
Abstract
This study aimed at exploring the role of EgRad54 and the effect of harmine (HM) or HM derivatives (HMDs) on DNA damage in Echinococcus granulosus. DNA damage in E. granulosus protoscoleces (PSCs) was assessed by using a comet assay, after treatment with HM or HMDs. Efficiency of electroporation-based transfection of PSCs and subsequent EgRad54 knockdown was evaluated by using real-time quantitative polymerase chain reaction (RT-qPCR) and fluorescence intensity. Viability of PSCs was determined via eosin exclusion test, and expression of related genes was analyzed via RT-qPCR. HM and HMDs significantly (p < 0.05) increased DNA damage in E. granulosus, and upregulated EgRad54 expression. Compared with HM and HMD-only treatment groups, EgRad54 knockdown combined with HM and HMD treatment further reduced E. granulosus viability. This combined approach resulted in significant (p < 0.05) downregulation of Rad54 and Topo2a expression, and upregulation of ATM expression, whereas H2A and P53 expression was significantly higher compared with control groups. These data show that EgRad54 knockdown, combined with HM or HMD treatment, enhances DNA damage in E. granulosus via upregulation of ATM and H2A, and downregulation of Rad54 and Topo2a, thereby inhibiting E. granulosus growth, and suggest that EgRad54 is a potential therapeutic target for cystic echinococcosis treatment.
Introduction
Echinococcosis is a zoonotic parasitic disease caused by Echinococcus larvae, and ∼95% of cystic echinococcosis (CE) is caused by Echinococcus granulosus (Agudelo Higuita et al., 2016; Deplazes et al., 2017). CE has become a serious global public health problem (Budke et al., 2006), with human incidence exceeding 50 cases per 100,000 person-years in endemic areas; in parts of China, the prevalence rate is as high as 5–10% (Craig et al., 2007). Currently, surgical treatment and drug therapy are applied to treat CE, with surgical treatment mainly being used to treat patients with low tomont numbers and early CE (Agudelo Higuita et al., 2016). Due to the complexity of CE, the postoperative recurrence rate is high. Albendazole is the preferred drug for the treatment of CE (Brunetti et al., 2010), but it can cause adverse side effects and drug resistance, and it does not completely cure CE. Therefore, there is an urgent need to develop novel therapeutic drugs and identify novel therapeutic targets for the prevention and treatment of CE.
Harmine (HM), a natural β-carboline alkaloid found in the seeds of Peganum harmala L., has a series of pharmacological properties, including antibacterial, anti-plasmodium, anti-inflammatory, anti-tumor, anti-oxidation, and neuroprotective properties (Dos Santos and Hallak, 2017; Chen et al., 2018; Ding et al., 2019). Dos Santos and Hallak (2017) indicated that HM exerts neuroprotective and cognition-enhancing effects by decreasing reactive oxygen species (ROS), and increasing neurotrophic and anti-inflammatory factors. Another study showed that HM induced DNA damage and was cytotoxic in V79 Chinese hamster lung fibroblasts (Boeira et al., 2001). However, the effects of HM on CE, and on DNA damage in E. granulosus are unclear.
A previous study indicated that in mammalian cells, the outcome of DNA damage varies and is usually adverse, and that the acute effects of disturbed DNA metabolism can lead to cell cycle arrest and cell death (Hoeijmakers, 2001). To maintain genome stability, cells can mitigate DNA damage by transmitting DNA damage signals to specific effectors, triggering DNA repair, damage tolerance, cell cycle checkpoints, and cell death pathways (Chatterjee and Walker, 2017). The DNA repair and recombination proteins, RAD54 and RAD54-like protein, belong to the SWI2/SNF2 protein family that regulate protein-DNA interactions in an ATP-driven manner (Solinger et al., 2001). Rad54 reportedly participates in a number of DNA repair pathways (Tan et al., 1999). A study by Hirakawa et al. (2017) showed that in Arabidopsis thaliana Rad54 accumulated at sites of DNA damage and formed DNA repair foci in response to DNA damage. In addition, Rad54 is a key homologous recombination (HR) factor, essential for rapid response to post-replicative stress and DNA damage. Choi et al. (2017) demonstrated that proliferation, cell cycle progression, and viability of HeLa cells are closely related to the abundance of HR factors. However, the roles of Rad54 in E. granulosus are still unknown.
Therefore, in this study, E. granulosus protoscoleces (PSCs) were treated with HM or HM derivatives (HMDs), and the effects of these treatments on DNA damage in E. granulosus were determined. Further, E. granulosus Rad54 knockdown was achieved through an electroporation-based method, and the effects of Rad54, combined with HM or HMDs treatment on DNA damage in E. granulosus were explored. The results provide a crucial theoretical basis for new therapeutic targets and drugs for the prevention and treatment of CE.
Materials and Methods
E. granulosus culture
E. granulosus PSCs were isolated from liver cysts of sheep infected with E. granulosus, at Hualing slaughter house (Urumqi, Xinjiang). E. granulosus does not belong to experimental animals, and is a kind of parasite. It is not necessary to provide IRB approval.
Briefly, the liver cyst fluid was extracted under aseptic conditions and transferred to a sterile container. The fluid was allowed to stand to allow E. granulosus PSCs to sediment, after which the supernatant was discarded. Subsequently, PSCs were washed with 0.9% saline containing penicillin (v/v 1:100), and then 2 mL of the protoscolex-containing solution was added to 50 mL 1% pepsin. After pepsin digestion at 37°C for 30 min, the activated PSCs were washed by using 0.9% saline containing penicillin. After filtration, isolated PSCs were cultured in RPMI 1640 medium (HyClone, Logan, UT) supplemented with 10% fetal bovine serum (MRC Global, Inc., Houston, TX), and 1% penicillin (HyClone), and maintained in an incubator with 5% CO2 at 37°C. The culture medium was changed every 4–5 days.
Determination of DNA damage and Rad54 expression levels
The effects of HM and HMDs (DH-330 and DH-004) on DNA damage in E. granulosus were determined by using comet assays, based on the method of Lu et al. (2017) with minor modifications. Briefly, E. granulosus PSCs (2000/well) were seeded into a 96-well plate, which was divided into five groups: blank control group, negative control (NC) group, HM group, DH-330 group, and DH-004 group. The PSCs in the HM, DH-330, and DH-004 groups were treated with 100 μM HM, 100 μM DH-330, and 100 μM DH-004, respectively. The PSCs in the blank control group and NC group were treated with the same quantity of PBS and 1% dimethyl sulfoxide (DMSO; AMRESCO, Inc., Framingham, MA). HM, DH-330, and DH-004 were provided by Xinjiang Huashidan Pharmaceutical Co. Ltd (Urumqi, Xinjiang). After incubation for 2 days, the cells were washed with PBS, resuspended in 0.75% low-density agarose (100 μL), and finally fixed on slides. After solidification, the slides were placed in lysis solution for 4 h. After washing, the slides were placed in an electrophoresis chamber with alkaline electrophoresis solution and electrophoresed for 30 min (25 V, 300 mA). Subsequently, the slides were stained with propidium iodide (Beijing Baishayi Technology Co. Ltd, Beijing, China) in the dark for 20 min, and then de-colored by using ultrapure water for 5 min. Finally, the slides were observed under an inverted fluorescence microscope (Nikon, Japan) and images were taken. The Olive Tail Moment (OTM) was analyzed by using CASP software (version 1.2.3). Total RNA was extracted from PSCs from different treatment groups, and a real-time quantitative polymerase chain reaction (RT-qPCR) was used to examine the expression level of Rad54. The primer sequence of Rad54 is shown in Table 1.
Sequences of All Primers
SiRNA interference and electrotransfection efficiency assay
SiRNA-NC and siRNA-EgRad54-210, -772, and -1205 (labeled with green fluorescent modification) were designed and provided by Shanghai GenePharma Co., Ltd. (Shanghai, China). The sequences of these siRNAs are shown in Table 1. Transfection of PSCs was performed by using an electroporation-based method, based on the study of Han (Han, 2018). Briefly, E. granulosus PSCs were cultured in normal medium (detailed above) for 1 day, and ∼4000 PSCs were added to electroporation buffer (200 μL). Afterward, siRNA-NC and siRNA-EgRad54-210, -772, or -1205 were added at the final concentration of 5 μM. After electric shock at 125 V for 20 ms, the shock cup was quickly placed in an incubator at 37°C for 10 min. The transfected PSCs were transferred to 3 mL normal medium and cultured for another 3 days. The PSCs in the siRNA-EgRad54-210, -772, and -1205 groups were then treated with HM (100 μM) for 1 day, and the total RNA was isolated from the PSCs in different groups. The transfection efficiency was evaluated by determining the expression level of Rad54 using RT-qPCR.
The optimal siRNA obtained from the procedure mentioned earlier was selected for E. granulosus transfection. After transfection for 1 h, ∼250 PSCs were collected in each group, washed with PBS, and placed onto microscope slides. Fluorescence images were acquired by using an inverted fluorescence microscope, and green fluorescence intensity in each group was observed.
Viability of E. granulosus PSCs treated with siRNA, HM, or HMDs
E. granulosus PSCs were cultured in the normal medium for 1 day and divided into eight groups: blank control, siRNA-NC, HM, HM+siRNA-EgRad54, DH-330, DH-330+siRNA-EgRad54, DH-004, and DH-004+siRNA-EgRad54 groups. The PSCs in the HM+siRNA-EgRad54, DH-330+siRNA-EgRad54, and DH-004+siRNA-EgRad54 groups were first transfected with siRNA-EgRad54 for 3 days and then treated with 100 μM HM, 100 μM DH-330, or 100 μM DH-004, respectively. The PSCs in the NC group were transfected with siRNA-NC and treated with PBS. In addition, PSCs in the blank control, HM, DH-330, and DH-004 groups were treated only with PBS, 100 μM HM, 100 μM DH-330, or 100 μM DH-004, respectively. After 1 day treatment, PSCs in each group were washed three times with PBS and stained with 1% eosin dye solution for 2 min. An inverted fluorescence microscope was used to observe and calculate the viability of PSCs.
Real-time quantitative polymerase chain reaction
Total RNA was extracted from the PSCs in different groups by using TRIzol reagent (Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer's instructions, and total RNA concentrations were measured by using a microplate reader. Total RNA was reverse-transcribed into cDNA by using a PrimeScript™ RT Reagent Kit (Takara, Beijing, China). All primers were designed and synthesized by Sangon Biotech (Shanghai) Co., Ltd (Shanghai, China), and they are shown in Table 1. The reaction volume of RT-qPCRs was 20 μL, which included: 10 μL Ex-Taq™II, 0.8 μL forward primer, 0.8 μL reverse primer, 0.4 μL ROX, 2 μL cDNA, and 6 μL double-distilled water. RT-qPCRs were initiated at 95°C for 30 s, followed by 95°C for 5 s, and 60°C for 30 s, for 40 cycles. The melting curve was as follows: 95°C for 15 s, 65°C for 15 s, and 95°C for 15 s. β-actin served as a housekeeping gene and the relative mRNA expression of ATM serine/threonine kinase (ATM), tumor suppressor p53-binding protein (P53), topoisomerase 2A (Topo2a), histone H2A (H2A), and EgRad54 in E. granulosus PSCs was calculated by using the 2−ΔΔCt method (Xu et al., 2019).
Statistical analysis
Data are expressed as mean ± standard deviation. SPSS 22.0 statistical software was used to perform all the statistical analyses. Comparisons between two groups were analyzed by using t-tests, and the comparison between multiple groups was analyzed by using one-way analysis of variance. p < 0.05 indicated that the difference was statistically significant.
Results
The effects of HM and HMDs on DNA damage
DNA damage in E. granulosus PSCs treated with different HM or HMDs was evaluated by using comet assays. The PSCs in the blank control group and NC group showed complete and compact nuclei without tailing (Fig. 1A). However, DNA fragments from PSCs in the HM, DH-330, and DH-004 groups migrated from the nucleus to the anode, and they formed a tail under the same electrophoretic conditions (Fig. 1A). In addition, OTM was analyzed in different groups. There was no significant difference in OTM values between the blank control group (0.013 ± 0.029) and NC group (0.047 ± 0.103) (p > 0.05, Fig. 1B), which indicated that DMSO had no effect on DNA damage in E. granulosus. The OTM values in the HM, DH-330, and DH-004 groups were 24.052 ± 5.311, 47.258 ± 4.383, and 61.879 ± 3.125, respectively, which showed a significant increase compared with the control groups (p < 0.05, Fig. 1B). These results suggest that HM and HMDs exacerbate DNA damage in E. granulosus.

The effects of HM and HMDs on DNA damage in Echinococcus granulosus.
Rad54 expression in different treatment groups
The effects of Rad54, a functional gene associated with DNA damage repair, on DNA damage in E. granulosus were investigated. Relative expression of Rad54 in the blank control, NC, and DH-004 groups was 1.01 ± 0.24, 1.11 ± 0.13, and 1.84 ± 0.19, respectively (Fig. 2). The results show that there was no significant difference in Rad54 expression among the blank control, NC, and DH-004 groups. Compared with the control group, relative expression of Rad54 in the HM (7.91 ± 0.85) and DH-330 groups (2.64 ± 0.40) was markedly increased (p < 0.05), and HM had a greater effect on Rad54 expression compared with the DH-330 group (p < 0.05, Fig. 2). These results indicate that HM and HMDs (except DH-004) upregulate Rad54 in E. granulosus.
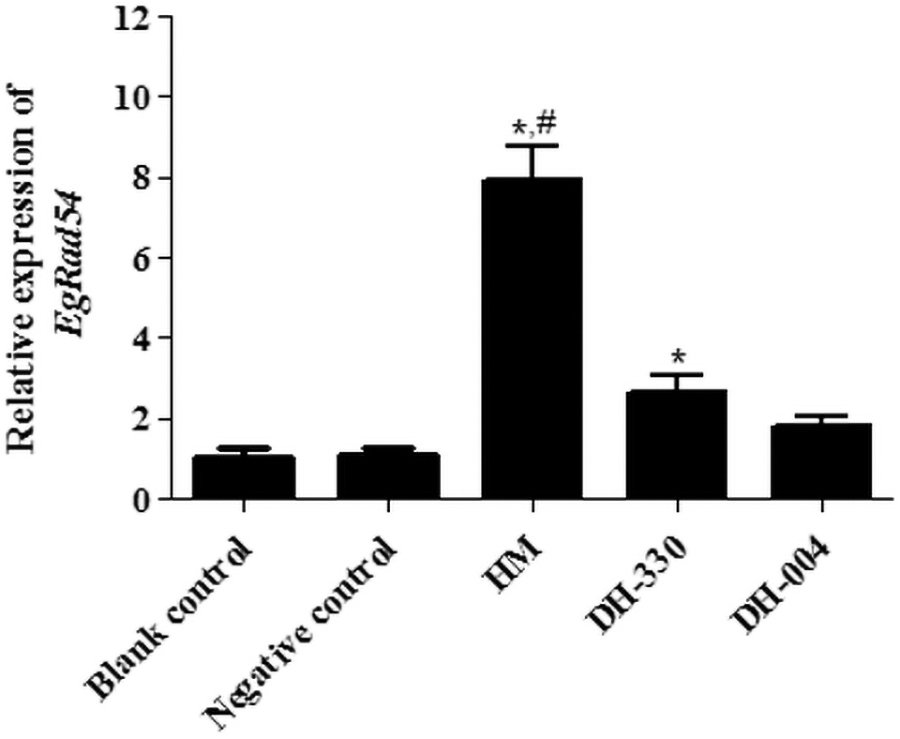
Relative expression of EgRad54 in different treatment groups determined by using RT-qPCR. The symbol “*” indicate statistically significant (p < 0.05) differences from the blank control group, whereas statistically significant (p < 0.05) differences from the DH-330 treatment group are indicated by the symbol “#”. RT-qPCR, real-time quantitative polymerase chain reaction.
Electrotransfection efficiency analysis
Relative expression of Rad54 in different treatment groups was measured to evaluate electrotransfection efficiency. The expression levels of Rad54 in the control and siRNA-NC groups were 1.09 ± 1.00 and 1.49 ± 1.61, respectively, which showed no significant difference (p > 0.05, Fig. 3A). After treatment with HM, the expression of Rad54 was significantly increased to 31.24 ± 2.99 compared with the control group (p < 0.05). However, compared with the HM group, Rad54 expression in the siRNA-EgRad54-210, -772, and -1205 groups was all downregulated, whereas the Rad54 expression in the siRNA-EgRad54-772 group (9.94 ± 4.59) was significantly lower than that in the HM group (p < 0.05, Fig. 3A). Thereafter, E. granulosus PSCs were transfected with siRNA-EgRad54-772 and the fluorescence intensity was observed. There were no green fluorescent spots in PSCs of the control group, whereas PSCs in the siRNA-EgRad54-722 and siRNA-NC groups exhibited bright green fluorescent spots (Fig. 3B). Based on these results siRNA-EgRad54-772 was chosen for Rad54 interference in the subsequent experiment.

Electrotransfection efficiency after siRNA interference.
Viability of E. granulosus treated with HM or HMDs
Images of eosin staining are shown in Figure 4A. There was no significant difference in E. granulosus viability between the control (99.9% ± 0.26%) and siRNA-NC groups (99.1% ± 0.32%, p > 0.05, Fig. 4B). Compared with the control group, the viability of E. granulosus was significantly reduced in the HM (86.51% ± 2.09%), DH-330 (36.20% ± 5.28%), and DH-004 groups (28.87% ± 2.48%, p < 0.05, Fig. 4B). The viability of E. granulosus in the HM+siRNA-EgRad54, DH-330+siRNA-EgRad54, and DH-004+siRNA-EgRad54 groups was 30.11% ± 4.6%, 8.66% ± 3.75%, and 7.71% ± 1.92%, respectively. These data show that when siRNA-EgRad54 interference was combined with HM or HMD treatment, viability of E. granulosus was decreased even more compared with HM or HMD treatments only (p < 0.05, Fig. 4B). Further, these data show that EgRad54 knockdown reduced the tolerance of E. granulosus to HM and HMDs.
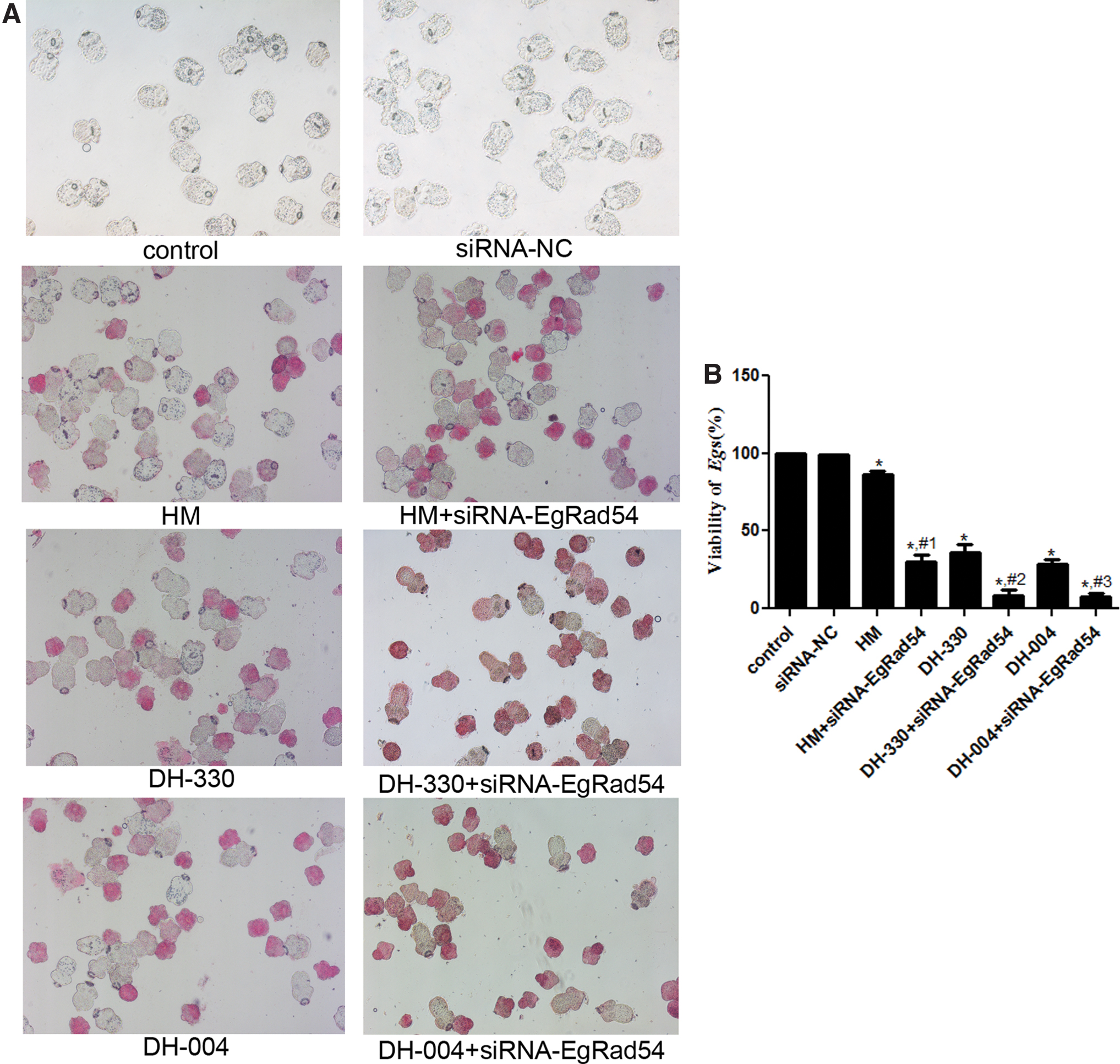
The effects of siRNA-EgRad54 combined with HM or HMDs treatment on viability of E. granulosus.
RT-qPCR analysis
To explore how EgRad54 knockdown in combination with HM or HMDs affects DNA damage, RT-qPCR was carried out to study the mRNA expression levels of ATM, P53, Topo2a, H2A, and Rad54. There were no significant differences between the blank control and siRNA-NC groups in the expression levels of all genes (p > 0.05, Fig. 5). In addition, compared with the blank control group, the expression of both Rad54 and Topo2a was markedly upregulated (p < 0.05); whereas Rad54 and Topo2a expression was significantly downregulated after EgRad54 knockdown compared with treatment groups that received only HM or HMDs (p < 0.05, Fig. 5A, B). The expression of ATM was notably higher after HM and HMD treatment compared with that in the blank control group (p < 0.05), and it was further upregulated after EgRad54 knockdown (p < 0.05, Fig. 5C). Interestingly, there was no significant difference in H2A expression among the blank control, HM, DH-330, and DH-004 groups (p > 0.05), whereas H2A expression was significantly increased after EgRad54 knockdown compared with that in the treatment groups that only received HM or HMDs (p > 0.05, Fig. 5D). Further, P53 expression was significantly upregulated in the treatment groups that only received HM or HMDs compared with the blank control group (p < 0.05). EgRad54 knockdown had no significant effect on P53 expression compared with expression in the treatment groups that only received HM or HMDs (p > 0.05, Fig. 5E).
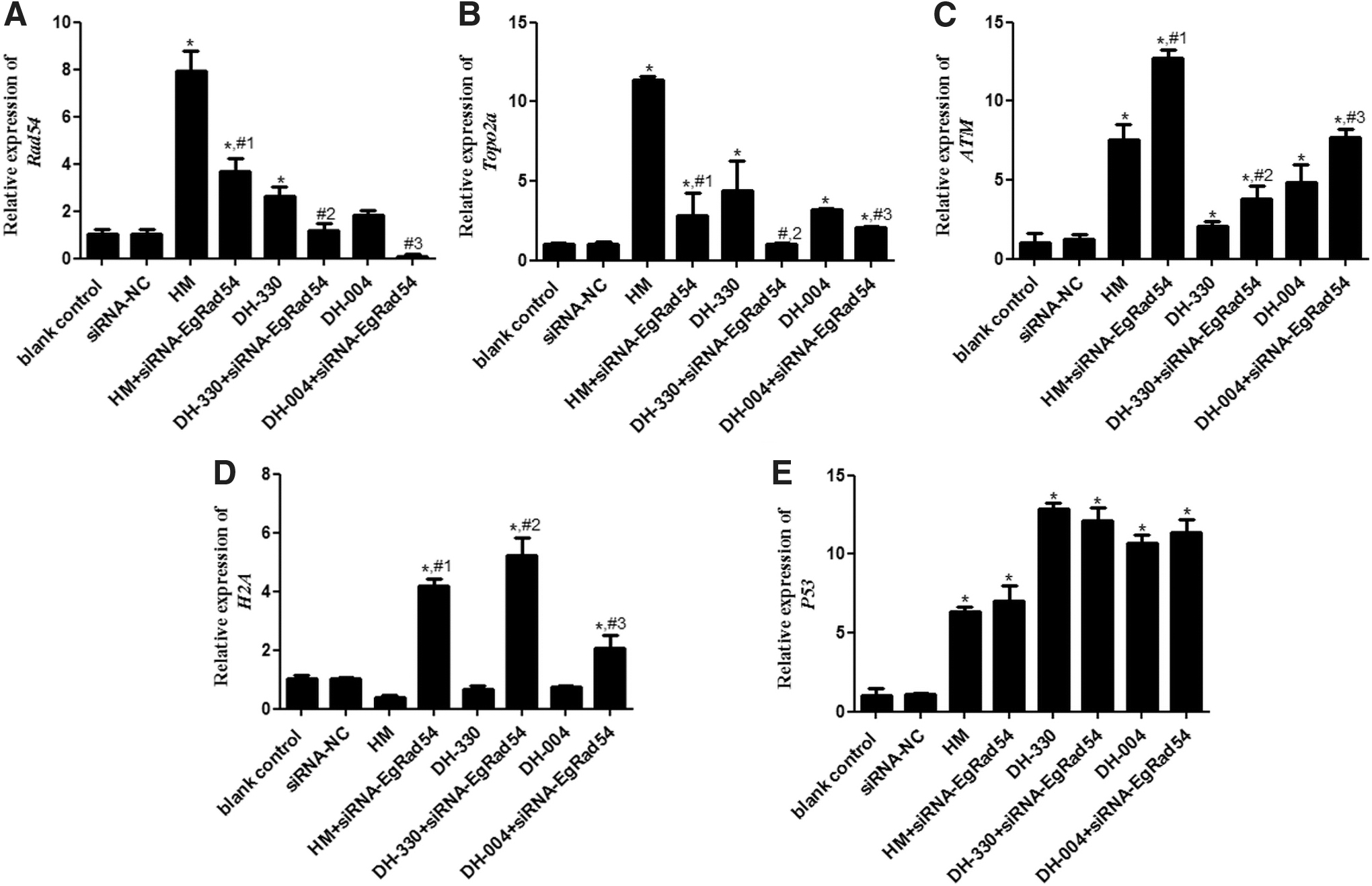
Relative expression of
Discussion
CE is a well-known but neglected parasitic disease caused by E. granulosus, and it seriously affects human health, leading to substantial economic losses (McManus et al., 2003). HM, an inhibitor of HR, reportedly exerts anti-tumor effects by causing DNA damage. Further, HM has many traditional medicinal uses, and has pharmacological activities, such as anti-parasitic, anti-cancer, and anti-inflammatory properties (Zhang et al., 2015). In addition, previous studies have demonstrated that HM inhibits cell proliferation, migration, and invasion of various tumor cells, and it induces apoptosis (Himpel et al., 2001; Uhl et al., 2018). Our study found that HM and HMDs exacerbated DNA damage in E. granulosus and upregulated the expression of EgRad54, thereby reducing E. granulosus viability. A study by Di Giorgio et al. (2004) indicated that the anti-proliferation property of HM and harmane, which inhibited the growth of Leishmania infantum, was associated with DNA metabolism and S-G2 cell cycle phases. Another study showed that HM and two other β-carboline alkaloids suppressed invasion and growth of Toxoplasma gondii via DNA replication interference during cellular growth (Alomar et al., 2013). This, together with data from the current study, suggests that HM and HMDs might inhibit the growth of E. granulosus through DNA damage, thus improving CE.
In this study, HM and HMDs upregulated Rad54 expression. Rad54 is associated with DNA damage repair (Hirakawa et al., 2017), but there are a few reports on the effects of Rad54 in E. granulosus. We hypothesized that Rad54 knockdown could promote HM- or HMD-induced DNA damage in E. granulosus, thereby ameliorating CE caused by E. granulosus infection. To test this hypothesis, EgRad54 knockdown was successfully achieved in E. granulosus via an electroporation-based method, after which E. granulosus PSCs were treated with HM or HMDs. Rad54 is a potent, progressive motor protein that transposes on dsDNA, and it may have multiple functions in recombinant DNA repair (Ceballos and Heyer, 2011). Our research, to the best of our knowledge, has demonstrated for the first time that EgRad54 knockdown, in conjunction with HM or HMD treatment, further suppresses the viability of E. granulosus, compared with HM or HMD treatment alone. This implies that Rad54 might be a potential target for CE treatment, and that Rad54 knockdown combined with HM or HMDs has a better inhibitory effect on E. granulosus. Peng et al. (2018) showed that fluconazole treatment led to the upregulation of Rad54 in Cryptococcus neoformans, and increased DNA damage in vitro. Another study by Cole et al. (1987) demonstrated that Rad54 in Saccharomyces cerevisiae participated in both DNA repair and recombination, and it was inducible by X-rays. Therefore, we speculated that in E. granulosus, Rad54 could serve as a potential target of DNA damage, and that Rad54 knockdown could suppress the growth of E. granulosus by exacerbating DNA damage.
To better understand the molecular mechanisms underlying the observed effects of Rad54 knockdown combined with HM or HMD treatment on DNA damage in E. granulosus, RT-qPCR was used to investigate the expression of Rad54, Topo2a, ATM, H2A, and P53. EgRad54 knockdown combined with HM or HMDs further downregulated the expression of Rad54 and Topo2a, whereas it further upregulated the expression of ATM, and increased H2A expression, compared with HM or HMD treatment alone. There was no significant difference in P53 expression between HM and HMD treatment alone and that observed when treatment was combined with EgRad54 knockdown. During the DNA damage process, ATM, an important sensing protein, can quickly recognize DNA damage signals (Nanda and Roberts, 2020). Activated ATM not only specifically identifies DNA damage but also recruits, via phosphorylation, various downstream proteins, including P53, Topo2a, and H2A, to the damaged site for repair (Yamamoto et al., 2016; Legina et al., 2020; Nanda and Roberts, 2020). P53 usually functions in cell division and monitors the extent of DNA damage; if the damage is minor, P53 would induce cellular DNA repair pathways and if DNA damage is extensive, P53 would induce cell apoptosis (Yeo et al., 2016). Topo2a, a major downstream effector protein of ATM, reportedly regulates the superhelix structure of DNA and plays an important role in DNA damage repair and gene transcription regulation (Ligia Cebotaru et al., 2016). Based on this and the findings from the current study, we speculate that EgRad54 knockdown combined with HM or HMD treatment can further promote DNA damage in E. granulosus via downregulating Rad54 and Topo2a expression, and upregulating ATM expression, thus further reducing E. granulosus viability.
H2A is a member of the histone family, and its phosphorylation product, γH2AX, is an important biomarker, specifically for DNA damage (Kopp et al., 2019). A study by Stewart-Ornstein and Lahav (2017) indicated that P53 expression in response to DNA damage varied across cell lines and was influenced by activity and DNA repair efficiency of ATM. Another study showed that KML001 (a telomere-targeting drug) increased the level of γH2AX, and inhibited Topo2a and ATM activity, compromising the telomere integrity of T cells (Cao et al., 2019). In the current study, HM and HMDs did not alter the expression of H2A compared with the blank control group; however, H2A expression was significantly upregulated after EgRad54 knockdown. Considering these data, we speculated that EgRad54 knockdown, combined with HM or HMD treatment may enhance DNA damage in E. granulosus by upregulating H2A expression. However, the specific modes of action of these genes require further verification through gain- and loss-of-function tests.
Conclusion
In conclusion, EgRad54 may be closely related to DNA damage in E. granulosus, and it could be a potential therapeutic target for the treatment of CE caused by E. granulosus infection. In addition, EgRad54 knockdown combined with HM or HMD treatment could further enhance DNA damage in E. granulosus through the upregulation of ATM and H2A, and the downregulation of Rad54 and Topo2a, thereby inhibiting the growth of E. granulosus and improving CE. Our findings provide new insights into the prevention and treatment of CE, with HM as a potential novel drug, and EgRad54 as a potential novel therapeutic target.
Footnotes
Authors' Contributions
Conception and design of the research: J.W. and J.Z.; acquisition of data: Y.G., C.T., and S.L.; analysis and interpretation of data: Y.G., L.W., and B.C.; statistical analysis: H.G. and H.Z.; writing—original draft preparation: Y.G. and C.T.; writing—review and editing: Y.G., C.T., J.W., and J.Z.; supervision: J.W. and J.Z. All authors have read and approved the final version.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the National Natural Science Foundation of China (Grant No. 81860666), State Key Laboratory of Pathogenesis, Prevention and Treatment of High Incidence Diseases in Central Asia, Department of Pharmacy, The First Affiliated Hospital of Xinjiang Medical University (No. SKL-HIDCA-2018-5, SKL-HIDCA-2020-1), and Natural Science Foundation of Xinjiang Uygur Autonomous Region (No. 2020D01C240).