × 106 cells/well) were washed twice with PBS, lysed with 150 μL of lysis buffer, and centrifuged at 2000 rpm for 5 min to produce insoluble pellets. ATP measurement solution was added in accordance with the manufacturer's protocol. Detection was conducted by using a Microplate Reader (iMark™; America). ATP concentration was calculated based on a standard curve.
qRT-PCR and changes in mRNA expression validation
Trizol reagent was used to isolate total RNA from cells according to manufacturers' instructions. Subsequently, 500 ng of total RNA was used to obtain cDNA by reverse transcription in a final volume of 21 μL, which contained buffer, random hexamers, DTT, dNTPs, and Superscript III First-Strand Synthesis System (Invitrogen). The quantitative expression of mitochondrial genes was done by RNA-Seq after qRT-PCR with SYBR as a fluorescent reporter. Reactions were carried out in well optical plates containing a final volume of 10 μL consisting of TB Green Premix Ex Taq II, Primer F, Primer R, ROX Reference Dye (50 × ), DNA sample, and ddH2O (Takara, Code No. RR820A). GAPDH-specific primers were used to normalize the quantity of applied RNA. ABI Vii7 Sequence detection system (PE Applied Biosystems, Foster City, CA) was used to measure transcript levels. The 2−ΔΔCT method was used to determine the relative changes in gene expression. The abundance of specific mRNAs in samples from the ox-LDL-treated cells was compared with that from the control-treated cells as fold changes in the transcription.
Bioinformatics analysis
The following websites were used for CpG island analysis: MethPrimer and UCSC.
Statistical analysis
Data are shown as mean ± SD. Group differences were analyzed by using one-way ANOVA and Newman–Keuls multiple-comparisons test (GraphPad Prism version5.0, CA). The two groups were compared by two-tailed Student's t-test; values at p < 0.05 were considered statistically significant.
Results
FGF21 curbs ox-LDL-induced HUVEC pyroptosis
ox-LDL is an important atherogenic factor (Zeibig et al., 2019). Our previous study reported that ox-LDL induced HUVEC pyroptosis in a concentration-dependent manner (Zhaolin et al., 2019). In this study, HUVECs were divided into three groups and incubated with cell medium, ox-LDL (100 μg/mL) or ox-LDL (100 μg/mL)+FGF21 (200 ng/mL) for 24 h to elucidate the relationship between FGF21 and pyroptosis. The expression levels of pyroptosis-related molecular caspase-1, NLRP3, and proinflammatory cytokines (IL-1β) increased (Fig. 1A–C) in the ox-LDL group. This phenomenon was reversed by FGF21. The immunofluorescence results showed that caspase-1 expression increased with ox-LDL treatment, and the pyroptosis was reversed by FGF21 (Fig. 1D). Hence, FGF21 inhibited ox-LDL-induced HUVEC pyroptosis.
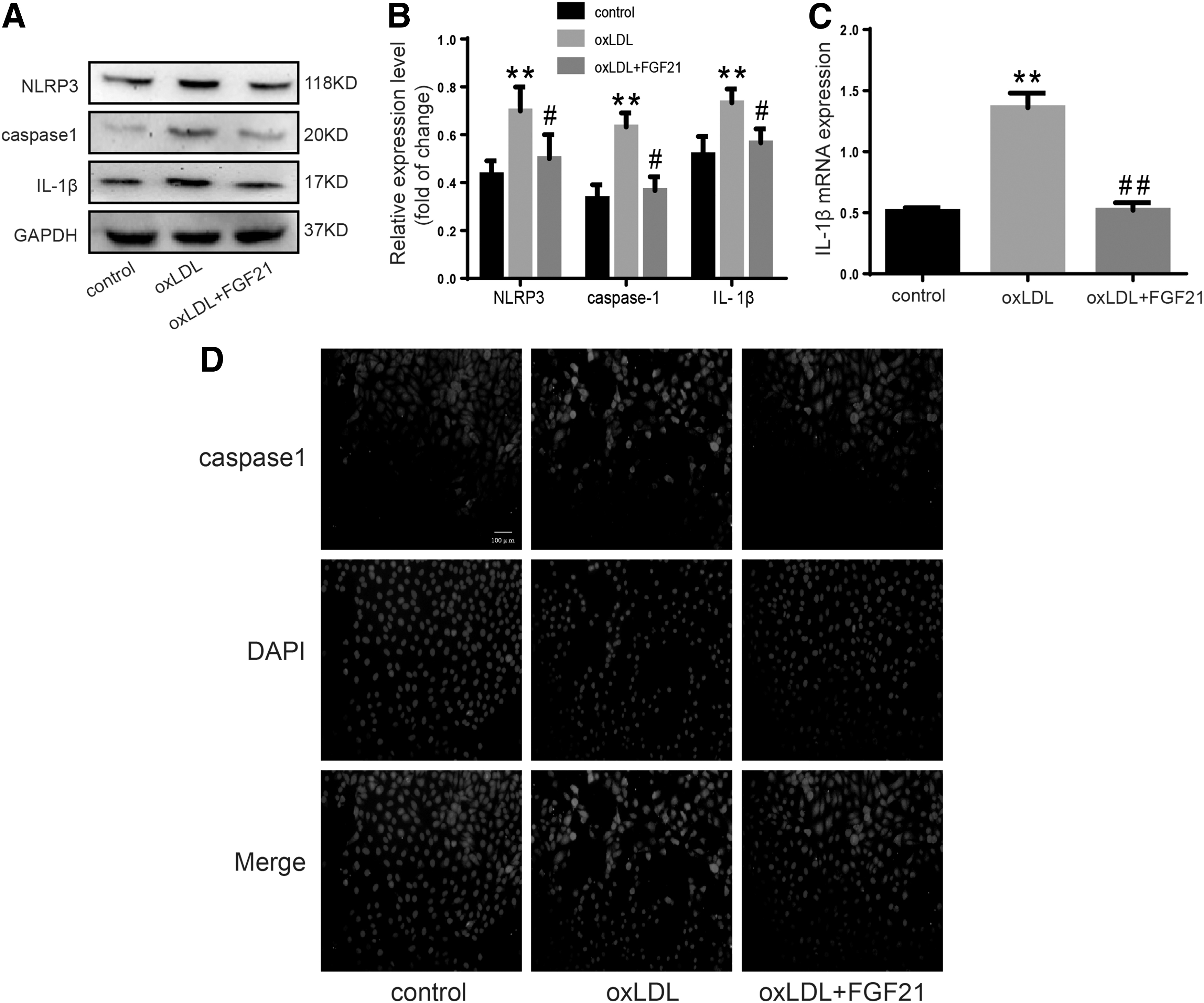
FGF21 curbs HUVEC pyroptosis induced by ox-LDL.
ox-LDL damages mitochondrial function and increases ROS production
Mitochondrial dysfunction exists in many diseases, cell development (Hughes et al., 2019; Perez Ortiz and Swerdlow, 2019), and cell death (Zhaolin et al., 2019). In this work, after treatment of HUVECs with ox-LDL with or without FGF21, mitochondrial membrane potential abnormalities occurred due to decreased ATP production and increased ROS level was detected with DCFH-DA as described in the ox-LDL treatment group; this phenomenon was prevented by FGF21 (Fig. 2A–C).
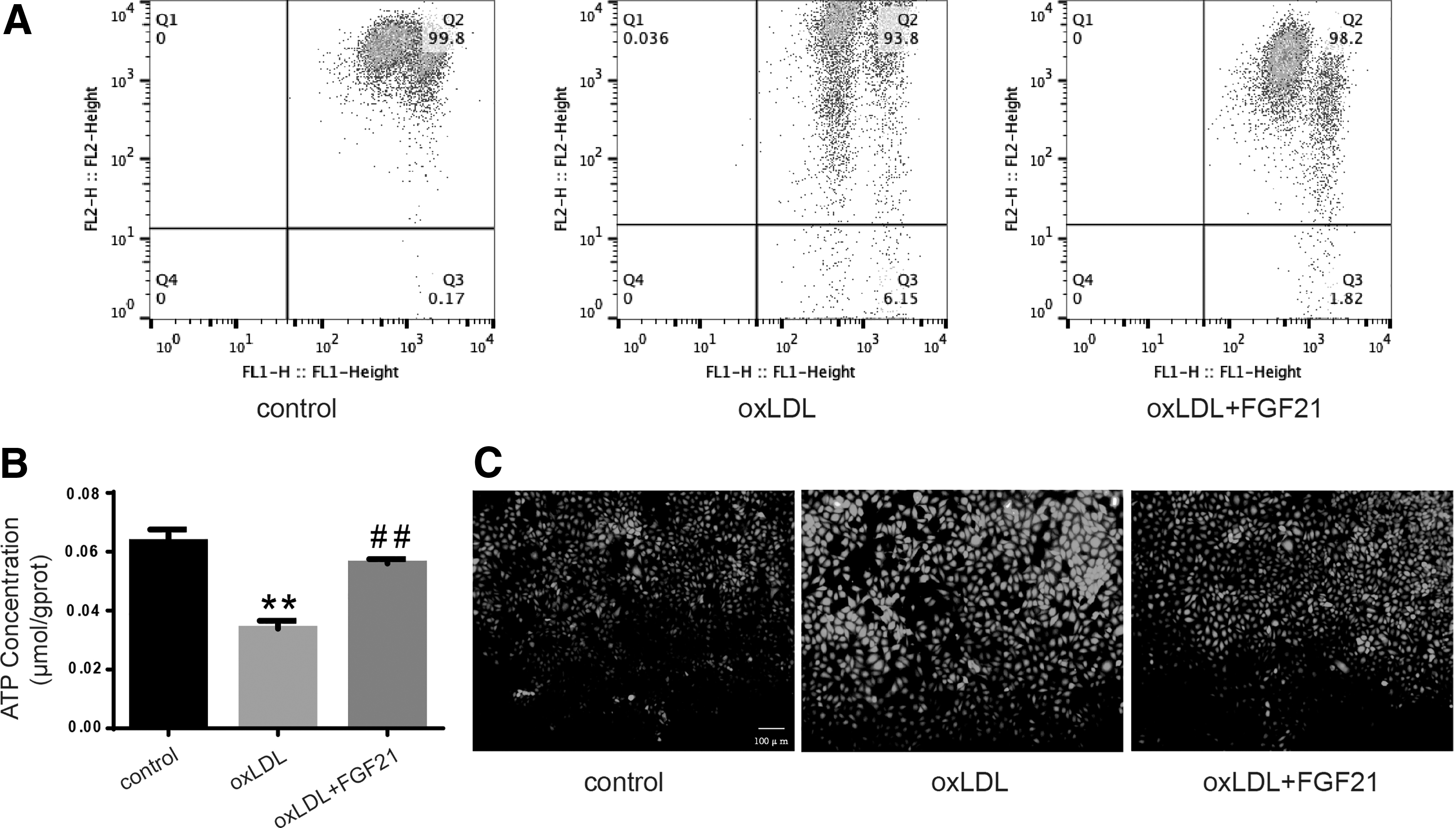
Oxidized low-density lipoprotein damages mitochondrial function and increases ROS production but can be reversed by FGF21.
FGF21 promotes UQCRC1 expression in HUVECs
After treatment of HUVEC with ox-LDL, a macroassay was performed by employing the mitochondria to screen relative change in mitochondrial mRNA expression under ox-LDL. Change in UQCRC1, a subunit of complex III in the respiratory chain of mitochondria, was significant (Fig. 3A). ROS is mainly produced by the mitochondria and released from the mitochondrial respiratory chain. The expression of UQCRC1 decreased during ox-LDL treatment but was restored by FGF21 at the protein and mRNA levels (Fig. 3B, C). The same tendency was observed with the cellular immunofluorescence dye (Fig. 3D).

FGF21 promotes UQCRC1 expression in HUVECs.
UQCRC1 deficiency promotes pyroptosis
To observe the relationship between UQCRC1 and pyroptosis, we knocked down UQCRC1 with UQCRC1siRNA and determined the efficiency (Fig. 4A, B). NLRP3, caspase1, and IL-1β were upregulated; whereas TET2 was downregulated after UQCRC1 was diminished (Fig. 4C, D). These results demonstrated the relationship between UQCRC1 and pyroptosis.
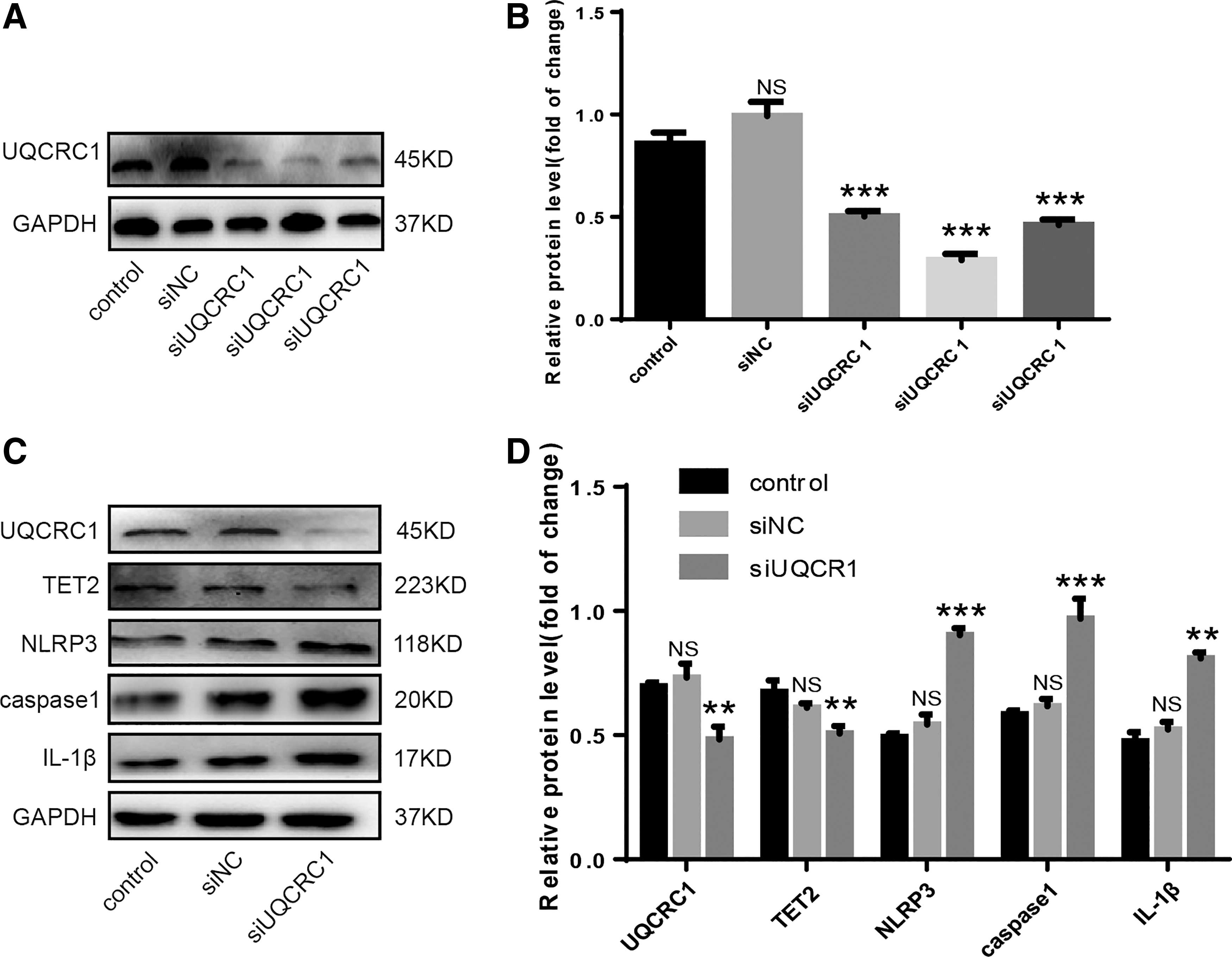
UQCRC1 deficiency promotes pyroptosis.
UQCRC1 is involved with FGF21 mediating mitochondrial function maintenance and pyroptosis
The role of UQCRC1 in ox-LDL-induced HUVECS pyroptosis and the mechanism of the antagonistic effect of FGF21 were determined. The ATP level decreased after UQCRC1 knockdown (Fig. 5A), the ROS production increased (Fig. 5B), the mitochondrial membrane potential decreased (Fig. 5C), and the levels of NLRP3, caspase1, and IL-1β increased (Fig. 5D, E). However, these findings were reversed by FGF21. Hence, UQCRC1 played an important role in the mitochondrial function maintenance and for the pyroptosis antagonistic effect of FGF21.
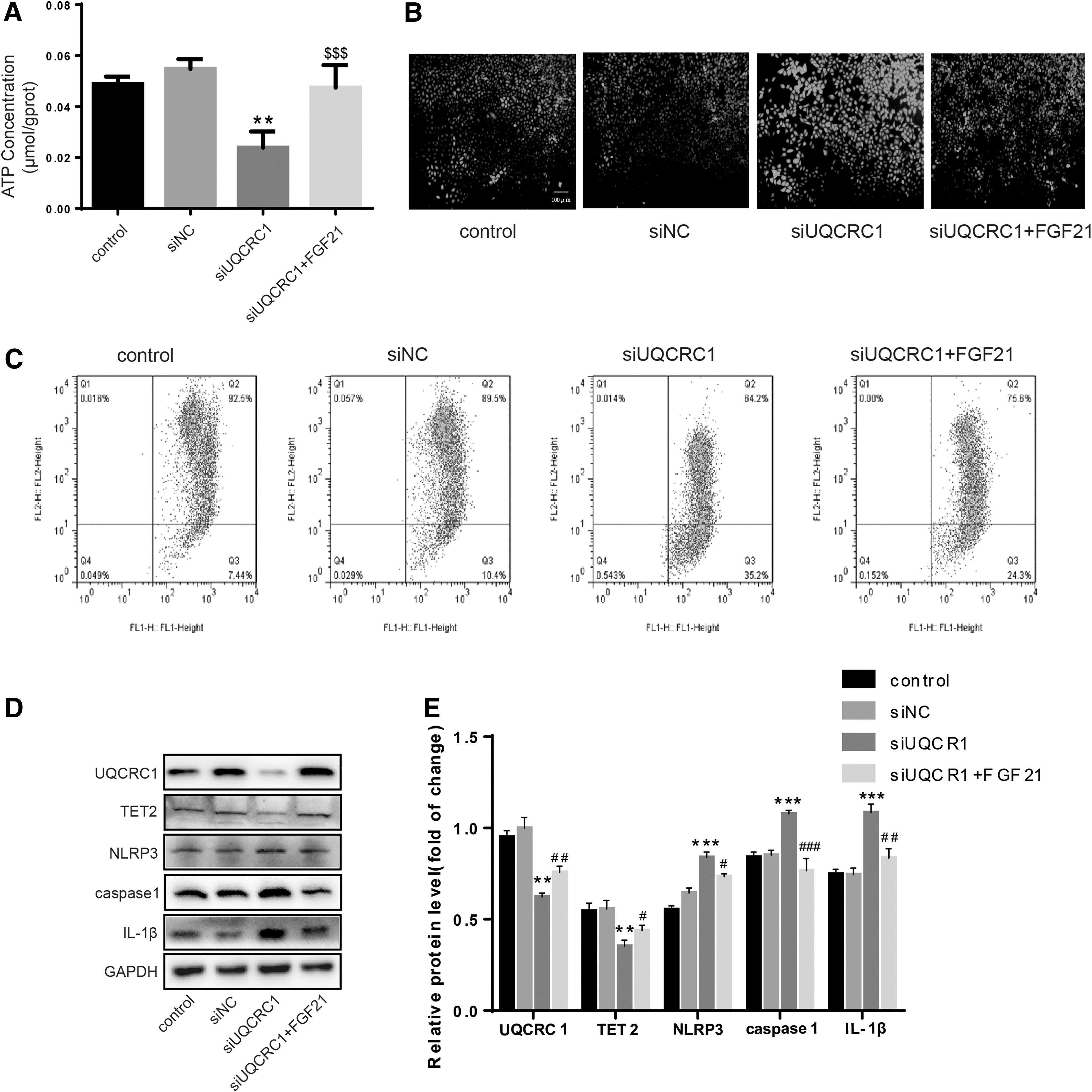
UQCRC1 is involved with FGF21 mediating mitochondrial function maintain and pyroptosis regulation.
TET2 is involved with FGF21 in the upregulation of UQCRC1 expression
TET2 plays an important role in DNA methylation and AS alleviation (Peng et al., 2016; Jiang et al., 2019). Our previous study showed that ox-LDL decreased the expression of TET2 (Zhaolin et al., 2019). A CpG island was found in the UQCUC1 gene promoter by bioinformatics analysis (Fig. 6A). Therefore, we hypothesized that FGF21 mediates UQCRC1 expression, which may be regulated by the methylation modification of TET2. The expression of TET2 was decreased under ox-LDL (Fig. 6B, C). UQCRC1 was downregulated when TET2 expression was inhibited but upregulated when TET2 was overexpressed (Fig. 6D, E). The impaired expression of UQCRC1 by ox-LDL was recovered by FGF21 through TET2.
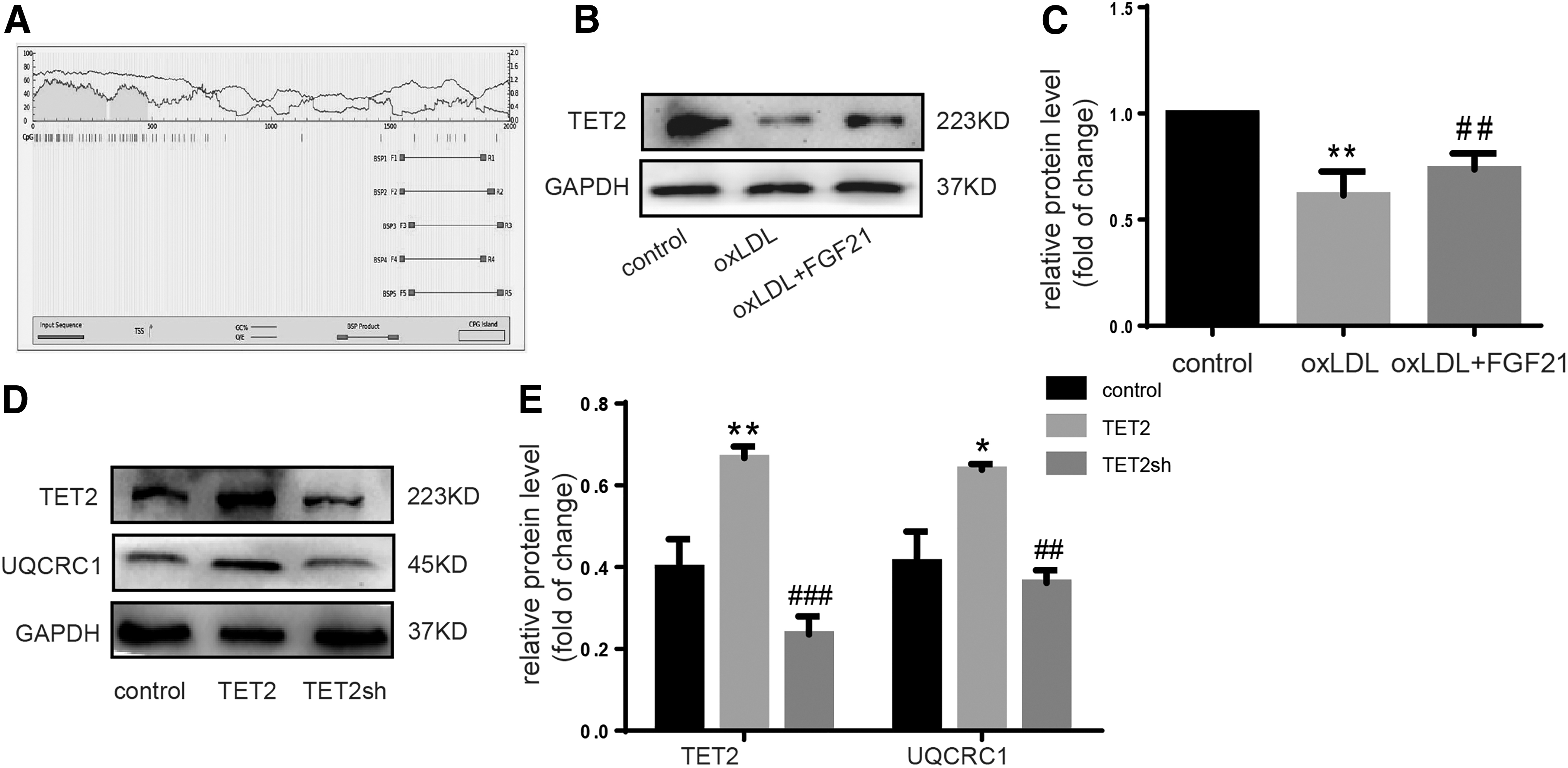
TET2 engages in FGF21's role by upregulating UQCRC1 expression.
Discussion
In this study, we demonstrated that FGF21 can inhibit ox-LDL-induced HUVEC pyroptosis and mitochondrial dysfunction and enhance ROS production through the upregulation of UQCRC1 expression by TET2. We discovered a new mechanism of FGF21 for inhibiting AS development.
AS is a complicated chronic CVD accompanied by cell death and inflammatory cytokine release (Welsh et al., 2017). Pyroptosis is a form of programmed cell death that depends on NLRP3 and caspase-1 activation, which induces IL-1 and IL-18 (Man et al., 2017; Hoseini et al., 2018). Lopez-Pastranaet al. (2015) reported that pyroptosis promotes adhesion molecules (intercellular and vascular cell adhesion molecule-1), thereby triggering monocyte adhesion on release. This phenomenon is beneficial for monocyte migration and atherosclerotic lesion progression. Increased pyroptosis in human aortic endothelial cells leads to plaque area growth and proinflammation cytokine release by the ROS-NLRP3 pathway (Wu et al., 2018). Here, we demonstrated that mitochondrial dysfunction, which was caused by the downregulation of UQCRC1 by ox-LDL-induced excessive ROS production, and pyroptosis could be restored by FGF21 in HUVECs.
Various studies have reported on the role of FGF21 in AS development. FGF21 is an important factor in the regulation of energy metabolism, including glucose, and lipid metabolism (Woo et al., 2013; Kharitonenkov and DiMarchi, 2015; Fisher and Maratos-Flier, 2016). FGF21 reduces lipid profile in animal models, atherosclerotic plaque area (Talukdar et al., 2016), serum triglycerides, VLDLC, and LDL-C and increases HDL-C (Lin et al., 2014). However, the exact mechanism remains unclear. Some transcription factors and transporters may be involved (Lin et al., 2014; Shang et al., 2015). FGF21 protects against AS induced by oxidative stress by enhancing the activation of nuclear factor E2-related factor 2/antioxidant responsive element (Wang et al., 2014) and relative downstream genes, such as SOD, heme oxygenase-1, and GSH (Jia et al., 2018). FGF21 also inhibits ROS effect and apoptosis in the presence of pro-apoptosis proteins, such as the MAPK family (Zhu et al., 2014). Here, we demonstrated that FGF21 reduces ROS production by restoring the function of mitochondria, subsequently alleviating pyroptosis, which is a pro-inflammatory cell death process under ox-LDL, which is a pro-atherogenic factor.
Mitochondria are the powerhouse of the cell and are crucial in maintaining cell homeostasis and function, which are related to pyroptosis (Qiu et al., 2019; Wang et al., 2019, 2019). Mitochondrial homeostasis integrity disruption induced by extracellular stimulations causes mtROS production. Excessive ROS leads to upregulated NLRP3 expression, which triggers the activation of caspase-1, maturation of IL-1β and IL-18. Gasdermin D (GSDMD) can be cleaved by caspase and that promotes cell membrane pore formation, which helps in the release of IL-1β and IL-18 (Zhaolin et al., 2019a). NLRP3 can also be activated by oxidative or apoptosis-induced mitochondrial DNA, which is released during autophagy inhibition (Sollberger et al., 2015). Cardiolipin is necessary for NLRP3 activation; which is located in the inner layer of mitochondria and relocates in the outer layer when the mitochondria lose homeostasis (Iyer et al., 2013). We found that FGF21 plays a protective role in mitochondrial function, but its mechanisms remain unknown.
Limited information is known about the biological function of UQCRC1, a member of the respiratory chain complex III. UQCRC1 deficiency in mice led to decreased mitochondrial membrane potential and ATP production but increased ROS production (Shan et al., 2019). However, increased intercellular ROS regulates pyroptosis and the release of proinflammatory cytokines in endothelial cells under ox-LDL (Zhaolin et al., 2019b). Our results demonstrated that FGF21 improves mitochondrial function and reduces ROS production due to the upregulation of UQCRC1.
Demethylase TET2 can catalyze the oxidation of 5mc in DNA into 5hmc (Ko et al., 2010) and inhibit the development of AS (Jiang et al., 2019). The downregulation of TET2 in ApoE−/− mice increased the plaque area and TET2 silencing reduced autophagy and autophagic flux, resulting in EC dysfunction (Peng et al., 2016). TET2 protects against DNA damage-induced apoptosis (Chen et al., 2018). Our previous study showed that TET2 silencing induces pyroptosis by increasing ROS production in HUVECs (Zhaolin et al., 2019b). Our results showed that FGF21 restores TET2 expression under ox-LDL to protect against proptosis. However, the reason that FGF21 would upregulate the expression of TET2 is unclear, and further research is required.
In summary, our results provide the first evidence that FGF21 protects HUVECs from ox-LDL-induced pyroptosis by repairing mitochondrial function, upregulating TET2 and UQCRC1, and reducing ROS production. Therefore, FGF21 can be a new factor in protecting against AS induced by oxidative stress by demethylation and thus increasing relative protein expression. TET2 and UQCRC1 may be new targets for AS intervention.
This study is not without limitations. No specific evidence was provided in support of the role of FGF21 in AS, and the effect of TET2 on demethylation in improving mitochondrial function must be further experimented and tested in vivo. Another limitation of this study is the lack of experimental data that show whether TET2 effect on mitochondrial function and pyroptosis is achieved by specific methylation genes. We did not test the demethylation effect of FGF21 on other mitochondrial factors. Further studies are needed to establish the concrete molecular regulation mechanism of FGF21.
Conclusion
In this study, we revealed that FGF21 alleviates pyroptosis induced by mitochondrial dysfunction and ROS production by ox-LDL. Further, we also demonstrated that the TET2–UQCRC1 axis plays a key role in mitochondrial function and ROS production. Therefore, FGF21 inhibits pyroptosis induced by ox-LDL, and this phenomenon could further be exploited as a new therapeutic approach for treatment of AS development.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Natural Science Foundation of China (No. 81970389, No. 81070221, No. 81600342, and No. 81770460) and the Innovative Research Team for Science and Technology in Higher Educational Institutions of Hunan Province and the Construct Program of the Key Discipline in Hunan Province (No. 15C1201), the Aid Program (2017KJ268), the Construction Program of Innovation Platform (2017KJ182) from the Science and Technology Bureau of Hengyang City, the third level of Chuanshan Talent project of University of South China (2017CST20), the scientific research foundation for the Excellent Youth of the education department of Hunan province (18B264), and the Innovation project of Huai'an Municipal Science and Technology Bureau (HAB201844).