
Abstract
Cytochrome b6f complex is a thylakoid membrane-localized protein and catalyses the transfer of electrons from plastoquinol to plastocyanin in photosynthetic electron transport chain. In the present study, Cytochrome b6 (KaCyt b6 ) gene from Kappaphycus alvarezii (a red seaweed) was overexpressed in tobacco. A 935 base pair (bp) long KaCyt b6 cDNA contained an open reading frame of 648 bp encoding a protein of 215 amino acids with an expected isoelectric point of 8.67 and a molecular mass of 24.37 kDa. The KaCyt b6 gene was overexpressed in tobacco under control of CaMV35S promoter. The transgenic tobacco had higher electron transfer rate and photosynthetic yield over wild-type and vector control tobacco. The KaCyt b6 tobacco also exhibited significantly higher photosynthetic gas exchange (PN) and improved water use efficiency. The transgenic plants had higher ratio of PN and intercellular CO2. The KaCyt b6 transgenic tobacco showed higher estimates of photosystem II quantum yield, higher activity of the water-splitting complex, PSII photochemistry, and photochemical quenching. The basal quantum yield of nonphotochemical processes in PSII was recorded lower in KaCyt b6 tobacco. Transgenic tobacco contained higher contents of carotenoids and total chlorophyll and also had better ratios of chlorophyll a and b, and carotenoids and total chlorophyll contents hence improved photosynthetic efficiency and production of sugar and starch. The KaCyt b6 transgenic plants performed superior under control and greenhouse conditions. To the best of our knowledge through literature survey, this is the first report on characterization of KaCyt b6 gene from K. alvarezii for enhanced photosynthetic efficiency and growth in tobacco.
Introduction
The world population is growing and United Nation's reports expect it to reach 9.7 billion by 2050. The global climate changes and depletion of natural resources especially water and arable land affected natural ecosystem in various ways and these are the major challenges to meet the food security for growing population. The Food and Agriculture Organization (FAO) anticipated a requirement of 70% escalation in food production by 2050 to meet the supply for the growing population (FAO, 2016). The increase in crop yields can only meet the demands for food, feed, and energy for the growing population (Fedoroff et al., 2010; Reynolds et al., 2011). Photosynthesis fixes solar energy into NADPH and ATP (chemical energy) through electron transfer and employ this energy for biomass production. Photosynthetic efficiency is critical to growth, development, and yield in crop plants and it has been a promising target to enhance the plant productivity and yield (Miyagawa et al., 2001; Yamori, 2013; Yadav et al., 2018). The photosynthetic efficiency is an outcome of plant's ability to harvest the sun light, efficient transformation of solar energy into chemical energy, and amount of the chemical energy fixed as biomass (Yadav et al., 2018). In addition to improving crop biomass and yield, the enhanced photosynthesis would help to cope up with global warming through utilizing the main greenhouse gas that is, CO2. Literature reports are available on attempts to improve the productivity and yield in crops by improving leaf photosynthesis (Murchie et al., 2009; Reynolds et al., 2009; Makino, 2011; Raines, 2011). In addition to traditional practices, different targets and approaches have been identified and reviewed to enhance the photosynthetic efficiency in crops (von Caemmerer and Evans, 2010; Raines, 2011; Reynolds et al., 2011; Parry et al., 2013; Singh et al., 2014; Mathan et al., 2016). Therefore, improving the photosynthetic efficiency can be an emerging route to improve the crop productivity to feed the growing population (Long et al., 2015).
In the recent past, genetic engineering has been widely used to enhance photosynthesis and growth of plants by overexpressing transgenes encoding for the enzymes, pathways, or transporters (Murchie and Niyogi, 2011; Karki et al., 2013; Parry et al., 2013; Price et al., 2013). Rice, potato, tobacco, Arabidopsis, and soybean were genetically engineered for improved CO2 fixation and growth through introgression of genes encoding enzymes of Calvin–Benson cycle (Lefebvre et al., 2005; Feng et al., 2007; Rosenthal et al., 2011; Kandoi et al., 2016). Arabidopsis mutant for R-type anion channel (AtQUC1) showed enhanced photosynthesis, stomatal and mesophyll conductance, and accumulated higher contents of organic acid (Medeiros et al., 2016). Engineering of tobacco and soybean with cyanobacterial fructose-1,6-/sedoheptulose-1,7-bisphosphatase (FBP/SBPase) improved CO2 assimilation and growth (Miyagawa et al., 2001; Köhler et al., 2017). Similarly overexpression of Brachypodium distachyon SBPase gene improved the biomass yield in wheat (Driever et al., 2017). The overexpression of the Rubisco large (LS) and small (SS) subunits with the Rubisco assembly chaperone RUBISCO ASSEMBLY FACTOR 1 (RAF1) resulted in ∼30% increase in Rubisco and 15% increase in CO2 assimilation along with increased fresh weight (FW) in UBI-LSSS-RAF1 transgenic plants (Salesse-Smith et al., 2018). Engineering of tobacco with gene encoding CO2-concentrating mechanism (CCM) from Synechococcus elongates improved the photosynthesis (Orr et al., 2020). The synthetically designed alternative photorespiratory pathways along with carbon concentrating cycles have also been hypothesized to enhance the crop productivity (Naseem et al., 2020).
The genetic engineering for increased photosynthetic electron transfer, NADPH, and ATP contents resulted in enhanced photosynthesis and growth of transformed plants (Hajirezaei et al., 2002; Chida et al., 2007; Rodriguez et al., 2007; Yadav et al., 2018). The overexpression of combination of Arabidopsis violaxanthin de-epoxidase (VDE), PSII subunit S (PsbS), and zeaxanthin epoxidase in tobacco resulted in enhanced photoprotection and improved photosynthesis (Kromdijk et al., 2016). Algal cytochrome c6 enhanced photosynthesis and growth in Arabidopsis (Chida et al., 2007) and tobacco (Yadav et al., 2018). Rieske FeS protein overproduction improved the functioning of photosystem I (PSI) and PSII, and enhanced the maximum carbon assimilation in Arabidopsis (Simkin et al., 2017). Genetic engineering in tobacco for improved PsbS showed reduced loss of water molecule per CO2 molecule assimilation that is, improved water use efficiency (WUE) (Głowacka et al., 2018). The cytochrome b6/f complex, ATP synthase and Ferredoxin-NADP(+) reductase are major components of photosynthetic electron transfer chain. The cytochrome b6/f complex and ATP synthase play a key role in the generation of NADPH and ATP for CO2 fixation. The CO2 fixation has been reported proportional to the Rubisco and Cyt b6/f proteins (Onoda et al., 2005; Yamori et al., 2005). The chloroplast electron transport rates decreased more severely with reductions in cytochrome b6/f complex as compared with ATP synthase activity in tobacco (Anderson et al., 1997; Ruuska et al., 2000) and this confirmed direct relationship of cytochrome b6/f with photosynthetic electron transfer. Thus, cytochrome b6/f is a key rate-limiting step (Rott et al., 2011) in chloroplast electron transport and is a potential target for enhancing photosynthetic performance of crops (Yamori et al., 2011).The algae are aquatic plants with huge variability and possess comparatively higher growth rate due to their better photosynthetic performance (Zou and Gao, 2010). Algae acclimatize to different light intensities by adjusting its photosynthetic functioning due to higher numbers of reaction centers (RCs) supporting the efficient electron transport (Sukenik et al., 1987). Algae offer a number of potential genes in genetic engineering of crops to improve photosynthetic efficiency, productivity, and biomass. In the recent past, cytochrome c6 from Porphyra species (Chida et al., 2007), Ulva fasciata (Yadav et al., 2018), and CCM from Chlamydomonas reinhardtii (Nölke et al., 2019) were engineered in Arabidopsis and tobacco for betterment of photosynthesis. The cytochrome b6 are important components of electron transfer and algae provides an advantage to study the mechanisms and regulation of photosynthetic electron flow. The cytochrome b6 is a major subunit of cytochrome b6/f complex and transfers electrons between the PSII and PSI RCs in photosynthetic electron transfer chain and contribute electrochemical gradient through pumping protons into the thylakoid space for ATP synthesis. Due to rate-limiting components in chloroplast electron transport, the cytochrome b6 offers an opportunity in genetic engineering of crops to enhance the photosynthetic efficiency. Kappaphycus alvarezii (Doty) Doty ex P.C.Silva is a red seaweed and it exhibits average daily growth rate ranging from 3.64% to 13.98% (Rao et al., 2008). Considering higher daily growth rate and commercial acceptability, in the present study, K. alvarezii was used as a source of cytochrome b6 gene. The overexpression of Cytochrome b6 gene (KaCyt b6 ) from K. alvarezii in plants was expected to achieve the enhanced photosynthesis through improved electron transfer and, thus, higher productivity in crop plants. The present investigation aimed at cloning KaCyt b6 from K. alvarezii and its overexpression in transgenic tobacco for enhanced photosynthetic yield and growth. To the best of literature survey, it is the first report on characterization of a cytochrome b6 protein from K. alvarezii in tobacco to improve the photosynthetic and growth performances. The T1 transgenic tobacco plants showed higher electron transfer and CO2 assimilation rate, and improved accumulation of photosynthetic pigments, soluble sugar, and starch contents. The present results showed KaCyt b6 as a potential gene and photosynthetic electron transport as a potential target site for genetic engineering to enhance photosynthetic efficiency and crop yield. The study provided new insights in the improvement of photosynthetic and crop yield; and this would add to the development of molecular breeding strategies to meet the requirement of higher crop productivity for global food security.
Materials and Methods
Seaweed collection, RNA extraction, and cDNA synthesis
The present research work didn't involve any kind of human participants (cells or tissues) therefore this does not require any ethics committee approval. K. alvarezii thallus was collected from CSIR-CSMCRI seaweed experimental farm at Simbor (N 20°46′12.00′′; E 71°09′27.00′′), Una district, Gujarat. The samples were washed with autoclaved seawater, frozen in liquid nitrogen, and immediately processed for RNA isolation. The total RNA was extracted following cetyltrimethylammonium bromide (CTAB)-LiCl method (Zeng and Yang, 2002) with minor modifications. Briefly, 1 g thallus tissue was ground in liquid nitrogen in a prechilled pestle and mortar. The homogenate was transferred to a centrifuge tube containing prewarmed (65°C) 10 mL of CTAB buffer [2.0% (w/v) each CTAB and polyvinylpyrrolidone (PVP), 100 mmol Tris-HCl (pH 8.0), 25 mmol EDTA, 2.0 M NaCl, and 2.0% β-mercaptoethanol] and mixed well through vortexing. Samples were incubated at 65°C for 30 min with regular mixing. The samples were centrifuged at 11,000 rpm for 10 min at 18°C and supernatant was collected in a centrifuge tube. To this equal volume of chloroform:isoamyl alcohol (24:1 v/v) was added, mixed gently, and centrifuged at 13,000 rpm for 15 min at 4°C. The upper aqueous phase was collected in a fresh tube and the above step was repeated. To the upper phase, 1/3 volume of 8.0 M LiCl was added, mixed well, and incubated at −20°C overnight. The samples were centrifuged at 13,000 rpm for 15 min at 4°C and supernatant was discarded. The RNA pellet was washed twice with 80% ethanol. The pellet was dried and resuspended in 40 μL ultrapure RNase-free water and stored at −80°C. The quantity and quality of the extracted RNA was measured by NanoDrop spectrophotometer ND-1000 (NanoDrop Technologies) and agarose gel electrophoresis. The cDNA was synthesized using 1 μg of total RNA with ImProm-II™ Reverse Transcription System (Promega) and used as a template for PCRs.
Molecular cloning of full-length KaCyt b6 gene
The sequences of algal Cyt b6 genes were retrieved from NCBI database. Using the conserved sequences in the retrieved sequences, the degenerate primers for cloning of KaCyt b6 gene were designed and purchased from Sigma-Aldrich (India). The KaCyt b6 gene was partially amplified using degenerate primers (F 5′-GTGAACTAACATGGGTTACAGGAG-3′; R 5′-ATTTCTAATGGTGTAGCAAATGGA -3′) through PCR following the standard PCR program in a thermal cycler (Bio-Rad). The purified amplicon was cloned in pGEM®-T vector (Promega Corporation) and sequenced (Macrogen, Inc., Seoul, South Korea). The KaCyt b6 gene was made full length from both the ends through rapid amplification of cDNA ends (RACE). The 5′ end of the gene was completed following 5′-RACE protocol as per instructions of the manufacturer (Invitrogen, San Diego, CA). The gene-specific primer1 (GSP1; 5′-CTACGATGGGAACTGCTTCAGG-3′) and Superscript RTII were used to synthesize the first strand of cDNA. From reaction mixture, the RNA template was degraded using RNase H and the cDNA was purified using a column. The purified cDNA was subjected to terminal deoxynucleotidyl transferase reaction to add the homopolymer dC tail at 3′ end of the cDNA. The dC tailed cDNA was amplified with GSP2 (5′-CTGCCAAAATGACTCCTGTAACC-3′) and an abridged anchor primer (AAP; 5′-GGCCACGCGTCGACTAGTAC(G)16-3′) supplied with the kit. For 3′ end amplification of KaCyt b6 gene, the first strand of cDNA was synthesized with PK1 oligo dT adapter primer (AP1; 5′-CCAGTGAGCAGAGTGACGAGGACTCGAGCTCAAGC(T)7-3′). Subsequently, the first strand of cDNA was amplified with 3GSP1 (5′-GGAGAACCAGCATGGCCAAATGA-3′) and PK2 (AP2; 5′-CCAGTGAGCAGAGTGACG-3′). Nested PCR with the above PCR product was performed using 3GSP2 (5′-GCCATCAGCAATAGGAGAACAAGC -3′) and PK3 (AP3; 5′-GAGGACTCGAGCTCAAGC-3′) primers. The amplified product of 5′- and 3′-RACE reactions were cloned and sequenced for confirmation. The full length of KaCyt b6 sequence was obtained through alignment of the sequences of partially cloned fragment, 5′- and 3′-RACE products. Using aligned sequence, primers for full-length cloning of KaCyt b6 gene were designed. The full-length KaCyt b6 gene was cloned using Pfu and Taq DNA polymerase (2:1) (Invitrogen) and full-length primers (forward primer-5′-GAATAACCATGGGCAAAGTATATG -3′; reverse primer-5′-CGCTCGAGGAATTCCTACCAACCTTTTTC -3′). The amplified product was cloned into in pGEM-T vector and sequenced for confirmation. The KaCyt b6 gene sequence was submitted to the GenBank database.
Bioinformatics analysis
The in silico analysis of KaCyt b6 gene was performed using various online tools. The BLAST-N and P programs were used for homology comparisons and prediction of the conserved domains, respectively. The amino acid composition and physiological properties were predicted using ProtParam tool. The PSIPRED server and FoldIndex program were used for prediction of secondary structure of KaCyt b6 and its folding characteristics, respectively. The membrane topology and subcellular localization were predicted with TMpred program and localizome server, respectively. The multiple sequence alignments of nucleotide and predicted protein sequences were carried out using ClustalW program. The evolutionary relationship among the KaCyt b6 and different Cyt b6 homologs from various algae and plant species was determined using MEGA 5 software (Tamura et al., 2011). The phylogenetic tree was constructed by the neighbor-joining method. Bootstrap analysis with 1000 replicates was conducted to optimize the confidence levels for the branches. The ORF in full-length cloned KaCyt b6 gene was searched using ORF finder tool.
Construction of the plant expression vector and tobacco transformation
Full-length KaCyt b6
was cloned using K. alvarezii cDNA as template and DNA polymerase with primers Cyt bF (5′-
Molecular and β-glucuronidase analysis of transgenic tobacco
The putative transgenic tobacco lines were confirmed through β-glucuronidase (GUS) expression and PCR amplification of KaCyt b6 gene, hygromycin (hptII) selection marker, and gus (uidA) reporter gene. The GUS expression in the leaves of tobacco plants was assessed using the GUS Reporter Gene Staining Kit (Sigma). GUS staining was performed by dipping the leaves of wild-type (WT), vector control (VC), and transgenic tobacco lines into GUS staining buffer, vacuum infiltration of the stained buffer-dipped leaves for 5 min, and finally incubation overnight at 37°C in the dark. The stained leaves were bleached with 80% ethanol and documented under microscope. Genomic DNA was isolated from WT, VC, and transgenic tobacco leaves following the CTAB method. Quality and quantity of the genomic DNA was determined by agarose gel electrophoresis and NanoDrop Spectrophotometer, respectively. The molecular analysis of the transgenic tobacco lines was performed through PCR amplification of the genomic DNA with primers specific to KaCyt b6 , hptII, and uidA gene. The PCR product was analyzed on 1.2% agarose gel.
Physiological characterization of transgenic tobacco
The T0 seeds of VC and transgenic lines (L3 and L5) overexpressing KaCyt b6 were germinated on 0.75% agar-gelled hormone-free (HF) MS medium supplemented with 20 mg l−1 hygromycin. Separately, the seeds of WT seeds were germinated on HF MS basal medium. Two-week-old seedlings of WT, VC, and transgenic tobacco lines were transplanted into soil and acclimatized for 2 weeks. After acclimatization of WT, VC, and transgenic tobacco lines, plant height, chlorophyll content, total soluble sugar, and starch were estimated at 15th, 30th, 45th, and 60th days. The photosynthetic gas exchange and chlorophyll fluorescence parameter were also measured on the 45th day.
Determination of leaf area, water content, and photosynthetic pigments
The shoot length and leaf area of 6-week-old WT, VC, and transgenic tobacco lines were observed. The FW of leaves was measured and these leaves were oven dried at 90°C to record the dry weight (DW). The water content of leaves was calculated with (FW−DW)/(DW) × 100 and expressed as % of g−1 DW. The photosynthetic pigments are indicators of relative greenness and photosynthetic efficiency in plants. The photosynthetic pigment contents were estimated in WT, VC, and T1 transgenic tobacco lines grown under greenhouse condition. The photosynthetic pigment contents were extracted by homogenizing the leaf samples of WT, VC, and T1 transgenic tobacco line in N,N-dimethylformamide (DMF) under diffused light. The 100 mg leaf samples were crushed in DMF and centrifuged at 13,000 rpm for 10 min at 4°C. The supernatant was collected and absorbance was read at 664, 664.5 647, and 461 nm using a UV-Visible spectrophotometer (SpectramaxPlus 384). Chlorophyll a and b, total chlorophyll, and carotenoid contents were calculated following Inskeep and Bloom (1985) and Chamovitz et al. (1993). The ratio of chlorophyll a and b (Chl a/b), and carotenoid/total chlorophyll contents (Caro/TC) were determined.
Estimation of soluble sugar and starch contents in tobacco overexpressing KaCyt b6 gene
The fresh samples were ground to powder in liquid nitrogen and repeatedly extracted in 80% ethanol at 4°C. The ethanol extract was evaporated until dryness and residue was dissolved in deionized water for sugar estimation. The residual pellet left after ethanol extraction was used for starch content estimation. The residual pellet was digested in 52% perchloric acid and the digest was centrifuged at 10,000 rpm for 30 min. The supernatant was diluted with milliQ water and used for starch quantification. The soluble sugar and starch contents were determined following anthrone reagent (0.2% anthrone in ice-cold 95% sulfuric acid) method. The absorbance was read at 630 nm against a standard curve prepared with glucose. In the case of the starch estimation, the obtained value was multiplied by a factor of 0.9 to convert the sugar values into starch contents.
Gas exchange and chlorophyll fluorescence measurements in KaCyt b6 tobacco
The gas exchange and chlorophyll fluorescence were simultaneously measured in leaves of control and transgenic tobacco using fluorometer attached to a portable infrared gas analyzer (Li-6400XT) with an open system (Li-Cor). Measurements were recorded on the dorsal surface of the fully developed leaves (fourth leaf from top) in 45 days old soil-transplanted WT, VC, and transgenic lines (L3 and L5) under ambient CO2 conditions and at a constant irradiance of 1000 mmol photons m−2 s−1. The plants were dark adapted for 45 min using dark room facilities. The minimal (F0) and maximal (FM) fluorescence in the dark-adapted state was measured using a 0.1 μmol photons m−2 s−1 and saturating (>3000 μmol photons m−2 s−1) actinic light, respectively, for ≤1 s duration. The measurements in dark-adapted WT, VC, and transgenic tobacco lines were recorded under diffused light conditions. The plants were exposed under 1200 μmol m−2 s−1 for 2 h in a plant growth chamber (PGC-105; Percival Scientific) to achieve the steady-state fluorescence. Gas exchange measurements were carried out at 1000 μmol m−2 s−1 PPFD, ambient atmospheric CO2 (380 μmol−1 mol−1), and 60–65% RH and 26°C block temperature. With gas exchange measurement net photosynthetic rate (PN; μmol of CO2 m−2 s−1), transpiration rate (Tr; mmol of H2O m−2 s−1), intracellular CO2 (Ci, μmol of CO2 m−2 s−1), stomatal conductance (gs, μmol of CO2 m−2 s−1), and ratio of PN and Ci were determined. WUE (μmol CO2 mmol−1 H2O) was calculated as the ratio of photosynthesis and transpiration (PN/Tr). With fluorescence measurements, minimal (F0), maximal (FM), variable (FV) fluorescence, activity of the water-splitting complex (FV/F0), basal quantum yield of nonphotochemical processes in PSII (F0/FM), maximum efficiency of PSII (FV/FM), variable to maximal fluorescence (FV’/FM’), redox state of the plastoquinone pool/nonphotochemical quenching (1-qP), PSII quantum yield/PSII photochemistry/photosynthetic yield (φPSII), photochemical quenching (qP), non-qP (qN and NPQ), and electron transfer rate (ETR) were determined in WT, VC, and transgenic (L3 and L5) lines (Baker, 2008).
Data recording and statistical analysis
During the course of investigations, each experiment was performed thrice with five biological replicates. Data were subjected to one-way ANOVA for analysis of variance to determine the significance among mean values of control [WT/negative control (WT) and positive control (VC)] and transgenic tobacco plants using SIGMA plot 13. The statistical significance among the control (WT and VC) and transgenic tobacco plants was calculated at p ≥ 0.05. Data were presented as mean ± standard error and the statistically significant differences among mean values were denoted as different lower case letters.
Results
Molecular cloning of KaCyt b6 cDNA and in silico analysis
In the present study, full-length Cyt b6 gene was cloned from 14 different seaweeds comprising 4 red and 5 each of green and brown seaweeds and details of these cDNA sequences have been provided as Supplementary Table S1. Furthermore, based on natural growth rate and importance of K. alvarezii in global seaweed cultivation program, Cyt b6 gene from K. alvarezii (KaCyt b6 ) was functionally characterized in planta for enhanced photosynthesis and yield in plants through transgenic approach. Full-length KaCyt b6 gene was cloned successfully from K. alvarezii through 5′ and 3′ rapid amplification of cDNA ends (RACE) approach. The KaCyt b6 cDNA (GenBank accession no. KT184318) was 935 bp long. It had a 43 bp 5′ untranslated region (UTR), a 648 bp ORF, and a 244 bp 3′UTR (Supplementary Fig. S1a). Cloned ORF encoded a protein of 215 amino acids with an expected molecular weight of 24.37 kDa and an isoelectric point of 8.67. The amino acid sequence showed maximum (97%) identity with Grateloupia tenuistipitata Cyt b6 protein. Hydropathicity analysis showed four transmembrane (TM) domains in KaCyt b6 protein (Supplementary Fig. S1b). FoldIndex program revealed 3 disordered regions with 23 disorder-promoting amino acid residues in KaCyt b6 protein (Supplementary Fig. S1c). Secondary structure prediction of KaCyt b6 protein revealed 9 α-helixes and 1 strand in peptides (Supplementary Fig. S1d). Phyre2 software revealed 81% α-helixes, 0% β-sheets, and 43% TM helixes in secondary structure and predicted 5% disorder (Supplementary Fig. S1e). ScanProsite predicted four iron (heme)-binding (86, 100, 187 and 202) sites in KaCyt b6 protein. Supplementary Figure S2a showed predicted 3D structure of KaCyt b6. Localizome server predicted KaCyt b6 to be a transmembrane protein (Supplementary Fig. S2b). KaCyt b6 showed possibility of phosphorylation with protein kinase-(PK) C, PK-A, cyclin-dependent kinase 2 (cdc2), caseine kinase II (CKII), and an unspecific protein (un-sp). Highest score was 0.94 for un-sp at position T-61 and S-89. NetPhos 3.0 server search for overall phosphorylation site exhibited three serine, two threonine, and two tyrosine phosphorylation sites (Supplementary Fig. S2c).
Construction of KaCyt b6 expression vector and tobacco transformation
The ORF of KaCyt b6 was cloned in a binary vector pCAMBIA1301 under the control of constitutive promoter 35S (Fig. 1a). Tobacco regenerates from cocultured leaf discs on medium containing hygromycin were considered as putative KaCyt b6 transgenic tobacco and these exhibited positive GUS histochemical assay. Soil-transplanted rooted plantlets of GUS-positive tobacco were grown until maturation in containment facility and seeds were harvested. Transgenic tobacco exhibiting positive PCR amplification of KaCyt b6 gene, hptII, and UidA genes (Fig. 1b) were selected as confirmed transgenic tobacco. Amplification of 648 bp KaCyt b6 ORF in transgenic tobacco confirmed its integration and overexpression under CaMV35S promoter. Seedlings of WT raised on MS medium and of VC and T1 transgenic lines raised on hygromycin-supplemented MS medium (Fig. 1c) were used for further gene function analysis. T1 seedlings of VC and transgenic tobacco exhibited GUS staining (Fig. 1d). These were subsequently confirmed through PCR amplification of KaCyt b6, UidA, and hptII genes as that of T0 tobacco. The transgenic lines L3 and L5 along with WT and VC were used for further transgenic analysis.
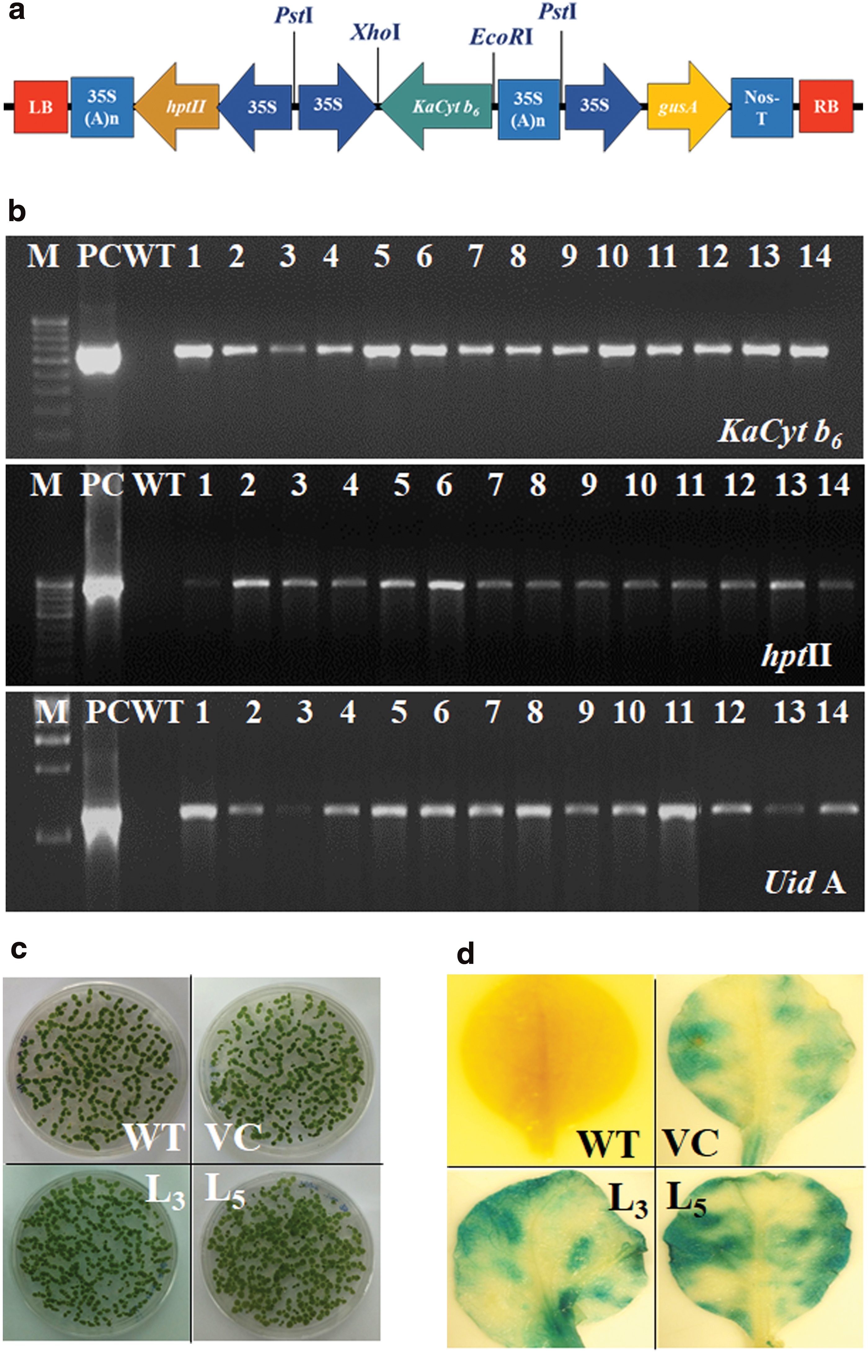
Genetic transformation of tobacco: Schematic diagram of the KaCyt b
6—pCAMBIA1301 construct
Transgenic tobacco overexpressing the KaCyt b6 showed improved growth attributes
The transgenic tobacco overexpressing KaCyt b6 gene exhibited comparatively higher leaf area, water contents, and plant growth as compared with WT plants. T0 transgenic tobacco plants during acclimatization under containment facility exhibited enhanced growth as compared with WT and VC plants (Supplementary Fig. S3). Similarly, the T1 transgenic tobacco showed enhanced growth under in vitro conditions (Fig. 2). Similar to T0 transgenic tobacco, soil-transplanted T1 transgenic tobacco also exhibited improved growth (Fig. 3a–d) and better phenotypic and photosynthetic appearance of leaves in terms of size (Fig. 3e–h) as compared with control plants during growth period. Shoot growth in T1 transgenic lines was estimated significantly higher as compared with control plants (Fig. 4a). In consonance with growth, T1 transgenic tobacco had higher contents of water (Fig. 4b). Transgenic tobacco overexpressing KaCyt b6 grew normal and had comparatively bigger leaves and better flowering during their growth cycle (Supplementary Fig. S4). The present results clearly confirmed the enhanced growth of transgenic tobacco overexpressing KaCyt b6 gene under controlled and greenhouse conditions.

Pictorial observation on plant growth under controlled conditions on 0.75% agar-gelled hormone-free MS medium after 15 days

Pictorial observations of plant growth
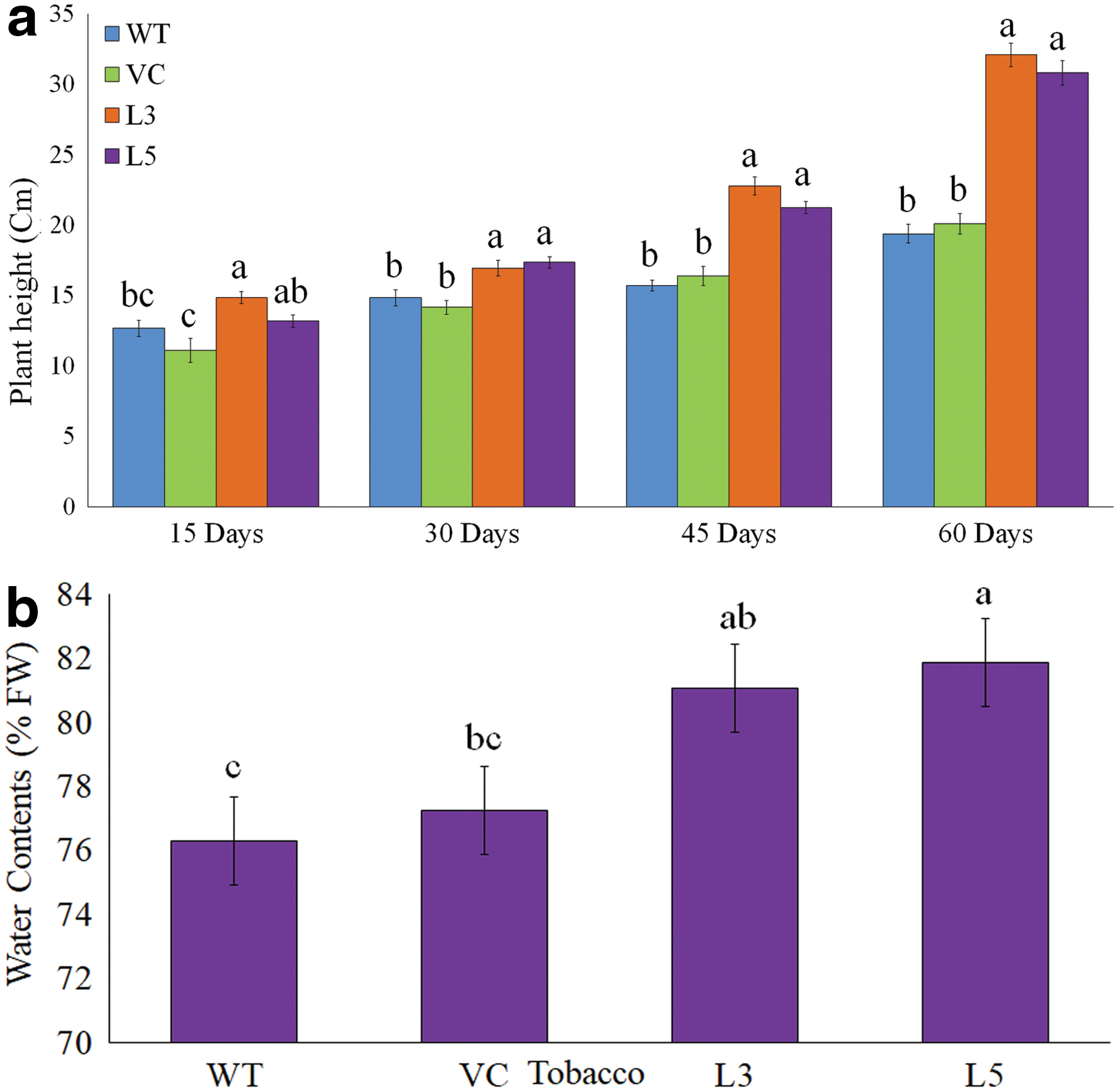
Plant height during different time intervals
Photosynthetic pigments estimated higher in tobacco overexpressing KaCyt b6
The transgenic tobacco overexpressing KaCyt b6 had higher contents of total chlorophyll, chlorophyll a, chlorophyll b, and carotenoid contents as compared with WT and VC tobacco. The contents of chlorophyll a and b were estimated higher in transgenic tobacco overexpressing KaCyt b6 (Supplementary Fig. S5a, b). Similarly, the T1 transgenic (L3 and L5) tobacco at different time intervals exhibited significantly higher contents of total chlorophyll and carotenoids as compared with WT and VC plants (Fig. 5a, b). In consonance with contents of chlorophyll and carotenoid contents, ratios of chlorophyll a and b (chlorophyll a/b), and carotenoids and total chlorophyll contents (carotenoids/total chlorophyll contents) were estimated significantly lower in KaCyt b6 transgenic tobacco as compared with WT and VC tobacco (Fig. 5c, d).

Total chlorophyll contents
Overexpression of KaCyt b6 in transgenic tobacco enhanced photosynthesis
In continuation of photosynthetic pigments, simultaneous photosynthetic gas exchange and chlorophyll fluorescence were studied in WT, VC, and transgenic tobacco overexpressing KaCyt b6 . The PN was significantly higher in KaCyt b6 transgenic tobacco lines (L3 and L5) as compared with WT and VC tobacco under ambient conditions (Fig. 6a). The T1 transgenic tobacco compared with control tobacco exhibited comparatively higher gs (Supplementary Fig. S6a). The Ci was estimated comparatively lower in transgenic tobacco as compared with WT and VC tobacco (Supplementary Fig. S6b). However, the differences in the values of gs and Ci of control and transgenic tobacco were nonsignificant. The PN/Ci was calculated higher in KaCyt b6 transgenic tobacco as compared with WT and VC tobacco (Fig. 6b). The E and WUE were recorded slightly higher in transgenic tobacco overexpressing KaCyt b6 as compared with WT and VC tobacco, however, the increase was not statistically significant (Supplementary Fig. S6c, d). The F0 and FM were estimated lower in KaCyt b6 transgenic tobacco lines than WT and VC plants (Supplementary Fig. S6e, f), however, the differences were nonsignificant. The FV/FM (Fig. 6c) and FV/F0 (Fig. 6d) were comparatively higher than control plants in transgenic tobacco lines. The F0/FM was comparatively lower in transgenic tobacco (L3 and L5) lines (Fig. 6e). The FV’ did not vary significantly in transgenic and control tobacco plants (Supplementary Fig. S6g). The KaCyt b6 transgenic tobacco lines exhibited comparatively higher FV’/FM’ as compared with WT and VC tobacco (Supplementary Fig. S6h). The KaCyt b6 transgenic tobacco overexpressing KaCyt b6 showed higher estimates of PSII photochemistry (ɸPSII), ETR, and qP as compared with control plants (Fig. 6f–h). The NPQ was estimated slightly lower in KaCyt b6 transgenic tobacco than WT and VC plants (Supplementary Fig. S6i), however, the differences were nonsignificant. The estimates of 1–qP supported the lower estimates of non-qP in transgenic tobacco overexpressing KaCyt b6 (Supplementary Fig. S6j). The KaCyt b6 transgenic tobacco overexpressing KaCyt b6 exhibited higher ETR (Fig. 6g) and photosynthetic yield as compared with control plants. The photosynthetic gas exchange and chlorophyll fluorescence measurements clearly showed the enhanced photosynthetic performance and ETR in transgenic tobacco overexpressing the KaCyt b6 gene as compared with control tobacco (WT and VC) plants.
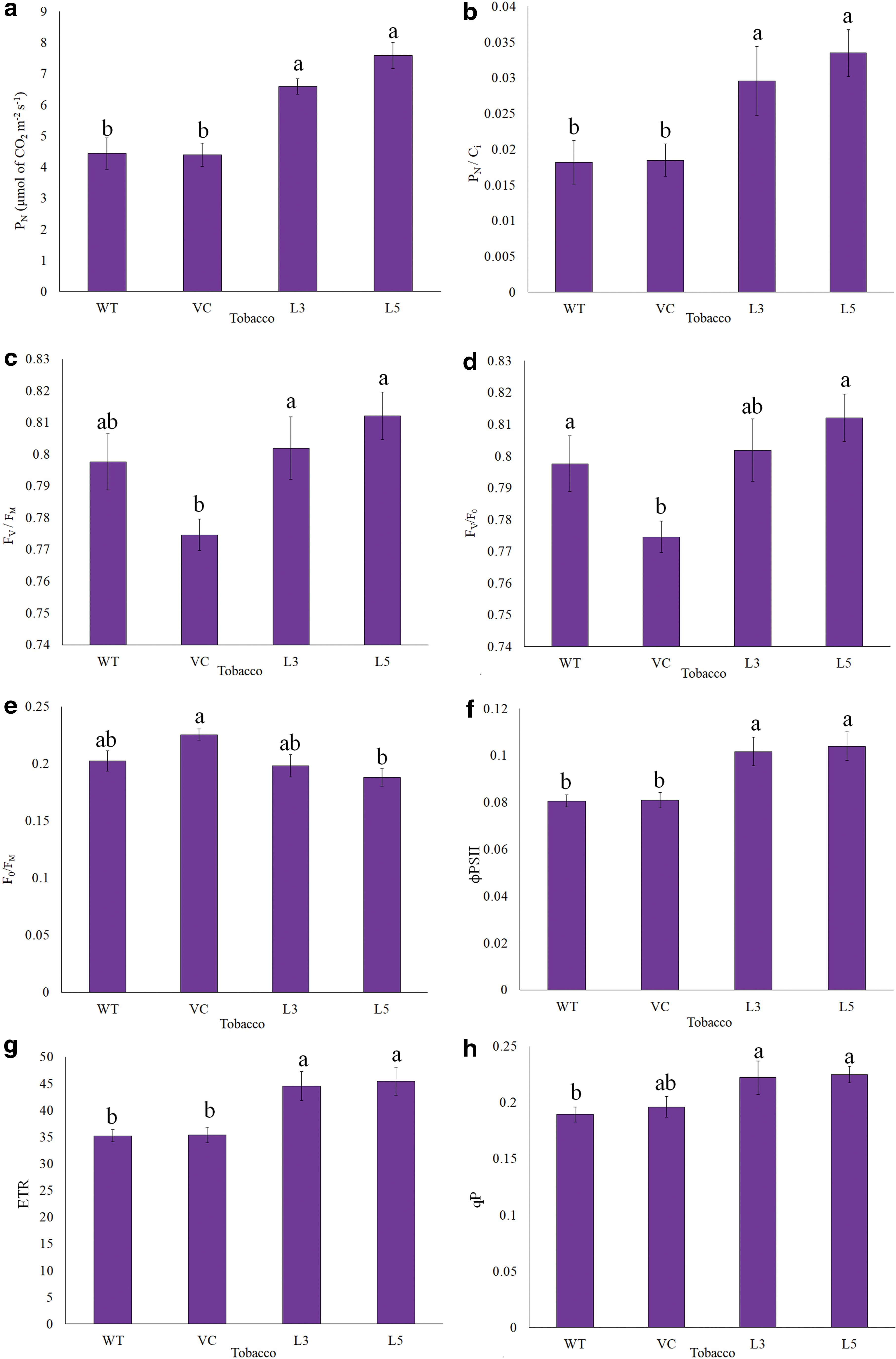
Net photosynthesis rate
KaCyt b6 tobacco accumulated higher amounts of sugar and starch
The transgenic tobacco overexpressing KaCyt b6 gene accumulated higher contents of ultimate photosynthetic products. The accumulation of total soluble sugar and starch contents differed with age of plants. Compared with WT and VC tobacco, the KaCyt b6 transgenic tobacco accumulated significantly higher contents of soluble sugar (Fig. 7a). In consonance with sugar contents and photosynthetic rate, transgenic tobacco accumulated significantly higher contents of starch as compared with control plants (Fig. 7b).
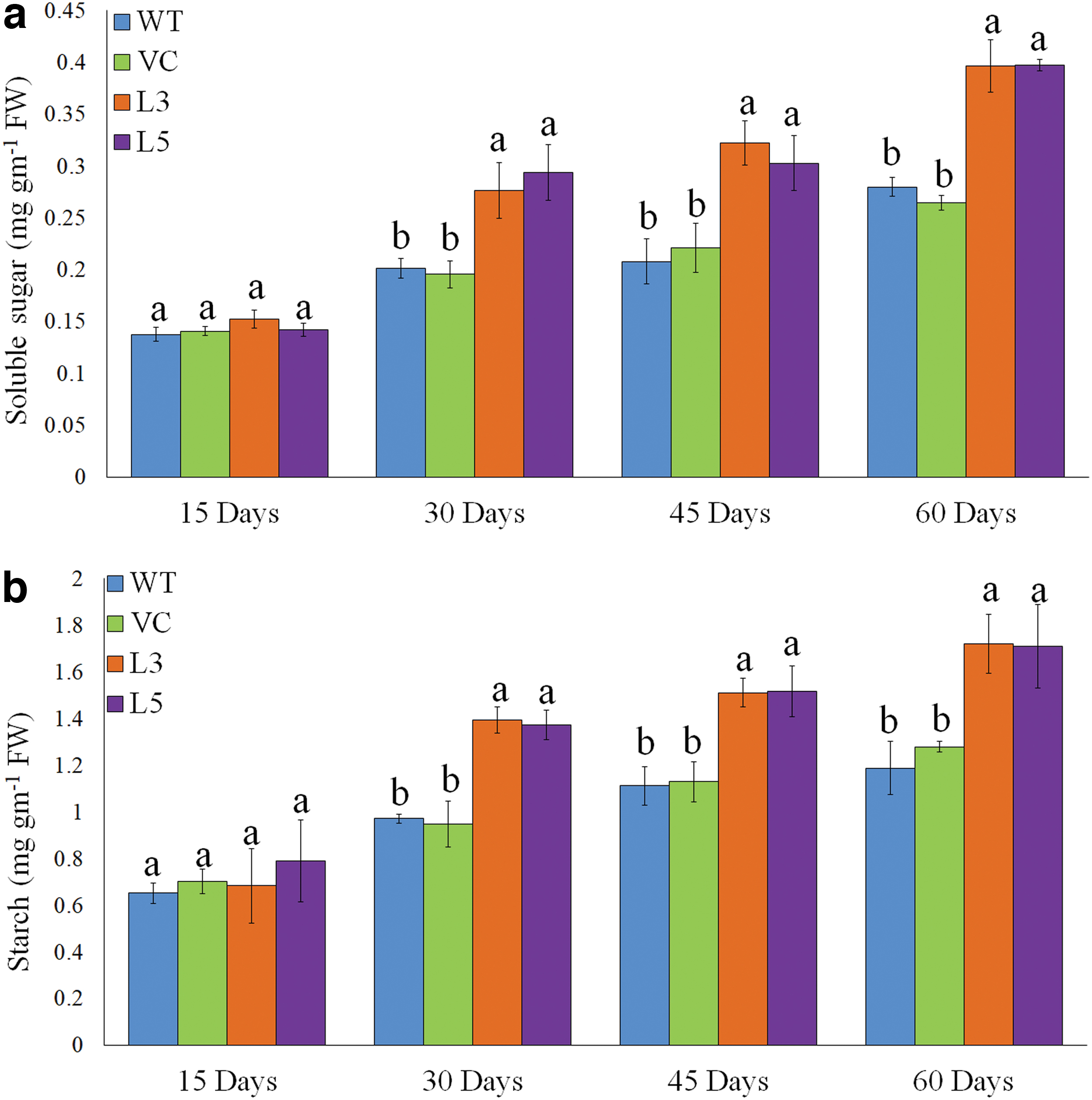
Total soluble sugar
Discussion
Improved photosynthetic efficiency results enhanced crop productivity and yield. The conventional plant breeding has limitations to improve photosynthesis efficiency, however, past many years' breeding efforts maximized the plant architecture to support light capture. The different reports on enhancement of photosynthetic capacity through genetic engineering approach will be useful in the coming decades to increase the photosynthetic efficiency in plants.
In the present study, KaCyt b6 was overexpressed in tobacco for enhanced photosynthesis and biomass yield through acceleration of photosynthetic electron transfer. The PCR results showed integration of KaCyt b6 and its overexpression under CaMV35S promoter in transgenic tobacco. The Cytochrome b6f complex is central to electron transport chain and plays essential role in cyclic and linear photophosphorylation. This creates a proton gradient to synthesize ATP and maintain the ratio of ATP/NADPH for carbon fixation. The higher ETR in tobacco overexpressing KaCyt b6 indicated the improved electron transport from PSII to Calvin cycle (Kalaji et al., 2017). The higher FV’/FM’ and qP along lower NPQ in tobacco overexpressing KaCyt b6 indicated prominence of linear electron flow and balance between electron transport and utilization of NADPH (Kalaji et al., 2014; Chang et al., 2017). The increased ETR and qP in transgenic plants maintained the oxidized state of QA and open PSII RCs (Stefanov et al., 2016). The lower 1-qP in KaCyt b6 tobacco protected the PSII from overexcitation and thus reduced the degree of photoinhibition (Quigg et al., 2012; Kalaji et al., 2017). The expression of KaCyt b6 in transgenic tobacco improved the transfer of electrons from plastoquinol to plastocyanin, thus electron transfer. The improved electron transfer supported higher quantum yield of PSII activity in KaCyt b6 in transgenic tobacco. The transfer of higher light energy toward PSII center in KaCyt b6 transgenic tobacco resulted in higher PSII activity (Ermakova et al., 2019). The lower F0 (redox state of the PQ pool) indicated active reaction centers and higher flow of electron from LHCII to PSII reaction center (Kalaji et al., 2011, 2017). The lower FM indicated enhanced photoprotection with an increase in NPQ (Bolhar-Nordenkampf et al., 1989). The higher FV/FM (maximum quantum yield of PSII photochemical activity) in transgenic lines can be correlated with higher pigment content, net photosynthetic rate, stomatal conductance, transpiration rate, efficient flow of electron from light-harvesting complex to PSII, PSII structural integrity, efficient interaction between PSII reaction centers and peripheral antennae, active oxygen-evolving complex, activation of photoprotective regulatory mechanisms and activation of photosynthetic electron transport chain (Wientjes et al., 2013; Sharma et al., 2015; Kalaji et al., 2017). The lower value of FV/FM in WT and VC indicated the amount of closed PSII RCs (Wientjes et al., 2013). Higher FV/F0 (water-splitting complex efficiency) indicated higher light energy conversion efficiency in transgenic lines (Zhao et al., 2017). Higher FV/F0 indicated improved electron transport to augment the water-splitting complex (Pereira et al., 2000). The present results showed that overexpression of KaCyt b6 in tobacco might have led to the increased production of sugar and starch contents in the transgenic tobacco plants due to availability of sufficient reducing power for Calvin cycle in the form of NADPH and ATP. These observations are in agreement with Chida et al. (2007) and our previous study (Yadav et al., 2018) using UfCyt c6 gene from Ulva fasciata for enhanced photosynthesis and biomass.
Pigment contents (total chlorophyll, chlorophyll a, chlorophyll b, and carotenoid) are important physiological parameters directly affecting photosynthetic performance in plants. The dark green color of leaves and pigment estimation indicated higher contents of photosynthetic pigments in transgenic plant overexpressing KaCyt b6
as compared with WT and VC tobacco. The higher photosynthetic pigments supported better light capture and directly improved the photosynthetic rate in KaCyt b6
transgenic tobacco (Simkin et al., 2017). The higher carotenoid contents supported improved light-harvesting capacity in KaCyt b6
tobacco (Simkin et al., 2017) and protected KaCyt b6
tobacco from oxygen and excessive light-induced damages (Peixoto et al., 2002). The chlorophyll a/b ratio indicates the PSII/PSI contents and lower ratio indicates higher efficiency of PSII as compared with PSI in KaCyt b6
tobacco. The lower chlorophyll a/b ratio in KaCyt b6
tobacco indicated increased stacks of thylakoid membranes, density of light-harvesting complex and larger antenna size indicating the optimum utilization of lower internal intensity (Evans, 1987; Stefanov et al., 2016). The higher ɸPSII estimates indicated efficient utilization of light as photochemical energy (Kalaji et al., 2014). The lower NPQ indicated lower thermal dissipation in transgenic plants (Ermakova et al., 2019) and higher CO2 assimilation rate (Kromdijk et al., 2016). The higher qP in transgenic lines indicated better separation of charges in reaction center, electron transport, PSII yield, and fraction of open PSII centers (Maxwell and Johnson, 2000; Guo et al., 2006; Mao et al., 2007; Kalaji et al., 2017). Photosynthesis in plants depends on stomatal and nonstomatal activities. Stomatal activity provides exchange of gases through transpiration and stomatal conductance to be fixed as carbohydrate. The transgenic tobacco has shown to have higher transpiration due to stomatal opening
The results showed that enhancement of photosynthesis resulted in increased growth of transgenic tobacco overexpressing KaCyt b6 under ambient climatic conditions. The improved growth of KaCyt b6 transgenic tobacco can be directly correlated with higher rate of photosynthetic gas exchange and higher contents of soluble sugar and starch. The higher starch contents showed conversion of trapped solar energy as biomass in transgenic tobacco (Ambavaram et al., 2018). Similar results were reported in Panicum virgatum overexpressing PvBMY1 and PvBMY3 (Ambavaram et al., 2018), rice overexpressing Rieske FeS protein of cytochrome (Cyt) b6/f complex (Yamori et al., 2016), and transgenic tomato plants overexpressing SBPase activity (Ding et al., 2016). In agreement with the present results, the cyt b6/f complex may improve the CO2 assimilation rate around the PSII (Yamori et al., 2016), hence biomass and yield. Similarly, Arabidopsis overexpressing Rieske FeS showed improved photosynthesis, biomass, and seed yield (Simkin et al., 2017). In the present case, the enhanced levels of sugar and starch content could be correlated with higher photosynthetic rate (Yin et al., 2015) and growth of KaCyt b6 transgenic tobacco. The higher sugar content in KaCyt b6 tobacco contributed to enhanced antioxidant defense potential, osmotic adjustment, maintenance of cellular structure and redox potential, plant metabolism, and development (Sperdouli and Moustakas, 2012). The increased soluble sugar maintains positive turgor pressure, which prevented dehydration and directly helped in leaf and growth expansion (Hasanuzzaman et al., 2017). The higher contents of starch can be correlated with the efficient supply of ATP and NADPH, which are utilized as energy molecules in the Calvin cycle (Chida et al., 2007) and supported higher synthesis of sugar and starch, hence growth of transgenic tobacco. The results of gas exchange and fluorescence measurements suggested that the overexpression of KaCyt b6 in transgenic tobacco could be responsible for the increased photosynthetic efficiency in transgenic tobacco as compared with control (WT and VC) tobacco. The results clearly supported the fact that the photosynthetic electron transport rate can be improved through genetic engineering of the cyt b6/f complex and this is one of the potential target site for enhancing photosynthetic efficiency and crop productivity.
Conclusion
Footnotes
Acknowledgments
CSIR-CSMCRI PRIS-15/2020. S.K.Y. and K.K. are thankful to the Academy of Council of Scientific and Industrial Research (AcSIR), Ghaziabad (INDIA) for registration in PhD program.
Disclosure Statement
No competing financial interests exist.
Funding Information
Authors thankfully acknowledge the Council of Scientific and Industrial Research (CSIR), Government of India (CSC0116) for XIIth five-year plan project and Gujarat State Biotechnology Mission (GSBTM) (80G2DT/GAP 2080) Government of Gujarat for financial support. K.K. acknowledges financial support from CSIR, Government of India, in the form of CSIR-SRF.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.