
Abstract
Formin-like genes (FMNLs) are members of formins family and have been implicated to the development and progression of multiple cancers. This research aims to analyze the expression profiles, prognostic values, and immune infiltrating associations of FMNLs in gastric cancer (GC) using multiple online bioinformatics website, including Oncomine, UALCAN, Kaplan–Meier Plotter, TIMER, GeneMANIA, DAVID, and LinkedOmics databases. The mRNA levels of FMNL1/2/3 were higher in GC tissues than normal. Meanwhile, FMNLs expressions tend to be upregulated in advanced and poorly differentiated GC. Prognostic value analysis suggested that high transcription levels of FMNL1/3 were associated with poor overall survival in GC patients. Correlation analysis between FMNLs expressions and immune infiltrating GC revealed that the expressions of FMNLs were significantly associated with immune infiltrating. Protein–protein interaction network and enrichment analysis of FMNLs in GC showed that FMNLs coexpressed genes mainly participated in organizing actin cytoskeleton through affecting small G proteins activity. Moreover, Gene Set Enrichment Analysis (GSEA) analysis uncovered FMNLs and their coexpressed genes was tightly associated with immune-related cellular functions. These findings demonstrate that FMNLs might play significant immunomodulatory roles in tumor immunity and could be novel therapeutic targets and potential prognostic biomarkers in GC.
Introduction
Gastric cancer (GC) is one of the most common malignancies in the digestive system. It has the fifth highest incidence and the third mortality rate of all human cancers (Bray et al., 2018). Surgery is the only curative strategy for patients with GC at early stages. However, over 70% of patients were diagnosed at advanced stages due to the lack of specific symptoms of early GC (Song et al., 2017). Therefore, it is essential to identify novel biomarkers for the early diagnosis and prognostic assessment for patients with GC.
Formins act as crucial regulators of actin and microtubule cytoskeletal dynamics during cell division and migration and are involved in various tasks, including cell polarization, adhesion, movement, and division (Chesarone et al., 2010; DeWard et al., 2010). The dysregulation of formins is suggested to loosen adhesion of cancer cells, migration, and ultimately invasion (Kuhn and Geyer, 2014). There are seven subfamilies of formins, which have been identified already, including DRFs (diaphanous-related formins), FMNLs (formin-like protein), DAAM (disheveled-associated activator of morphogenesis), INF (inverted formin), FHOD (formin homology domain-containing protein), and FMN (formin) (DeWard et al., 2010).
As main members of formin proteins, FMNL1, FMNL2, and FMNL3 might be implicated in cell polarity control, migration, and invasion through mediating the ras homolog family protein (Rho)-related signaling pathways (Katoh and Katoh, 2003). However, the exact expression patterns and prognostic values of FMNLs in GC have not been defined yet.
In recent years, as many interactive and user-friendly online platforms based on the TCGA database established and applied, an increasing number of tumor biomarkers have been recognized (Bai et al., 2020; Li et al., 2020). In this study, we analyzed the mRNA level of FMNLs and correlation with prognosis in GC through multiple online databases. Besides, we investigated the correlation of FMNLs with tumor-infiltrating immune cells in the tumor microenvironments through Tumor Immune Estimation Resource (TIMER) database. Our results demonstrated that FMNLs play important roles in GC prognostic evaluation and the expression of FMNLs might modulate tumor immunity by regulating the infiltrating of immune cells in GC.
Materials and Methods
Oncomine database analysis
The mRNA levels of FMNLs in various types of cancers were identified in the Oncomine database (Rhodes et al., 2004). The threshold was determined according to the following values: p value of 0.001, fold change of 1.5, and gene ranking of top 10%. The p value was calculated using the Student's t-test.
UALCAN database analysis
UALCAN is a comprehensive web resource for analyzing cancer omics data (Chandrashekar et al., 2017). In this research, the mRNA levels of FMNLs in GC as well as in various tumor subgroups based on individual cancer stages and tumor grades were analyzed by UALCAN.
Kaplan–Meier Plotter database analysis
Kaplan–Meier Plotter was applied to assess the prognostic value of FMNLs (Lanczky et al., 2016). This online database is capable of assessing the effect of 54,000 genes on survival using more than 10,000 cancer samples. The largest datasets include breast (n = 6234), ovarian (n = 2190), lung (n = 3452), and gastric (n = 1440) cancer. The relationship between FMNLs expressions and survival rates in GC was assessed, and the hazard ratio (HR) and log-rank p value were displayed online. The patients' cohorts were split at the median expression of FMNL mRNA level. The subtype analysis of prognostic values of FMNLs in GC based on gender, T/N/M stages, clinical stage, Lauren classification, differentiation, HER2 status, and treatment was also analyzed.
TIMER database analysis
TIMER database is a comprehensive resource for systematical analysis of immune infiltrates across diverse cancer types (Li et al., 2017). We explored the association between the levels of FMNLs gene expression and the abundance of infiltrating immune cells, including B cells, CD8+ T cells, CD4+ T cells, macrophages, neutrophils, and dendritic cells (DCs). Meanwhile, correlations between FMNLs expressions and gene markers of tumor-infiltrating immune cell subtypes were also explored. Tumor-infiltrating immune cells' abundance were displayed on the x-axis and FMNLs genes were on the y-axis. The gene expression level was displayed with log2 RSEM. In addition, we used the tumor purity for p value correction.
Protein–protein interaction network construction
GeneMANIA is an interactive and visual online protein–protein interaction (PPI) prediction tool, which provides customizable function of the detection of genes with similar functions (Mostafavi et al., 2008; Franz et al., 2018). GeneMANIA constructed PPI networks in terms of physical interaction, coexpression, predicted, colocalization, common pathway, genetic interaction, and shared protein domains. In this research, GeneMANIA was applied for PPI analysis of FMNL family members.
Database for Annotation, Visualization, and Integrated Discovery database analysis
The Database for Annotation, Visualization, and Integrated Discovery (DAVID) was employed to perform gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis of FMNLs and their cooperators, which identified in GeneMANIA (Huang et al., 2007). The human genome (Homo sapiens) was selected as the background variable. Enrichment terms were considered statistically significant when the false discovery rates were less than 0.05.
Linked Omics database analysis
Linked Omics is a publicly available portal that includes multiomics data from all 32 TCGA Cancer types (Vasaikar et al., 2018). The “LinkInterpreter” module was used to perform GO and KEGG analysis of FMNLs based on Gene Set Enrichment Analysis (GSEA). The criterion about GSEA is as follows: a minimum number of genes (size) of 3 and a simulation of 500. Top 5 terms of GO and KEGG analysis were exhibited.
Statistical analysis
Gene expression data from the Oncomine and UALCAN databases were analyzed using Student's t-test. Survival curves were produced by the Kaplan–Meier plots and were compared by log-rank test. The correlation analysis was evaluated in the TIMER database using Spearman's correlation analysis. p values when less than and/or equal to 0.05 were considered as statistically significant.
Results
The mRNA expression levels of FMNLs in GC and other cancers
The Oncomine database was used to compare the mRNA levels of FMNLs between the tumor and normal tissues. This analysis revealed that the FMNLs expressions were higher in GC tissues compared with the normal tissues (Fig. 1A). According to the information from the datasets in Oncomine, the mRNA levels of FMNL1 was 2.410, 1.493, 1.124 times higher in GC tissues with different histological types than normal tissues in D'Errico's (D'Errico et al., 2009), Chen's (Chen et al., 2003), and Cho's (Cho et al., 2011) datasets, respectively (Table 1).
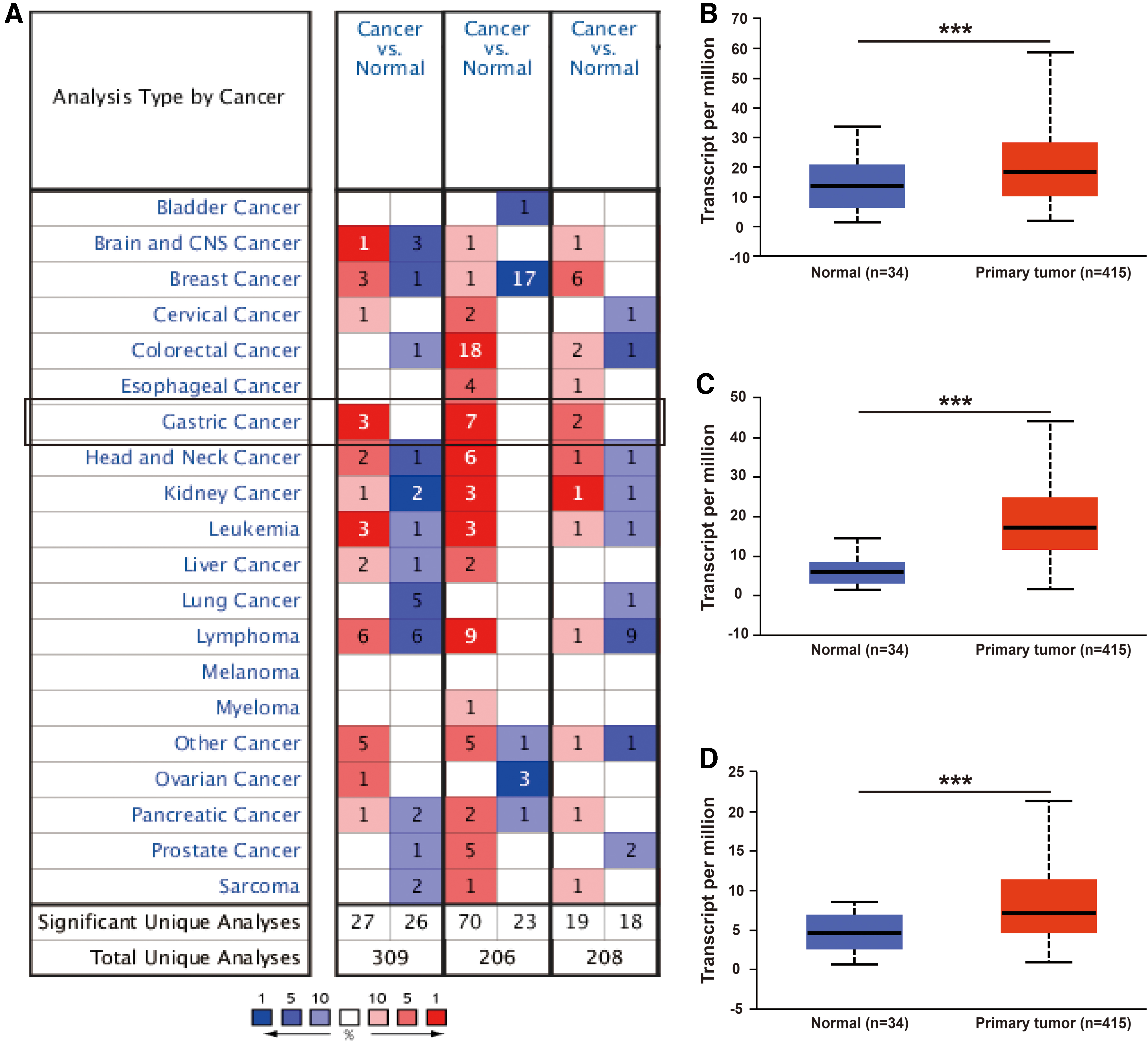
The expression of FMNLs in different cancers.
The Significant Changes of FMNLs Expression in mRNA Level Between Different Types of GC and Stomach Tissues
FMNL, formin-like gene; GC, gastric cancer.
In Chen's (Chen et al., 2003) dataset, the expression of FMNL2 was 2.060, 1.963, and 1.728 times higher in GC tissues with different histological types than normal tissues, respectively (Table 1). In Cho's (Cho et al., 2011) dataset, the expression of FMNL2 was 1.473 and 1.392 times higher in intestinal GC and mixed GC tissues than normal tissues, respectively (Table 1). In D'Errico's dataset, the expression of FMNL2 was 2.002 times higher in intestinal GC tissues than normal tissues (Table 1). In Cui's (Cui et al., 2011) dataset, the expression of FMNL2 was 1.349 times higher in general GC tissues than normal tissues (Table 1).
In Cui's (Cui et al., 2011) dataset, the expression of FMNL3 was 1.379 times higher in general GC tissues than normal tissues (Table 1). In Cho's (Cho et al., 2011) dataset, the expression of FMNL3 was 1.115 times higher in general GC tissues than normal tissues (Table 1).
In addition, the UALCAN database was also used to compare the mRNA expression of FMNLs between GC and normal tissues in TCGA samples. The expression levels of FMNLs were significantly higher in GC than they were in normal tissues (Fig. 1B–D) and the results were consistent with those of the Oncomine database.
Subtype analysis of mRNA expression levels of FMNLs in GC
To analyze the mRNA levels of FMNLs in subtypes of GC patients, UALCAN database was applied. According to clinical stages, the mRNA level of FMNL1 was higher in stage 2–4 than normal tissues and highest expression was found in stage 2 tissues (Fig. 2A). The mRNA level of FMNL2 was higher in stage 1–4 than normal tissues and highest expression was found in stage 3 tissues (Fig. 2B). The mRNA level of FMNL3 was higher in stage 2–4 than normal tissues (Fig. 2C). According to tumor grades, the mRNA level of FMNL1 was higher in grade 3 than normal and grade 1&2 tissues (Fig. 2D). The mRNA level of FMNL2 was higher in all tumor grades than normal tissues (Fig. 2E). The mRNA level of FMNL3 was higher in grade 3 than grade 2 and normal tissues (Fig. 2F). Taken together, the mRNA levels of FMNLs were higher in advanced and poorly differentiated GC.

Correlation between FMNLs expression and clinical stage together with tumor grade in GC (UALCAN). Patients who were in advanced stages tended to express higher mRNA expression of FMNLs.
Prognostic values of FMNLs in GC
To further investigate the prognostic values of FMNLs in GC, Kaplan–Meier Plotter database was applied. The results displayed that the high mRNA levels of FMNL1 and FMNL3 were correlated with poor outcomes in patients with GC (Fig. 3A, C), while the expression of FMNL2 had no association with the prognosis of GC patients (Fig. 3B).
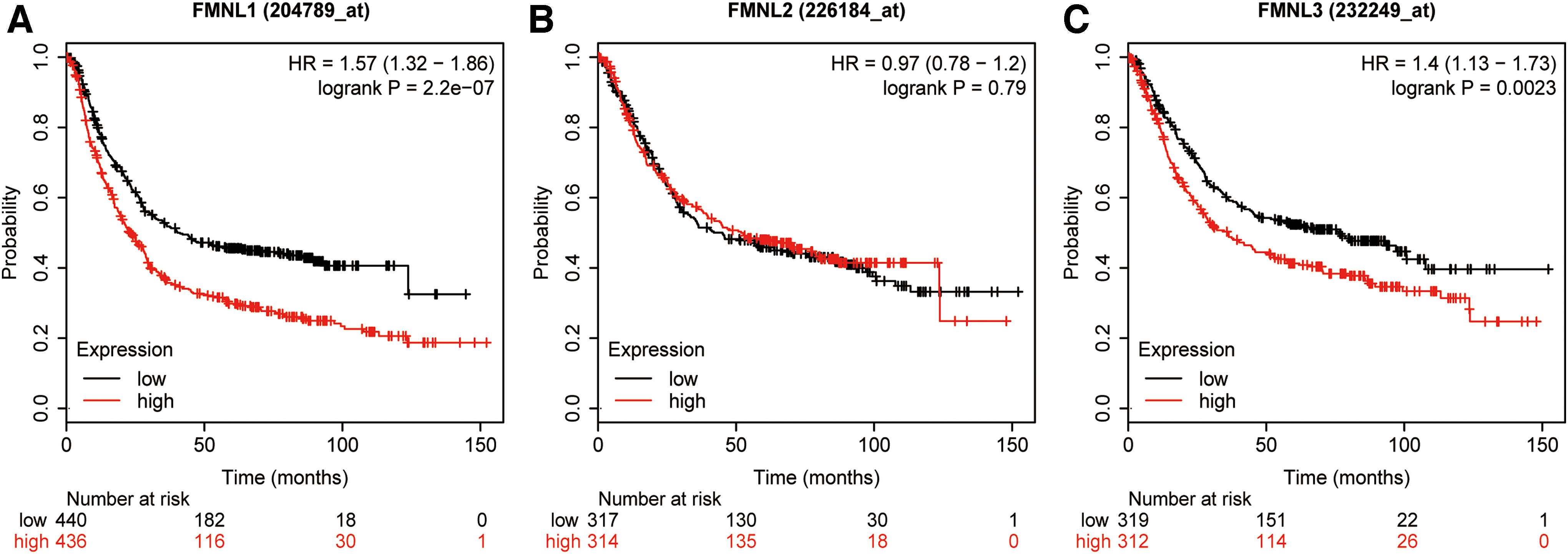
The prognostic value of mRNA levels of FMNLs in patients with GC (Kaplan–Meier plotter).
The prognostic values of FMNLs in different subtypes of GC were also assessed, including gender, T stage, N stage, M stage, clinical stage, Lauren classification, differentiation, and HER2 status. As shown in Table 2, high mRNA level of FMNL1 suggested poor prognosis in both male and female GC patients. High mRNA level of FMNL3 suggested poor prognosis in male GC patients. High mRNA expression level of FMNL1 was associated with poor outcomes in GC patients with N1 and M0, and clinical stage 3. While high mRNA expression level of FMNL2 showed better outcomes in GC patients with clinical stage 1, high mRNA expression level of FMNL3 showed poor outcomes in GC patients with T3, N1, and M0.
Correlation of FMNLs mRNA Expression and OS in GC with Different Clinicopathological Factors
HR, hazard ratio; OS, overall survival.
In terms of Lauren classification, high mRNA expression levels of FMNL1&3 were related to shorter overall survival (OS) in intestinal GC patients. High mRNA expression level of FMNL2 suggested shorter OS in diffuse GC patients. As to differentiation, high mRNA expression level of FMNL1 was related to poor OS in well-differentiated GC patients and FMNL3 in poorly differentiated GC patients suggested shorter OS. In terms of HER2 status, high mRNA level of FMNL1 suggested worse prognosis in HER2-negative and FMNL3 in HER2-positive GC patients.
Correlation between FMNLs expression and immune infiltrating level in GC
Tumor-infiltrating lymphocytes have been used to predict sentinel lymph node status and survival in cancers (Azimi et al., 2012; Dabrosin et al., 2019). Hence, TIMER database was used to investigate the relationship between the levels of immune infiltrating and the expressions of FMNLs in GC patients. As shown in Figure 4, high levels of FMNL1&3 mRNA expression had a significantly negative correlation with tumor purity (FMNL1, r = −0.278, p < 0.001, FMNL3, r = −0.214, p < 0.001) in GC.

Correlation of FMNLs expression with immune infiltrating level in GC. FMNL1&3 expressions were significantly negatively related to tumor purity. While FMNL1–3 expressions were significantly positively correlated with infiltrating levels of CD8+ T cells, CD4+ T cells, macrophages, neutrophils, and DCs in GC. DC, dendritic cell. Color images are available online.
High mRNA level of FMNL1 was negatively correlated with infiltrating levels of B cells (r = −0.116, p = 0.026). Whereas high expressions of FMNL1–3 were significantly positively correlated with infiltrating levels of CD8+ T cells (FMNL1, r = 0.601, p < 0.001, FMNL2, r = 0.146, p = 0.005, FMNL3, r = 0.432, p < 0.001), CD4+ T cells (FMNL1, r = 0.496, p < 0.001, FMNL2, r = 0.114, p = 0.029, FMNL3, r = 0.606, p < 0.001), macrophages (FMNL1, r = 0.481, p < 0.001, FMNL2, r = 0.240, p < 0.001, FMNL3, r = 0.640, p < 0.001), neutrophils (FMNL1, r = 0.667, p < 0.001, FMNL2, r = 0.177, p < 0.001, FMNL3, r = 0.492, p < 0.001), and DCs (FMNL1, r = 0.781, p < 0.001, FMNL2, r = 0.236, p < 0.001, FMNL3, r = 0.660, p < 0.001). These results strongly suggested that FMNLs played specific roles in regulating immune infiltrating in GC.
Correlation analysis between FMNL expression and immune marker sets
To investigate the relationship between FMNLs and the diverse immune infiltrating cells, we analyzed the correlations between FMNLs and immune markers of various immune cells, including CD8+ T cells, T cells (general), B cells, monocytes, tumor-associated macrophages (TAMs), M1 and M2 macrophages, neutrophils, NK cells, DCs, T helper 1 (Th1) cells, Th2 cells, and follicular helper T (Tfh) cells in GC using the TIMER database. Results showed that the FMNLs mRNA expression levels were significantly related with the immune cells (Table 3).
Correlation Analysis Between FMNLs and Relate Gene Markers of Immune Cells
cor, correlation coefficient revealed by Pearson test; TAM, tumor-associated macrophage; Tfh, follicular helper T; Th1, T helper 1.
Specifically, the FMNL1 mRNA level showed significantly positive correlation with the expression of markers of specific immune cells, namely, CD8+ T cell markers (CD8A, CD8B), T cell (general) markers (CD3D, CD3E, CD2), B cell markers (CD19, CD79A), monocyte markers (CD86, CD115), TAM markers (CCL2, CD68, IL10), M1 Macrophage marker (iNOS, IRF5), M2 Macrophage markers (CD163, VSIG4, MS4A4A), neutrophils markers (CD11b, CCR7), NK cell markers (KIR2DL1, KIR2DL3, KIR2DL4, KIR3DL1, KIR3DL2, KIR2DS4), DC markers (HLA-DPB1, HLA-DQB1, HLA-DRA, HLA-DPA1, BDCA-1, BDCA-4, CD11c), Th1 cell markers (T-bet, STAT4, IFN-γ, TNF-α, STAT1), Th2 cell markers (GATA3, STAT6, STAT5A, IL13), and Tfh cell marker (BCL6).
The FMNL2 mRNA level showed significantly positive correlation with the expression of markers of specific immune cells, namely, CD8+ T cell markers (CD8A, CD8B), T cell (general) markers (CD3D, CD3E, CD2), B cell marker (CD19), monocyte markers (CD86, CD115), TAM markers (CCL2, CD68, IL10),M1 macrophage markers (IRF5, COX2), M2 macrophage markers (CD163, VSIG4, MS4A4A), neutrophils markers (CD11b, CCR7), NK cell markers (KIR2DL1, KIR2DL3, KIR3DL1, KIR3DL2, KIR2DS4), DC markers (HLA-DPB1, HLA-DQB1, HLA-DRA, HLA-DPA1, BDCA-1, BDCA-4, CD11c), Th1 cell markers (T-bet, STAT4, IFN-γ, STAT1), Th2 cell marker (GATA3, SATA6), and Tfh cell marker (BCL6).
The FMNL3 mRNA level showed significantly positive correlation with the expression of markers of specific immune cells, namely, CD8+ T cell markers (CD8A, CD8B), T cell (general) markers (CD3D, CD3E, CD2), B cell markers (CD19, CD79A), monocyte markers (CD86, CD115), TAM markers (CCL2, CD68, IL10), M1 Macrophage markers (iNOS, IRF5, COX2), M2 macrophage markers (CD163, VSIG4, MS4A4A), neutrophils markers (CD11b, CCR7), NK cell markers (KIR2DL1, KIR2DL3, KIR3DL1, KIR3DL2, KIR2DS4), DC markers (HLA-DPB1, HLA-DQB1, HLA-DRA, HLA-DPA1, BDCA-1, BDCA-4, CD11c), Th1 cell markers (T-bet, STAT4, IFN-γ, TNF-α, STAT1), Th2 cell markers (GATA3, STAT6, STAT5A, IL13), and Tfh cell marker (BCL6).
PPI network of FMNLs in GC and enrichment analysis
Next, to explore the potential mechanisms that FMNLs participate in the carcinogenesis of GC, we used GeneMANIA to construct a PPI network for FMNLs, and the result is shown in Figure 5.
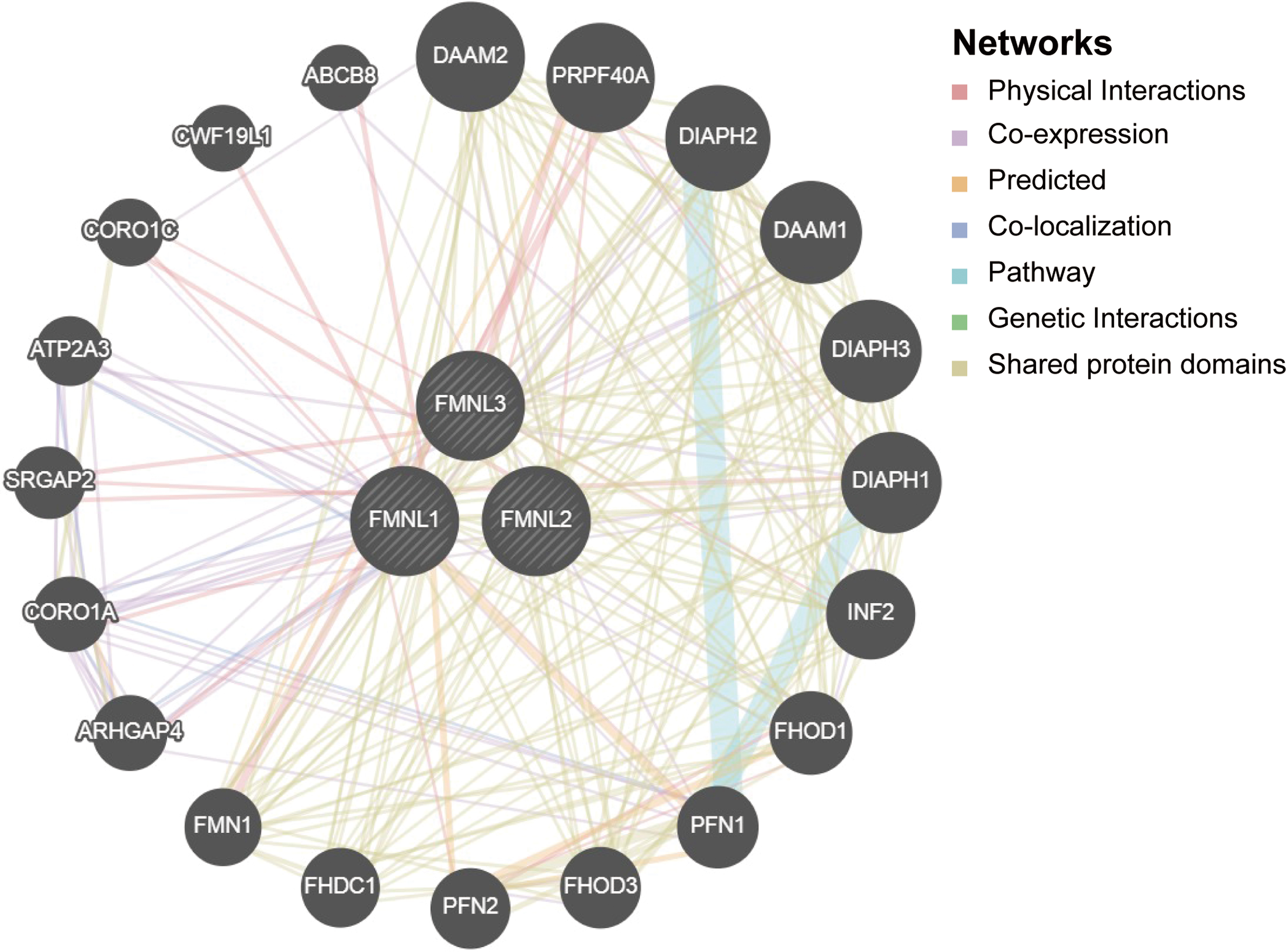
Predicted pathways of FMNLs in GC. PPI network for FMNLs was constructed in GeneMANIA. The interconnections between proteins were explored in terms of physical interaction, coexpression, predicted, colocalization, common pathway, genetic interaction, and shared protein domains. PPI, protein–protein interaction. Color images are available online.
Then, GO and KEGG analyses based on DAVID were performed to identify the functional enrichment of FMNLs and their associated genes (Table 4). Biological processes (BP) enrichment term exhibited that FMNLs and their cooperators were significantly associated with actin cytoskeleton organization, cytoskeleton organization, regulation of cell shape, negative regulation of actin filament polymerization, and positive regulation of stress fiber assembly; cellular components (CC) enrichment term suggested that FMNLs were located in cytosol; molecular functions (MF) enrichment showed that FMNLs were significantly correlated to binding Rho GTPase, actin, Rac GTPase, and actin monomer; KEGG enrichment revealed that FMNLs were related to regulation of actin cytoskeleton and shigellosis. Overall, FMNLs-coexpressed genes mainly participate in organizing actin cytoskeleton, which may affect small G proteins, including Rho proteins and Rac proteins.
GO and KEGG Enrichment Analysis of FMNLs and Their 20 Interactors
CC, cellular component; FDR, false discovery rates; GO, gene ontology; KEGG, Kyoto Encyclopedia of Genes and Genomes; MF, molecular functions.
Moreover, results from the LinkedOmics database revealed that FMNLs participate in many immune processes. Table 5 shows the top 5 most highly enriched GO and KEGG terms. High expression of FMNL1 was associated with interleukin-2 production, respiratory burst, interleukin-4 production, interleukin-10 production, mast cell activation (BP category), MHC protein complex, immunological synapse, mast cell granule, protein complex involved in cell adhesion, side of membrane (CC category), coreceptor activity, nucleotide receptor activity, MHC protein binding, cytokine receptor activity, purinergic receptor activity (MF category), hematopoietic cell lineage, Staphylococcus aureus infection, asthma, Leishmaniasis, and intestinal immune network for IgA production (KEGG category).
Top 5 Terms of GO and KEGG Analysis by GSEA
CAM, cell adhesion molecule; ECM, extracellular matrix; ES, enrichment score; GSEA, Gene Set Enrichment Analysis.
High expression of FMNL2 was associated with hippo signaling, adaptive immune response, negative regulation of cell activation, B cell activation, negative chemotaxis (BP category), protein complex involved in cell adhesion, inclusion body, DNA repair complex, receptor complex, microtubule organizing center part (CC category), phosphatidylinositol 3-kinase activity, transmembrane receptor protein kinase activity, helicase activity, coreceptor activity, phosphatidylinositol bisphosphate kinase activity (MF category), measles, MicroRNAs in cancer, JAK-STAT signaling pathway, extracellular matrix (ECM)-receptor interaction, and hematopoietic cell lineage (KEGG category).
High expression of FMNL3 was associated with substrate-dependent cell migration, integrin activation, mast cell activation, cellular response to vascular endothelial growth factor stimulus, regulation of lipase activity (BP category), protein complex involved in cell adhesion, collagen trimer, mast cell granule, ECM, phagocytic cup (CC category), ECM structural constituent, coreceptor activity, cytokine binding, proteoglycan binding, cytokine receptor activity (MF category), ECM/receptor interaction, malaria, hematopoietic cell lineage, primary immunodeficiency and cell adhesion molecules (KEGG category).
Discussion
FMNLs are associated with poor prognosis in a variety of cancers (Higa et al., 2019),(Gardberg et al., 2016), (Liu et al., 2019). However, there are few studies on the expression of FMNLs in GC and their exact effects on prognosis. In this study, we analyzed the expression of FMNLs in GC, correlation with prognosis and immune infiltrating. The results suggest that high FMNL1 and FMNL3 mRNA levels were correlated with poor prognosis of GC. Furthermore, FMNLs mRNA levels correlated with the abundance of tumor-infiltrated immune cells based on the levels of markers for different immune cell types in GC. Overall, our research provided a novel insight into the critical roles of FMNLs in prognostic assessment and immune infiltrating in GC.
FMNL1, FMNL2, and FMNL3 are members of the formin proteins family (Wang et al., 2015). FMNL1 mediates the assembly of filamentous actin networks and organizes actin filaments into bundles. FMNL1 also stabilizes the Golgi complex and regulates phagocytosis, cell adhesion, podosome dynamics, cell migration, and survival in macrophages (Higa et al., 2019). It has been found that high FMNL1 expression is correlated with an unfavorable prognosis in several cancers, including glioblastoma multiforme (Higa et al., 2019), nasopharyngeal carcinoma (Chen et al., 2018), and non-small-cell lung cancer (Yang et al., 2019). It has been proved that high FMNL1 level plays an important role in regulating proliferation, migration, and invasion (Yang et al., 2019).
In this report, we found that the mRNA level of FMNL1 was higher in GC than normal. According to individual cancer stages and tumor grades, the mRNA level of FMNL1 increased in advanced and poorly differentiated GC. As to prognosis, high FMNL1 expression indicated shorter OS.
FMNL2 acts as effectors and upstream modulators of Rho GTPase signaling and control the actin-dependent processes, such as cell motility or invasion (Zeng et al., 2015a). It has been proved that FMNL2 plays a role in cell motility, invasion, and metastasis of colorectal carcinoma (Zhu et al., 2011). Several miRNAs, including miR-34a (Lu et al., 2015), miR-137 (Liang et al., 2013), and miR-206 (Ren et al., 2016) might function as tumor suppressors in the progression of colorectal carcinoma by regulating FMNL2. On the contrary, downregulation of FMNL2 predicts poor prognosis in hepatocellular carcinoma (Liang et al., 2011). In GC, it has been shown that silencing FMNL2 suppressed proliferation, migration, and invasion, and induced apoptosis of GC cells. The integrin internalization induced by PKC was declined by FMNL2 silencing (Zhong et al., 2018). Our results conformed the tumor-promoting role of FMNL2 in GC based on the evidence from gene expression.
In this research, we found that the FMNL2 expression was higher in GC than normal and high FMNL2 mRNA level was associated with poor prognosis in specific GC subtype, including clinical stage 1 and diffuse GC.
FMNL3 plays a crucial role in cytoskeletal mediation and is potentially a biomarker for cell migration (Wu et al., 2017). FMNL3 functions in assembly of actin-based protrusions that are specialized for cell-cell adhesion (Gauvin et al., 2015). High expression of FMNL3 leads to metastasis and poor prognosis in cancers such as colorectal cancer (Zeng et al., 2015b), tongue squamous cell carcinoma (Liu et al., 2019), and nasopharyngeal carcinoma (Wu et al., 2017). In colorectal cancer, FMNL3 might overregulate the Rhoc/FAK pathway and actin assembly to promote the invasion of colorectal cancer cells (Zeng et al., 2018). Our current study also demonstrated that FMNL3 expression was higher in GC than normal and related with poor prognosis, suggesting its oncogenic role in GC.
In recent years, the importance of immune cells infiltrating in tumor gradually comes to be recognized. As early as in 1968 and 1976, Hamlin et al. and Bertrand et al. reported the relationship between immune infiltration and prognosis of breast cancer, respectively (Hamlin, 1968; Husby et al., 1976). Immune cell infiltration is associated with clinical outcomes in a variety of tumors such as colorectal cancer (Xiong et al., 2018), melanoma (Dabrosin et al., 2019), and non-small-cell lung cancer (Edlund et al., 2019).
Tumor-specific cytotoxic T cells exist in GC tissues but cannot inhibit tumor growth due to poor immune response in the tumor microenvironment (He et al., 2017). Soluble LAG3 acts as a potential prognostic marker of GC and its positive correlation with CD8+ T cell frequency and secretion of IL-12 and INF-γ in peripheral blood (Li et al., 2018). Tumor antigen-specific CD8+ T cells might be negatively regulated by PD-1 and Tim-3 in human GC (Lu et al., 2017). Moreover, immune checkpoint blocking has become a promising treatment for cancer (Wang et al., 2018).
However, the relationship between FMNLs and immune infiltrating in GC is unstudied. In this article, the relationship between the expression of FMNLs and immune infiltrating in GC was analyzed using the TIMER database. The expressions of FMNL1/3 were significantly associated with various immune infiltrating cells. The FMNL2 mRNA level was also significantly positively correlated with the expression of markers of specific immune cells. The correlation between the expressions of FMNLs and these gene markers suggested that FMNLs play prominent roles in regulating tumor immune cell infiltrating.
To explore the potential mechanisms that FMNLs participate in the carcinogenesis of GC, we constructed a PPI network and performed GO and KEGG analyses for FMNLs based on DAVID. The results show that FMNLs co-expression genes mainly participate in organizing actin cytoskeleton, which may affect small G proteins, including Rho proteins and Rac proteins. Rho GTPase has been reported to interact with DRFs, FMNLs, and DAAMs (Kuhn and Geyer, 2014; Arnold et al., 2017; Basant and Glotzer, 2018; Yan et al., 2018; Mei et al., 2019). Moreover, ROCK (Rho-associated coiled-coil kinase) is the downstream of RhoA and plays an important role in the interaction between formins and Rho (Kuhn and Geyer, 2014). ROCK blockade might enhance cancer cell phagocytosis as well as DC-mediated T cell priming to elicit anticancer immunity (Nam et al., 2018).
In a word, the interaction between RhoA/ROCK pathway and FMNLs may become a novel antitumor therapy strategy by regulating immune infiltrating. Besides, enrichment analysis revealed that FMNLs and their coexpressed genes were tightly associated with immune-related terms. These results provided novel insights into the FMNLs-mediated immunoregulation in GC.
Conclusion
In conclusion, the mRNA levels of FMNLs are higher in GC compared with normal tissues and associated with poor prognosis and the level of immune cell infiltrating. Therefore, FMNLs might play an immunomodulatory role in tumor immunity and may be new therapeutic targets and potential prognostic biomarkers in GC.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Natural Science Foundation of China (81773227), the Young Medical Talents of Wuxi (QNRC036).