
Abstract
Pulmonary microvascular endothelium barrier plays a critical role in protecting the pulmonary tissue from inflammatory injury in acute respiratory distress syndrome and acute lung injury (ARDS/ALI). The dysregulation of IQ-GTPase-activating protein 1 (IQGAP1) was an important etiology of endothelium barrier injury. However, significant differentially expressed genes (DEGs) and signaling pathways directly regulated by IQGAP1 are too complicated to fully understand. In this research, we identified a total of 1216 DEGs regulated by knockdown of IQGAP1 in rat pulmonary microvascular endothelial cells on the basis of transcriptomic RNA sequencing (RNA-Seq). Among them, 665 were upregulated DEGs and 551 were downregulated DEGs. Gene ontology analysis has revealed that upregulated DEGs were mainly enriched in DNA replication, cell cycle, and chromosome formation, while downregulated DEGs were mainly involved in the regulation of many cellular bioprocesses including cell proliferation, cell adhesion, and cell migration. Kyoto Encyclopedia of Genes and Genomes pathways analysis toward DEGs showed that upregulated pathways were mainly about DNA replication, while the significantly downregulated pathways were about TNF signaling pathway and some inflammatory- and proliferation-related pathways. Furthermore, we choose 30 DEGs for validation by qRT-PCR, the results were quite consistent with the RNA-Seq. In addition, we also found that knockdown of IQGAP1 caused a significant impact on many cytokines and inflammatory factors, which play a vital role in ARDS/ALI. In summary, in this study on the basis of RNA-Seq, we found IQGAP1 not only exerts a crucial role in microvascular endothelium barrier but also plays an important role in inflammation, which might provide a new insight for future study on IQGAP1 in the related diseases such as ARDS/ALI.
Introduction
Acute respiratory distress syndrome (ARDS) and acute lung injury (ALI) are acute pulmonary diseases that pose great threats to human life. (McNicholas et al., 2018). Approximately, 10% patients, ∼3,000,000 patients are admitted in the intensive care unit (ICU) around the world every year, among which 24% are treated with mechanical ventilation. Although with the widespread use of ventilatory strategies in clinic therapy, the mortality of the ARDS has been declined. The mortality remains from 35% to 46% (Bellani et al., 2016; Fan et al., 2018). The main pathogenesis of ARDS refers to the pulmonary vascular endothelium and epithelium barrier dysfunction, leading to vascular leakage accompanied by uncontrolled release of inflammatory cells and cytokines (Bachofen and Weibel, 1977; Millar et al., 2016).
The pulmonary microvascular endothelium forms the single layer and semipermeable barrier. In comparison to the macrovascular endothelium barrier, pulmonary microvascular endothelium is a more restrictive barrier to protein and blood flux (Parker and Yoshikawa, 2002; King et al., 2004). The integrity of the barrier relies on the tight junction and adhesion junction between endothelial cells, which are served by cytoskeletal microtubes and microfilaments. All these structures and components not only play a vital role in maintaining the integrity and the function of endothelial barrier but also in transmitting signals between cells (Sukriti et al., 2014). As the endothelium barrier injury is the direct factor of vascular leakage, dysfunction of endothelial cells is considered to be one of the most important initiators in the process of uncontrolled inflammatory cascade response (Teijaro et al., 2011). In addition, with the wide use of mechanical ventilation, many complications such as the continuous inflammation and mechanical injury also raised great attention to the endothelium injury by ventilatory strategies. Hence, more and more research emphasized on the endotypes of ARDS, aiming to recover the integrity and function of endothelium barrier and to prevent further injury (Verbrugge et al., 2007). However, the regulatory mechanism in maintaining the homeostasis of microvascular endothelium barrier is far more elusive than we currently know, and much work remains to be done (Müller-Redetzky et al., 2014).
IQ-domain GTPase-activating proteins (IQGAPs) is an evolutionarily conserved family consisting of three members, IQGAP1, IQGAP2, and IQGAP3, and participates in many cellular processes. IQGAP3 was identified in recent years and highly expressed in brain (Wang et al., 2007). Research about IQGAP3 remains elusive and poorly explored (Smith et al., 2015). Although IQGAP1 and IQGAP2 share over 60% similarities in sequence, they vary from function and distribution. IQGAP2 mainly exists in liver and was first described in last two decades (Brill et al., 1996). A decreased expression of IQGAP2 was found in several tumors such as liver (White et al., 2010), gastric (Jin et al., 2008), and prostate cancer (Xie et al., 2012). As the most extensively studied and ubiquitously expressed protein in vivo in comparison with other two members, IQGAP1 has already been known for its scaffolding function (Smith et al., 2015). Beyond functioning as a scaffold protein, IQGAP1 is considered to be an important factor in maintaining the function and integrity of the microvascular endothelium barrier (Mehta et al., 2014; Tian et al., 2016). Decreased expression of IQGAP1 leads to increased endothelium permeability, which is also responsible for the severity of ALI/ARDS (David et al., 2011; Bhattacharya et al., 2012). However, the underlying mechanism and significantly differentially expressed genes (DEGs) in the dysregulated microvascular endothelium barrier induced by IQGAP1 downregulation remain elusive. In this research, we explored the DEGs and their functions, as well as enriched signaling pathways induced by knockdown of IQGAP1 at transcriptional level. This could shed a new light for realizing the role of IQGAP1 in the disease with dysregulated pulmonary microvascular endothelium barrier such as ARDS/ALI.
Materials and Methods
Reagents
Antibody against factor VIII-related antigen and horseradish peroxidase-labeled secondary antibodies were from Boster Biotech (Wuhan, China). Antibody against CD34, IQGAP1, and GAPDH and fluorescent secondary antibody conjugated goat anti-rabbit IgG were purchased from Abcam (Cambridge, MA).
Isolation, identification, and culture of rat pulmonary microvascular endothelial cells
All experimental rats were from Experimental Animal Center of Anhui Medical University. Three Sprague-Dawley male rats weighted 110–130 g were sacrificed by artery bloodletting after intraperitoneal anesthesia with 2% pentobarbital sodium. All rats were maintained and treated strictly in accordance to the guidelines of Animal Center of Anhui Medical University. The animal experimental procedures were permitted by Experimental Animal Ethical Committee of Anhui Medical University (license no. 20190752). Isolation of microvascular endothelia cells was in accordance with the previous method provided by Chen et al. (1995). In brief, the fresh lung was isolated and washed by Dulbecco's modified Eagle's medium (DMEM; HyClone). After removing the pleura, the edges of lung tissue were cut into 1 × 1 × 1 mm and harvest in T25 culture flasks (Corning). Tissue was cultured in DMEM supplemented with 20% fetal bovine serum (Gibco, Australia) and 1% penicillin/streptomycin (Beyotime Biotech, Jiangsu, China) at 37°C incubator containing 5% CO2 for 60 h. Then the tissue was removed and the culture medium was replaced.
Rat pulmonary microvascular endothelial cells (PMVECs) were identified according to previous research (Zhang and Sun, 2005). In brief, cells were cultured in complete medium; when monolayer cell growth reached 80–90% confluency, cells were digested followed by seeding into sterile coverslips, which were coated with poly-L-lysine. Once cell fusion rate got to 70–80%, cells were fixed with 4% paraformaldehyde (PFA) for 15 min at room temperature. For binding to factor VIII-related antigen, cells were permeated using 0.5%Triton X-100 (Solarbio, Beijing, China) for 20 min. The second passage of PMVECs was identified under an inverted microscope (Olympus, Tokyo, Japan) after coating with anti-human factor VIII and CD34 antibody by immunocytochemical staining process (Zhang and Sun, 2005; You et al., 2010). The third passage cells were used for subsequent experiments.
Knockdown of IQGAP1 in PMVECs
PGLV3 (H1/green fluorescence protein [GFP] and Puro vector) lentiviral shRNA targeting IQGAP1 (shIQGAP1) and control lentiviral shRNA (shSc) were obtained from GenePharma (Shanghai, China). Target sequences were as follows: shSc, 5′-TTCTCCGAACGTGTCACGT-3′; and shIQGAP1, 5′-GCAGAATTCTGTGCAGCTTTC-3′.
For constructing IQGAP1 knockdown cells, the third generation of rat PMVECs was seeded into six plates at the density of 5 × 104 per well; when cell growth reached 30–40% confluency, cells were transfected with the lentivirus targeting IQGAP1 (shIQGAP1) or the control (shSc). Transfection was conducted using 5 μg/mL Polybrene according to the protocol provided by GenePharma. Cells transfected with lentivirus were cultured in the complete DMEM at 37°C with 5% CO2 for 8 h; then, cells were washed by phosphate-buffered saline (PBS) and culture medium was replaced. Positive transduction cells were selected using puromycin (Solarbio, Peking, China). To evaluate the efficiency of the transfection, portions of cells expressing GFP were observed using an inverted fluorescence microscope (Olympus). When GFP-positive cell growth reached 90% confluency, cells were collected.
Immunofluorescence
2 × 105 rat PMVECs were seeded on 25 mm diameter coverslips at room temperature. When cell growth reached 70% confluency, the coverslips were washed three times using PBS. After that, cells were fixed with 4% PFA for 20 min and blocked with 5% bovine serine albumin for 1 h. Then cells were incubated with antibody against IQGAP1 (1: 200) at 4°C overnight. Fluorescent staining of cells was conducted on the next day using fluorescent secondary antibody against IgG (1:500) and DAPI (Beyotime Biotech) reagent following the manufacturer's instructions. Cells were observed under inverted fluorescence microscope (Olympus).
Western blot
Total protein of rat PMVECs was harvested using radioimmunoprecipitation assay buffer (RIPA) lysate (Beyotime Biotech). The proteins were separated by 8% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to a polyvinylidene fluoride membrane (Millipore). Then bands were blocked with 5% nonfat milk (BBI Life Sciences, Shanghai, China) followed by incubation with the antibodies against IQGAP1 (1: 1000) or GAPDH (1: 1000) at 4°C overnight. Protein immunostaining was performed using an enhanced chemiluminescence detection kit (Thermo Scientific) according to the manufacturer's instructions.
Total RNA extraction, cDNA library construction, and sequencing
When GFP-positive cell growth reached 90% confluency, cellular total RNA was extracted using TRIzol (Invitrogen, MA) reagent. The integrity of RNA was measured by agarose gel electrophoresis. The quantity and purity of RNA were measured using a NanoDrop ND-1000 (Thermo Fisher Scientific, Wilmington, Delaware) by micro-ultraviolet spectrophotometer sampling assay. One microgram RNA was used to construct a cDNA library using KAPA Stranded RNA sequencing (RNA-Seq) Library Prep Kit (Illumina) according to the manufacturer's instruction. cDNA libraries mixed with different samples were subjected into the sequencing by Illumina HiSeq 4000 (service provided by Kangchen Biotech, Shanghai, China) for 150 cycles.
Bioinformatics analysis
Gene ontology (GO) terms (Maere et al., 2005) involving biological process (BP) terms, cellular component (CC) terms, and molecular function (MF) terms were conducted using a Database for Annotation, Visualization and Integrated Discovery (DAVID 6.8) (Zhong et al., 2019).
Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis was performed using KEGG Orthology-Based Annotation System online tools. The gene network of TNF signaling pathway was also analyzed using online tools. (Pruitt et al., 2009).
qRT-PCR
Cellular total RNA was extracted using TRIzol (Invitrogen) reagent according to the manufacturer's instruction. The first-strand cDNA was constructed by 1 μg RNA using a RevertAid™ First Strand cDNA Synthesis Kit (Fermetas, Waltham, MA).
cDNA was diluted and as the template for qRT-PCR reaction. The volume of reaction system was 20 μL using SYBR® Premix Ex Taq™ II (Tli RNaseH Plus) (Takara, Dalian, China) in accordance with the manufacturer's instructions. The whole process was performed on a QuantStudio™6 and 7 Flex Real-time PCR system (Applied Biosystems). Sequences of target genes' primers were synthesized by Sangon Biotech (Shanghai, China). The data were analyzed by calculating the value of 2−ΔΔCt method on the basis of housekeep gene GAPDH. Each sample was validated thrice and performed in triplicates. Primer sequences are listed in Supplementary Table S1.
Statistical analysis
All data are shown as mean ± standard error. DEGs must meet the criteria of foldchange >1.5 and p-value ≤0.05. Data obtained by qRT-PCR were analyzed using SPSS version 16.0 (IBM, Armonk, NY). Nonpaired student's t-test was used for two-group comparison. There is a statistical difference when p-value ≤0.05.
Results
Identification of rat PMVECs
Given the role of pulmonary microvascular endothelium barrier in ARDS/ALI and advantages of rat models in study of the disease, we choose rat PMVEC lines for research (Guice et al., 1988; Yang et al., 2011). First, we isolated the rat microvascular endothelial cells (PMVECs). Identification of microvascular endothelia cell lines mainly relied on their morphology and biomarkers (Chen et al., 1995). Under normal conditions, the morphology of cultured primary rat PMVECs is polygon or short fusiform under microscopy. At the confluence, these cells showed a typical cobblestone morphology. When subcultured cells grew well and reached confluence, the cells present a vascular-like morphology (Chen et al., 1995). As depicted in Figure 1a and b, the observed isolated cells were in the shape of polygon or short fusiform, which is consistent with previous research. As CD34 and factor VIII-related antigen are biomarkers of microvascular endothelial cells, we also tested the expression of these two biomarkers in cells using immunocytochemical staining (Magee et al., 1994; Muller et al., 2002). In comparison with the control (Fig. 1c), CD34 was shown in brown expressed at cytoplasm and cell membrane (Fig. 1d), while factor VIII-related antigen was stained in brown particles at cytoplasm (Fig. 1e). Taken together, the findings are consistent with previous research (Chen et al., 1995), we isolated and identified rat PMVECs through its morphology and biomarkers.
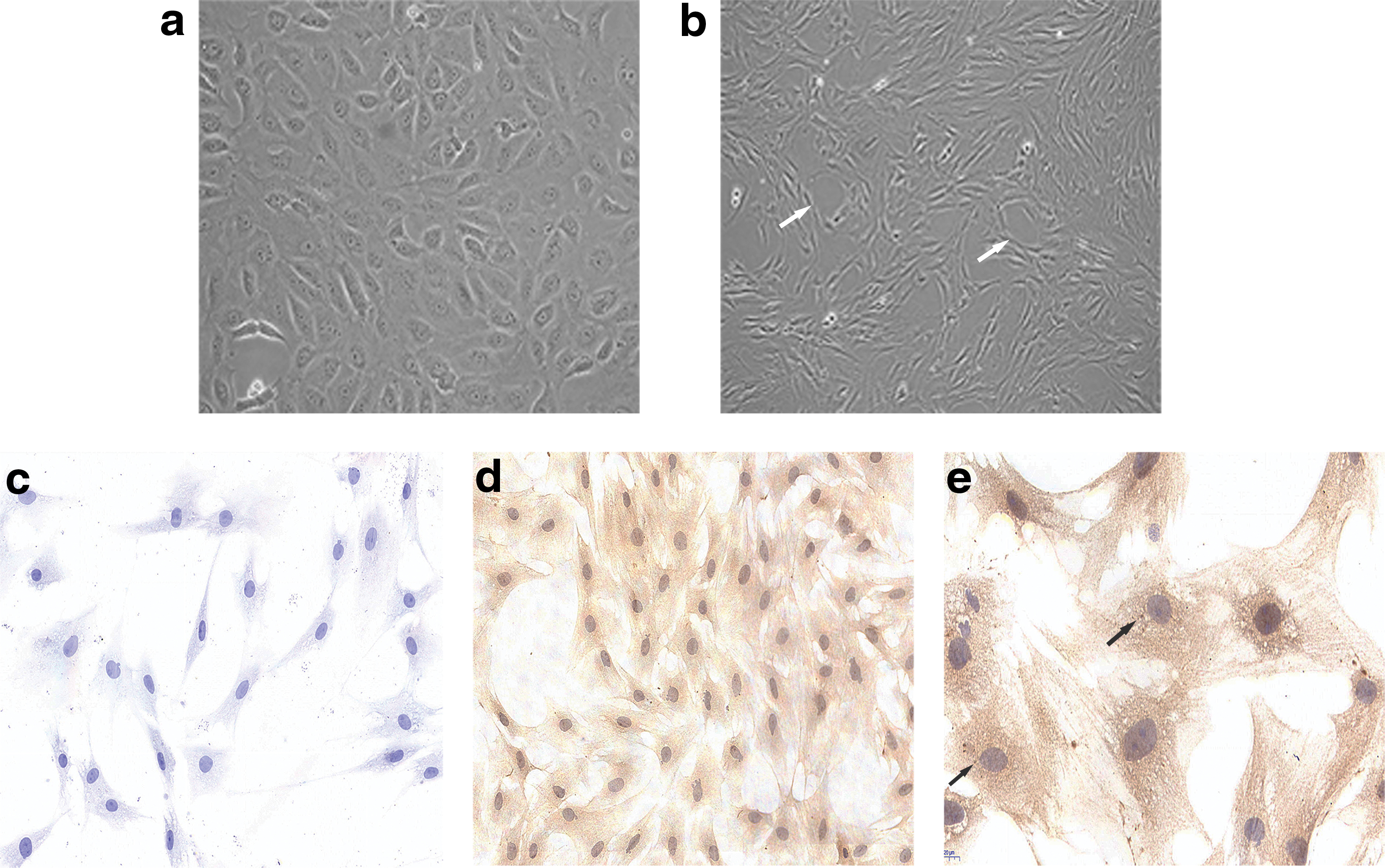
Isolation and identification of rat pulmonary microvascular endothelial cells. Morphology of rat primary (passage 1) PMVECs
Knockdown of IQGAP1
Previous studies have demonstrated that IQGAP1 expressed ubiquitously in cells and located at the sites of cell-cell contact (Kuroda et al., 1998). To explore DEGs and signals induced by the silence of IQGAP1, we knocked down IQGAP1 by transfecting lentivirus shRNA targeting IQGAP1. At first, to evaluate expression of IQGAP1, we observed the cells especially in cell-cell junction using immunofluorescence staining. In consistent with previous research, we found that in cells transfected with lentiviruses expressing empty vector (shSc), location of IQGAP1 is mainly at the cell-cell junction, while in cells with knockdown of IQGAP1, IQGAP1 is scarcely expressed at the cell-cell junction (Fig. 2a). We further validated a significantly downregulation of IQGAP1 in rat PMVECs by qRT-PCR and Western blot, respectively. As depicted in Figure 2b and c, in comparison with control (shSc), cells transfected with shIQGAP1 showed a weaker expression of IQGAP1 at both transcriptional and translational level. Taken together, all these results indicate that IQGAP1 has been successfully knocked down in primary rat PMVECs.
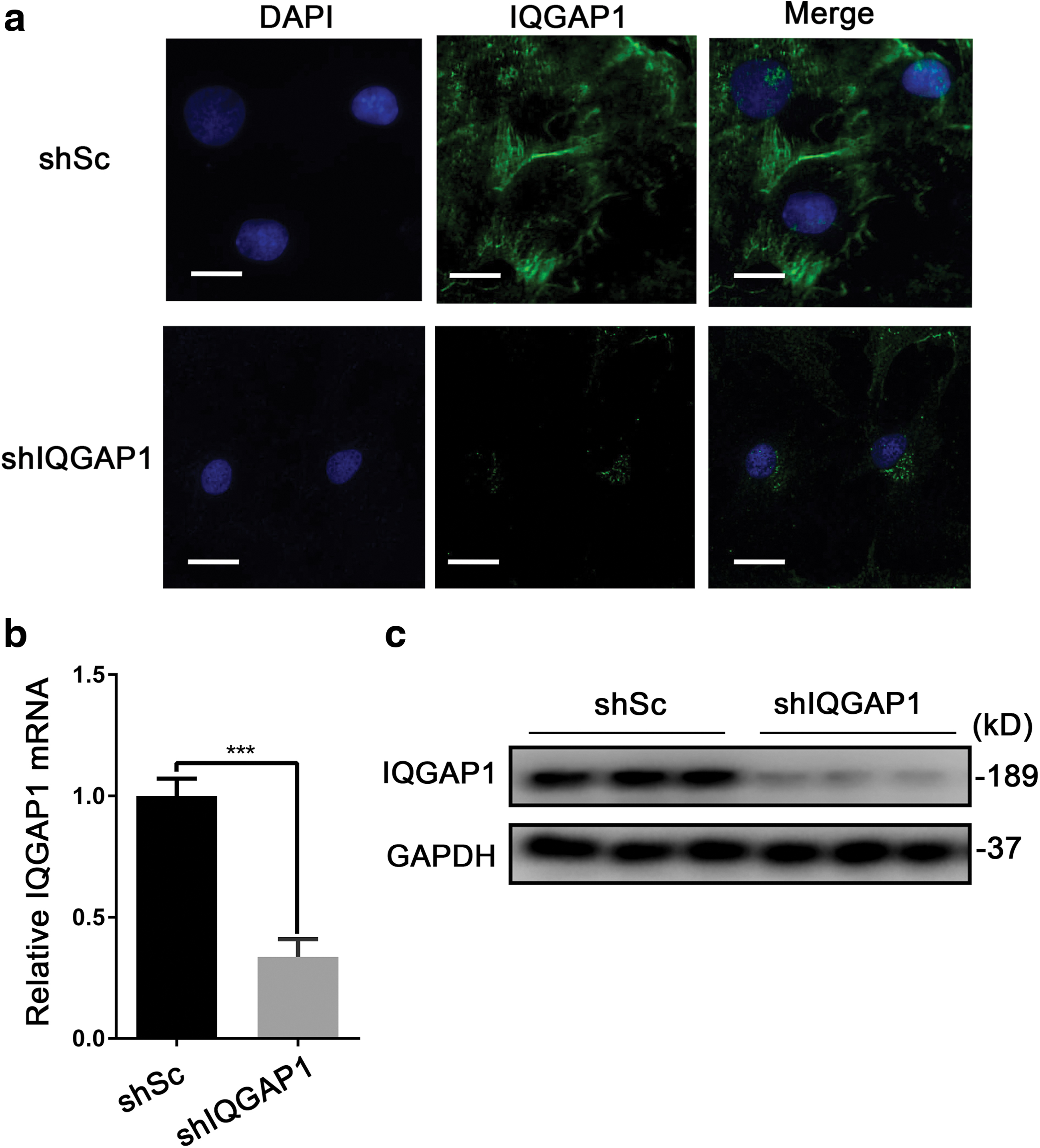
Knocking down IQGAP1 in pulmonary microvascular endothelial cells.
Cluster analysis of DEGs
After the successful construction of IQGAP1 knocked-down rat PMVECs in vitro, the RNA-Seq was performed to explore related DEGs. The threshold of DEGs was set as p-value ≤0.05 and fold change >1.5. As the results showed, a total of 1,216 genes were identified as DEGs, among which 665 were upregulated and 551 were downregulated (Supplementary Tables S2 and S3). Top 10 upregulated and downregulated DEGs are listed in Tables 1 and 2, respectively.
Top 10 Upregulated Differentially Expressed Genes
Top 10 Downregulated Differentially Expressed Genes
To know the distribution of all DEGs intuitively, cluster analysis was carried out through following methods. Hierarchical clustering has revealed the relationship between DEGs and samples (Fig. 3a); distribution of DEGs between the samples was evaluated by the scatter plot (Fig. 3b), while the volcano plot showed the statistical difference between samples (Fig. 3c). Taken together, cluster analysis by three methods has successfully differentiated the genes between control and sample with knockdown of IQGAP1.

Cluster analysis of differentially expressed genes between shIQGAP1 and shSc.
GO analysis of DEGs
To explore the function of DEGs, GO analysis was conducted. GO terms have analyzed DEGs through matching them to MF, CCs, and biological process (bp) terms, which have revealed the possible association between function and genes. For upregulated DEGs, 234 MF terms, 176 CC terms, and 825 BP terms were upregulated. The annotations for MF were mainly about molecule binding activity such as DNA binding (GO:0003677), protein binding (GO:0005515), and nucleotide binding (GO:0000166); CC terms were mainly related to chromosome (GO:0005694) and cytoskeleton formation (GO:0005856), while BP terms were mainly annotated for entries of cell cycle (GO:0000278) and cell division (GO:0051301). Top 10 upregulated GO terms classified by MF, CC, and BP terms, respectively, are presented and ranked by fold enrichment in Figure 4a.

GO analysis of DEGs.
As for downregulated DEGs, GO annotations have revealed that 211 MF terms, 93 CC terms, and 2206 BP terms were significantly changed. Among them, MF terms mainly referred to signaling receptor binding (GO:0005102), receptor ligand activity (GO:0048018), and cytokine activity (GO:0005125); CC terms were mainly enriched in extracellular region (GO:0005576), extracellular space (GO:0005615), and extracellular matrix (GO:0031012), while BP annotations were mainly related to cell proliferation (GO:0042127) and cell migration (GO:0030334). As depicted in Figure 4b, top 10 significant differentially expressed annotations were shown with MF terms, CC terms, and BP terms by bar chart, respectively. In all, knockdown of IQGAP1 plays an important role in cellular biological function, especially in downregulation of cell BP.
KEGG pathway analysis of DEGs
To analyze related biochemical metabolic signal pathways and signal transduction pathways that most DEGs involved in, KEGG pathway analysis was performed. Significant differentially regulated pathways were determined by both p-value (p ≤ 0.05) and the number of DEGs. Compared with the control, knockdown of IQGAP1 resulted in the upregulation of 33 pathways, while a total of 73 pathways were downregulated. Among all significantly expressed pathways, DNA replication and cell cycle and repair pathways were significantly upregulated, while the cytokine-related pathways such as TNF signaling pathway, cytokine-cytokine receptor interaction, and IL-17 signaling pathway were downregulated. Top 10 upregulated and downregulated KEGG pathways are shown in Figure 5a and b, respectively. Of note, the most significantly downregulated signaling pathway was TNF signaling pathway. Top 10 downregulated DEGs enriched in TNF signaling pathway also were annotated for the function of cell proliferation, cell migration, and regulation of cytoskeleton. The linkage between these genes enriched in TNF signaling pathway is exhibited in Figure 5c. In all, knockdown of IQGAP1 has a great impact on cell cycles, TNF signaling pathway, and cellular cytokines pathways.
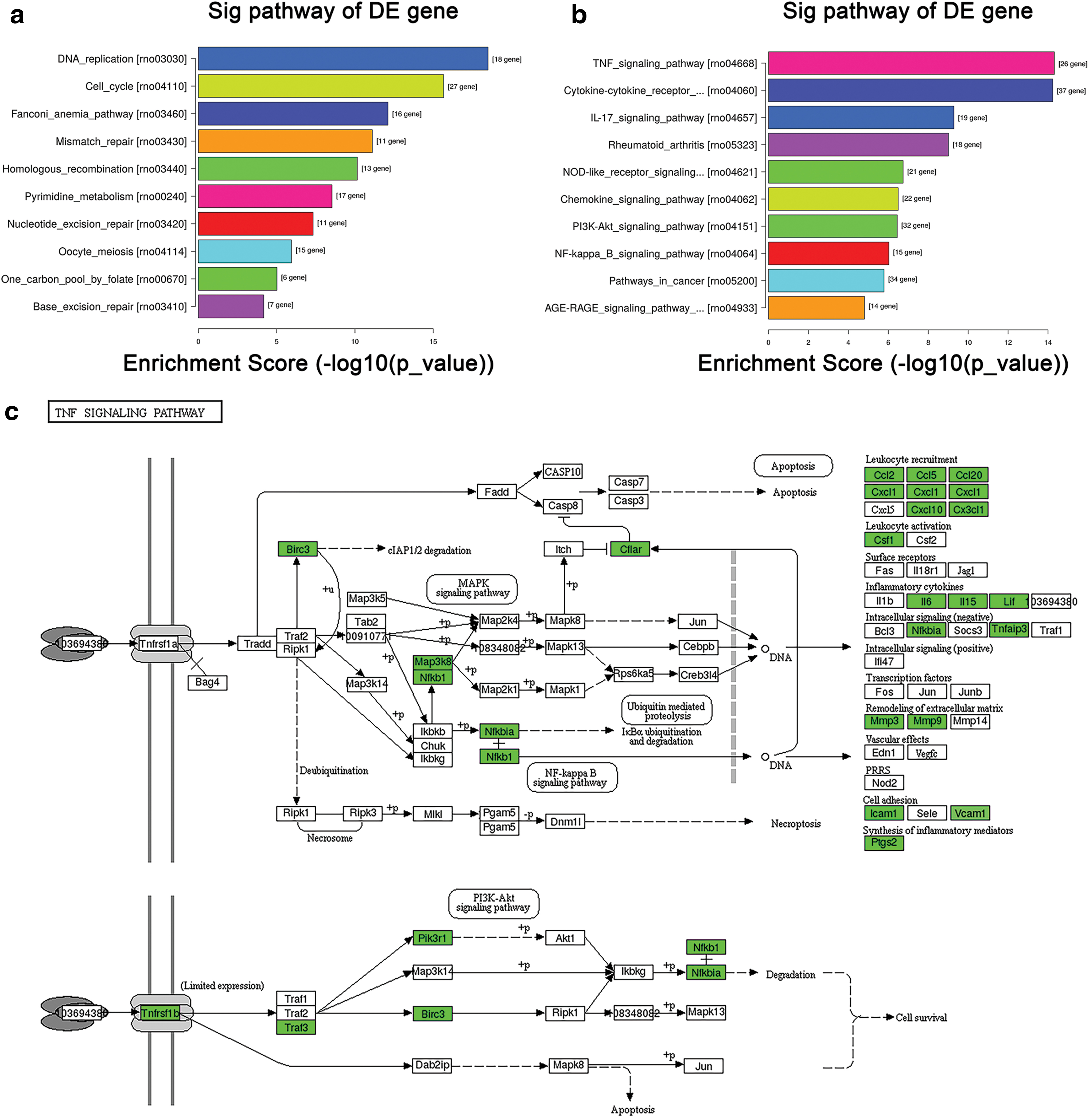
KEGG pathway analysis of DEGs.
Validation of DEGs by qRT-PCR
To validate the DEGs by RNA-Seq, we choose 30 DEGs for validation. Among them, 15 genes, Mcm3, Mcm5, Pole, Rfc2, Bub1, Cdk1, Plk1, Ttk, Orc6, Rpa1, Rfc4, Rfc5, Mcm6, Cdc6, and Cdc20, were upregulated, and Cxcl2, Ccl2, IL6, Ccl20, Lif, Ccl5, Ccl7, Cxcl1, Cxcl3, Cxcl6, Tlr2, Ptgs2, Gbp2, Mmp3, and Mmp13 were 15 downregulated genes. For 15 upregulated genes, all of them were enriched in DNA replication pathway, cell cycle, and DNA repair pathways, while 15 downregulated genes were mainly enriched in TNF signaling pathway, cytokine-cytokine receptor interaction, or Il-17 signaling pathways, and mainly involved in the regulation of inflammation, cell proliferation, and leukocyte migration, as shown in Figure 6. The results from qRT-PCR were consistent with RNA-Seq.
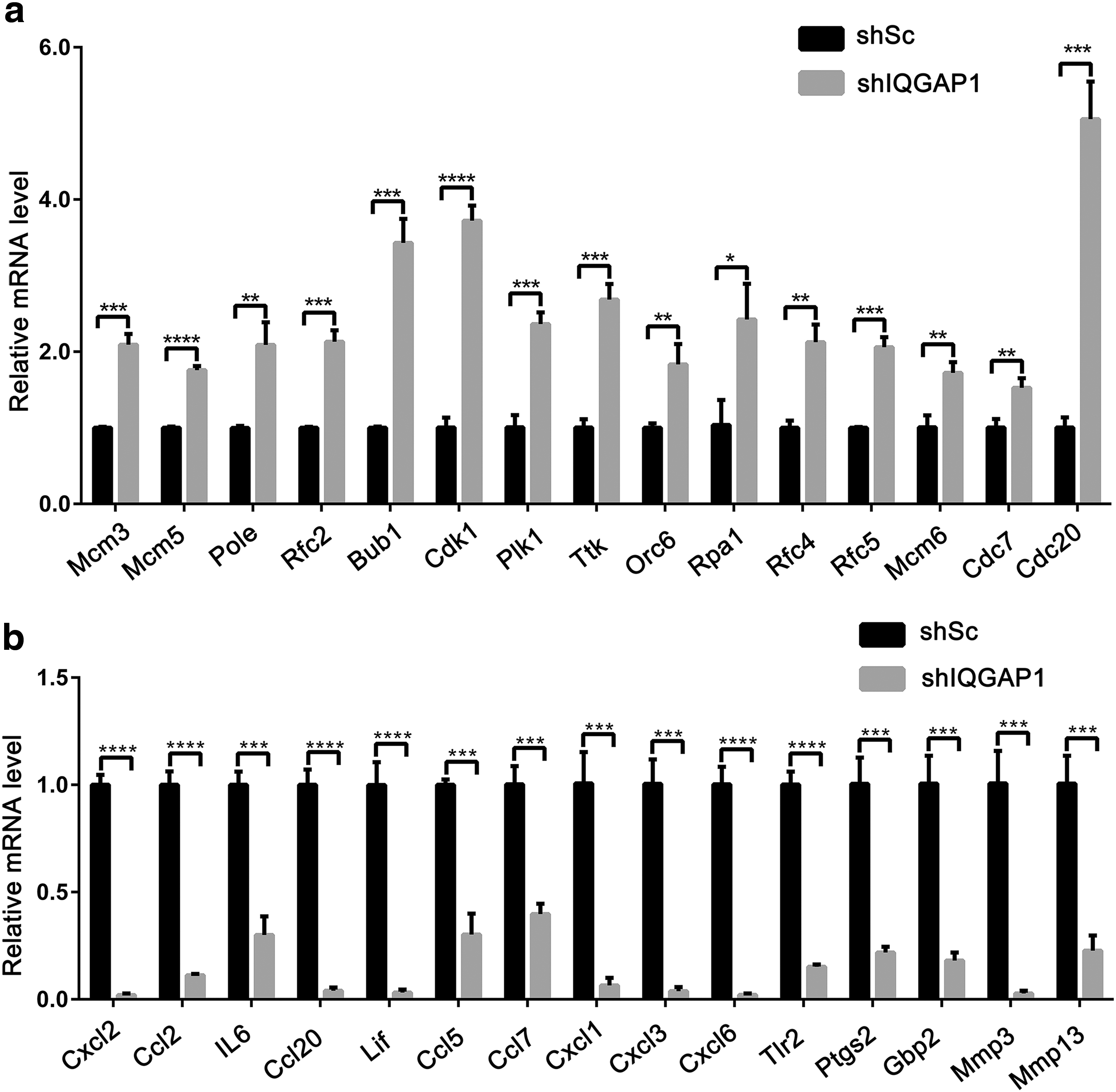
Validation of DEGs from RNA-Seq by qRT-PCR. Fifteen upregulated DEGs
Discussion
Under normal conditions, pulmonary microvascular endothelium forms a semipermeable barrier between pulmonary interstitium and pulmonary circulation, maintaining the homeostasis of lung in vivo, while destruction of the integrity of the pulmonary microvascular endothelium directly causes the leakage of blood fluid and proteins, leading to pulmonary edema, hypoxia, and inflammation. (Leligdowicz et al., 2018). Dysregulation of pulmonary microvascular endothelium barrier was initiator of uncontrolled inflammation and considered one of the most important causes of ARDS (Millar et al., 2016). Given the high genetic homology to human and economic in scientific research, Sprague-Dawley rats were widely used for reoccurrence of pulmonary diseases involving ARDS/ALI. In addition, due to the high fidelity of rat PMVECs being similar to human PMVECs, as well as the stability of rat PMVECs, rat PMVECs were extensively used to study the property of lung endothelium in vivo and in vitro (King et al., 2004). What is more, rat pulmonary microvascular endothelium has consistently expressed specific markers involving CD34 and factor VIII-related antigen in comparison to other mesothelial cells, making them easily identified and obtained (Chung-Welch et al., 1989). Considering all the above advantages, we choose rat PMVECs for the research. Based on previous studies, we successfully isolated and identified rat PMVECs through their morphology and specific biomarkers (Zhang and Sun, 2005; You et al., 2010).
IQGAP1 belongs to IQGAP family together with other two members, IQGAP2 and IQGAP3. Structure of IQGAPs family contains several specific domains, amino-terminal calponin homology domain, IQ regions, Ras GAP-related domain, and RasGAP C-terminus (Watanabe et al., 2015). In comparison to other two family members, IQGAP1 is the most well studied and ubiquitously expressed in vivo (Watanabe et al., 2015). Mounting research has explored that IQGAP1 functioned as a scaffold protein and plays an important role in maintaining the endothelium barrier in related diseases such as ARDS/ALI (Mehta et al., 2014). Besides, IQGAP1 is considered to be an oncogene, which is overexpressed in various tumors and promotes tumor proliferation and invasion. Given that, in many tumors, IQGAP1 is often overexpressed (Nabeshima et al., 2002; Wang et al., 2013), while in many endothelium barrier dysregulated diseases, expression of IQGAP1 is often downregulated (Bhattacharya et al., 2012; Hedman et al., 2015), and little research referred to DEGs and significantly regulated signaling pathways induced by the downregulation of IQGAP1. Hence, to further explore the related genes and pathways regulated by IQGAP1 in PMVECs, we knocked down IQGAP1 in PMVECs using lentivirus shRNAs. With the advent of next generation of transcriptomic RNA-Seq, more comprehensive and accurate transcripts and DEGs could be identified even for those with low expression levels (Trapnell et al., 2012; Wang et al., 2019). To our best knowledge, our research could be the first revealing DEGs in the context of dysregulated IQGAP1 in PMVECs.
By transcriptomic RNA-Seq, we have identified a total of 1,216 DEGs in regulation by IQGAP1. In comparison with the control, 665 DEGs were upregulated and 551 were downregulated. Among those genes, some genes have been reported before in destructive pulmonary microvascular endothelium regulated by IQGAP1, while many novel genes have also been discovered in our research. Of note, former studies have demonstrated that intercellular adhesion molecule-1 (ICAM-1, also called CD54) and IL-6 as biomarkers help indicate the severity of ARDS (Calfee et al., 2007). Intriguingly, ICAM-1 and IL-6 also have been significantly downregulated by knockdown of IQGAP1 (Supplementary Table S2). In normal conditions, ICAM-1 is scarcely expressed in tissues, while is significantly upregulated during inflammation (Roebuck and Finnegan, 1999). Expression of ICAM-1 in PMVECs was specific and only displays a significant upregulation in PMVECs in ARDS. Lou et al. (1998) have detected no expression of ICAM-1 in vein endothelial cells, which were separated from the ARDS patients. A similar was result also obtained by Müller (2002). Hence, higher ICAM-1 in patients who suffered from ARDS might indicate poor prognosis and increased risk of death (Flori et al., 2003). The role of ICAM-1 is to mainly mediate adhesion. Adherence of neutrophils to microvascular endothelial cells is dependent on ICAM-1, thereby promoting the dysfunction of endothelium barrier, also facilitating endothelium permeability and inflammation (Wang and Doerschuk, 2002; DiStasi and Ley, 2009). In addition, previous research has also demonstrated that IQGAP1 binds to ICAM-1 to regulate cell-cell adhesion. We can infer that IQGAP1 also mediates inflammation of the endothelium barrier in the regulation of ICAM-1. Increased expression of IL-6 in vivo facilitates the acute phase reaction and suggests a severity of disease (Geiger et al., 1988). A research by Razidlo et al. (2018) demonstrated that IQGAP1 is necessary for IL-6 to promote pancreatic tumor migration and invasive, downregulation of IQGAP1 leads to a decrease of IL-6. Our research based on the transcriptomic RNA-Seq of finding a decreased IL-6 caused by knockdown of IQGAP1 is consistent with their findings. Given those previous findings on ICAM-1, IL-6, and IQGAP1, the downregulation of ICAM-1 and IL-6 induced by knockdown of IQGAP1 in our research also suggests that IQGAP1, in regulation of these molecules, plays an important role in these biological processes such as inflammation and tumorigenesis.
GO reports have analyzed the function of DEGs induced by IQGAP1 knockdown. In our research, we found biological processes, especially cell proliferation, cell migration, cell adhesion, vascular development, and leukocyte migration, were modulated by IQGAP1. Under normal context, pulmonary microvascular endothelial cells are very active and have strong proliferation and angiogenic capacity (Guazzi et al., 2015). Proliferation of PMVECs was thought to play a vital role in controlling the restoration and pulmonary vascular homeostasis in ARDS/ALI. When injured, their powerful proliferation and angiogenic capabilities help repair endothelial function (Lv et al., 2017). Our research of downregulated biological processes related to cell proliferation and vascular development induced by knockdown of IQGAP1 suggests that IQGAP1 might exert a critical role in endothelium barrier repair and damage. In addition to cell proliferation in response to barrier injury, cell adhesion and cell migration also play an important role in protecting endothelial barrier function. A recent research by Ushio-Fukai et al. (2013) has shown the role of IQGAP1 in maintaining adhesion and structure of endothelium to protect against blood shear stress injury. Besides ICAM-1, vascular cell adhesion molecule-1 (VCAM-1 also called CD106) is also an important adhesion molecule in regulation of cell adhesion. Both of ICAM-1 and VCAM-1 were significantly downregulated as a result of IQGAP1 knockdown (Supplementary Table S2), while they all play a key role in leukocyte-endothelial adhesion, leukocyte migration, and leukocyte recruitment, especially neutrophil infiltration. Increase of ICAM-1 and VCAM-1 could be responsible for the endothelial permeability and lung edema (DiStasi and Ley, 2009). Considering the biological processes regulated by IQGAP1 described above, we speculate that IQGAP1 not only plays a key role in protecting the endothelial barrier, but also exerts an important role in inflammation.
According to CC terms in GO report, we also found that knockdown of IQGAP1 weakened the cytoskeleton, cell junction, and extracellular formation. All these cellular components are also critical for maintaining the integrity of microvascular endothelium barrier. Bhattacharya et al. have found that IQGAP1 is necessary for cell-cell junction by trafficking the integrin αvβ3. In addition, they also found that mice with depletion of IQGAP1 present an increased pulmonary vascular leakage with more protein in bronchoalveolar lavage fluid during ALI (Bhattacharya et al., 2012). Rac1 and Cdc42 are best known members of the Ras superfamily, cycling between active GTP-bound forms and inactive GDP-bound forms. It is widely studied that IQGAP1 can bind to Rac1 and Cdc42, forming a complex to regulate the cytoskeleton and cell junction. Their interaction promotes the endothelium barrier function and cytoskeleton remodeling (Watanabe et al., 2015). In our research, we also found several novel DEGs that were never reported before, such as Dlc1 and Itga1, were downregulated in the context of low expression of IQGAP1. These genes were also enriched in the regulation of cell junction and cell cytoskeleton, which provide a new vision of molecular mechanism regulated by IQGAP1, in regulation of the endothelial barrier. For MF terms, IQGAP1 closely related to the activity of some inflammatory cytokines such as IL family members and some chemokines, such as CXC and CC family members. These cytokines also play a pivotal role in the pathogenesis of ARDS.
According to KEGG pathway analysis, many pathways have been involved in the regulation by IQGAP1. The upregulated pathways mainly referred to DNA replication, cell cycle, and some repair process of endothelium. Among all downregulated pathways, some have been reported before such as PI3K-Akt, NF-κB, and MAPK signaling pathway. Also, these signals were proven to be important in the development of some diseases. For example, Sbroggiò et al. (2011) have demonstrated that IQGAP1 plays a positive role in protecting cardiac function and cardiac remodeling from the high pressure load by activating ERK1/2 and Akt signals. In gastric cancer, IQGAP1 activates ERK1/2 and NF-κB to promote tumor cell proliferation and growth (Tseng et al., 2014). Similarly, Ma et al. (2013) have found that IQGAP1 promotes cell proliferation by upregulation MAPK signaling pathway in the disease of multiple myeloma. The role of IQGAP1 in those diseases mainly correlated to cell proliferation and cell migration, which are in response to some annotations in GO report. Of note, although IQGAP1 as a scaffold interacts with these signals exerting important role in multiple cellular processes, former research also explored these signaling pathways in regulation of inflammatory process. For example, NF-κB and MAPK were found in regulation of IL-8 and ICAM-1 in human PMVECs (Su et al., 2008). In addition, many novel pathways also were identified in our research. Those pathways may also be responsible for some key pathophysiology in endothelium barrier or ARDS/ALI. For downregulated pathways induced by knockdown of IQGAP1, the most significant differential pathways were about TNF signaling pathway, cytokine-cytokine receptor interaction, and IL-17 signaling pathway, all these signaling pathways related to inflammation. Intriguingly, the top 10 significantly downregulated DEGs, such as Ccl20, Cxcl2, and IL6, are mainly involved in those pathways above. Ccl20 as proinflammatory cytokine can promote the inflammatory cell migration, as well as the development of the inflammatory disease (Hirota et al., 2007); Cxcl2 belongs to the Cxc chemokine family, the increased expression of it refers to the lung injury and the neutrophil accumulation (Belperio et al., 2002), while IL-6 as one of the most important proinflammatory mediators was considered to be one of the important biomarkers in ARDS (Xu and Song, 2017). These genes also validated by qRT-PCR and the results are consistent with the RNA-Seq.
In this research, we found that dysregulation of IQGAP1 caused a significant impact on TNF signaling pathway. TNF plays an important role in acute inflammation and is considered to be the center of the “cytokine storm” process (Azevedo et al., 2012). The inflammatory reaction induced by TNF in pulmonary endothelium is an early event in ARDS/ALI. Also, activation of TNF in PMVECs further facilitates the recruitment and infiltration of neutrophils by promoting the release of some inflammatory chemokines and adhesion molecules involving IL-8 and ICAM-1, while both of them are important biomarkers in ARDS/ALI (Bhatia and Moochhala, 2004). In addition, activation of TNF also promotes NF-κB and MAPK signaling pathways in many diseases. Abdollahi et al. (2005) have found one of the members of TNF, TRAIL, activates MAPK in ovarian cancer, while existence of IL-8 also inhibits the tumor apoptosis induced by TRAIL. Given a complex regulatory mechanism between TNF signaling pathway and others in diseases, and the contributions of IQGAP1 to TNF signaling pathway, we postulate that IQGAP1 could as a therapeutic target in those diseases. In accordance with this, a recent study by Akula et al. (2019) has found that Rac1, one of the members of Ras superfamily, interacted with IQGAP1 activates inflammatory reaction to promote the release of IL-6 and TNF in rheumatoid arthritis. They also suggest that IQGAP1 could as the target in some inflammatory diseases. Intriguingly, we also found a downregulation of rheumatoid arthritis signaling pathway induced by IQGAP1 knockdown. Although a great deal of research has paid attention on tumorigenesis involving IQGAP1, mounting evidence also indicates a chronic inflammation gives higher risk of tumorigenesis. Given that IQGAP1 caused a great impact on TNF signaling pathway and others, as well as those signals in the development of diseases, we emphasized here that IQGAP1 plays a critical role in the inflammation beyond its role in cytoskeleton, which could shed a new light for the clinical therapy in the diseases with dysregulated signaling pathways involving TNF induced by dysregulation of IQGAP1, such as ARDS/ALI.
However, the study still has some limitations. For example, among those hundreds of DEGs, some genes that are responsible for damage of endothelium barrier have already been identified, while the functions of the remaining DEGs are still poorly understood. Hence, much work is needed to explore the functions of those novel genes in the development of related disease. Furthermore, it could be more meaningful that those genes are validated in the disease animal models or patient serum. We will do further work in future research.
Conclusion
In this research, we have revealed the DEGs and signaling pathways of PMVECs induced by IQGAP1 knockdown on the basis of RNA-Seq. Among these genes, some regulated by IQGAP1 that plays a pivotal role in protecting endothelium barrier in ARDS/ALI have been demonstrated before, while many novel genes still need further research. According to GO annotations, many DEGs play a key role in cell proliferation, cell adhesion, and cell cytoskeleton. Through analyzing KEGG pathway, we found that many genes induced by IQGAP1 not only involve in the regulation of cytoskeleton but alsp exert critical roles in inflammation. We hence concluded that IQGAP1 also plays an important role in inflammatory diseases, which could provide a new insight for future study on endothelium barrier and IQGAP1-related diseases, such as ARDS/ALI.
Footnotes
Authors' Contributions
S.S., Y.C., and W.L. carried out the experiments. A.X., X.Z., and Y.W. designed the study and analyzed data. Y.W. and G.S. wrote the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by National Natural Science Foundation of China (nos. 81770081 and 81370170), the National Key Clinical Specialist Construction Programs of China (N3101005005025), and the Natural Science Foundation of Universities of Anhui Province (KJ2019A0219).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.