
Abstract
Angiotensin-converting enzyme 2 (ACE2) is known as the counter-regulator of the renin–angiotensin system, it cleaves angiotensin II to render Ag 1–7, a potent vasodilator with multiple roles in cardiovascular protection. A few studies have pinpointed ACE2 polymorphisms and their relationship with heart function and hypertension in a sex-dependent manner. These observations still lack replication mostly for admixed populations. This study aimed to report minor allele frequencies of four ACE2 intron variants, rs2285666, rs2048683, rs2106809, and rs4240157, derived from previous research using the GSA, v1.0, microarray in 1231 hypertensive and nonhypertensive patients. Logistic and multiple linear regression models were developed to identify potential associations with hypertension status and systolic and diastolic blood pressure (SBP and DBP). Allele frequency differences were identified for ACE2 rs2048683 and rs4240157 in populations with European ancestry and people of the Americas. Regression analyses identified a significant association of ACE2 rs2048683 and rs4240157 with SBP/DBP in males or females. Our observations confirm sex differences in ACE2 genetic associations with SBP and DBP and contribute to the collection of genetic variation in ACE2 for admixed populations.
Introduction
Hypertension and its complications are the most common risk factors responsible for all-cause morbidity and mortality worldwide, causing over 9 million deaths per year (Oparil et al., 2018). Pharmacological control of hypertension is hindered by the incomplete understanding of its physiology, lack of useful markers, and novel interventions (Oparil and Schmieder, 2015). The renin–angiotensin system (RAS) plays a crucial role in hypertension development, but not all the proteins and enzymes involved are well characterized. The gene angiotensin-converting enzyme 2 (ACE2) was discovered only two decades ago, it maps to the short arm on the X chromosome X.p22.2, has 18 exons, and codes for a transcript of 3.4 kbp, yielding a transmembrane protein that matures into an 805-amino acid chain of 92 kDa. ACE2 belongs to the peptidase M2 family, uses Zn2+ and Cl− as cofactors, and is expressed in the intestine, heart, testis, lung, kidney, and oral epithelial cells (Tipnis et al., 2000).
ACE2 cleaves one phenylalanine from angiotensin II rendering Ag1–7, a potent vasodilator and repressor of cardiac growth known to influence heart physiology and function (Unger et al., 2015). Ag1–7 binds to the AT2 receptor (AT2R), antagonizing the effects of angiotensin II, and is thus known as the counter-arm of ACE in the RAS by opposing the effects of angiotensin II, promoting cardioprotection (Thomas et al., 2010), and browning of the white adipose tissue (Kawabe et al., 2019). ACE2 can also prevent lung damage by abating the effects of ACE in epithelial lung cells, increasing vascular permeability, pneumocyte viability, and bronchoalveolar fluid, particularly in individuals with viral respiratory infection (Nicholls and Peiris, 2005; Tikellis and Thomas, 2012). In addition to its catalytic activities, ACE2 has shown noncatalytic functions, including amino acid transport, by partnering with SLC6A19 (Hashimoto et al., 2012). More recently, ACE2 has been identified as the receptor for SARS-CoV-2 responsible for the current coronavirus disease 2019 (COVID-19) pandemic (Kuba et al., 2005). The lethality of COVID-19 has been, in part, ascribed to the high affinity of SARS-CoV-2 spike protein (S) for ACE2 followed by virus endocytosis with a wide array of tissues affected, including the pulmonary, gut, heart, renal, and the central nervous system (Oudit and Pfeffer, 2020). COVID-19 severe symptoms have been reported in patients with hypertension (6–38%) or diabetes (7.3–20%), both diseases display a deficiency of ACE2 and a decreased Ang I/Ang II ratio.
The dual role of ACE2 in cardiovascular protection and as the SARS-CoV-2 receptor provides the rationale between viral infection and cardiovascular disease. In addition, SARS-CoV-2 infection causes downregulation of ACE2 through endocytosis and proteolytic processing, hence increasing angiotensin II levels observed in COVID-19 patients, which in turn correlate with SARS-CoV-2 viral load. Heart tissues from COVID-19 disease patients show lower expression of ACE2 with an increase of myocardial inflammation and fibrosis (Oudit and Pfeffer, 2020).
ACE inhibitors and angiotensin II receptor blockers (ARBs) prescribed as antihypertensive exert their cardiovascular protective effects partly by increasing ACE2 levels; these drugs, however, can facilitate COVID-19 infection risk (Kuba et al., 2005) and lethality (Fang et al., 2020; Sungnak et al., 2020) due to the increase of ACE2.
Hypertension, type 2 diabetes, and the higher prevalence of males being infected with SARS-CoV-2 hint toward a potential role of ACE2 genetic variation influencing cardiovascular health distinctively between the sexes. ACE2 levels have been found to be 20–35% higher in men versus women, independent of antihypertensive medication (Sama et al., 2020). To date, the NCBI lists around 400 polymorphisms of ACE2, several associated with its expression, cardiovascular and respiratory diseases, digestive changes, mortality, and aging (Itoyama et al., 2005), but there is limited information on common intronic variants and their impact on hypertension in diverse world populations.
This research reports ACE2 genotype information on several hundred hypertensive and nonhypertensive patients from which we have collected genotype information from previous research using the GSA, v1.0, Illumina array that probes for four ACE2 intron variants, rs2285666, rs2048683, rs2106809, and rs4240157 (Fig. 1). We report allele frequencies of these variants in 1262 Mestizos from Mexico and their potential association with hypertension and systolic or diastolic blood pressure (SBP/DBP). Our observations provide quantitative data for ACE2 genetic variation and add to previous observations on the association between hypertension and ACE2 genetic variation in males versus females. Information on ACE2 allele frequencies should be broadened through more comprehensive genotyping or sequencing techniques to help define the role of genetic variation on ACE2 in cardioprotection, hypertension, and COVID-19 infection.
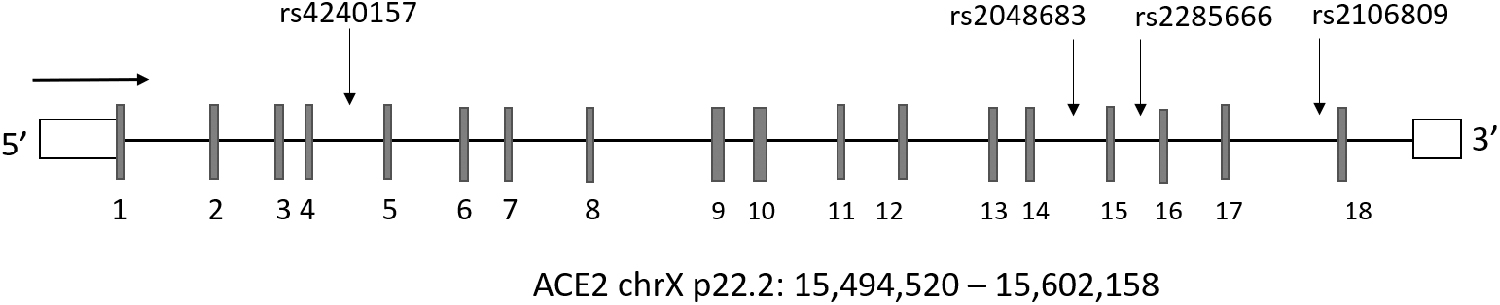
Schematic representation of the ACE2 gene structure. Footnote: introns are represented by horizontal lines and exons by vertical lines, white boxes represent the 5′ and 3′-UTRs. ACE2, angiotensin-converting enzyme 2.
Materials and Methods
Genetic data for ACE2 variants were collected from previous studies registered and approved by the Ethics and Research Committees at the National Institute of Genomic Medicine (INMEGEN-Mexico) and IMSS Centro Medico Nacional SXXI, CEI2016-23 and R-2018-785-064. Donors were from Mexico City and surrounding states, 1262 patients were recruited between 2016 and 2019, signed an informed consent, DNA was extracted from blood using the Puregene kit (Qiagen Valencia, CA) and genotyped in a GSA-24, v1.0, Illumina microarray. Hypertensive patients were under pharmacological treatment and clinical measurements, blood tests, and genomic analyses were performed under this consideration. We were able to filter four intron single-nucleotide variants (SNVs) on ACE2, rs2285666, rs2048683, rs2106809, and rs4240157, available for 1213 DNA samples, 1923 chromosomes, 1420 female chromosomes (n = 710) and 503 male chromosomes (n = 503). The GSA-24, v1.0, array is a multiethnic, genome-wide probe platform that includes clinically relevant variants validated by ClinVar, NHGRI, PharmGKB, and ExAC databases. On average, 3–5% of samples studied here have been previously investigated using different approaches, including the DMET array (Affymetrix), TaqMan genotyping, and targeted NGS; we collected these previous results from our archives and explored the concordance with GSA genotyping observing a consistent genotype agreement of >98.5%. We performed genomic descriptive statistics and model regressions using R (R Core Team, 2015), Admixture (Alexander and Lange, 2011), and PLINK (Purcell et al., 2007). We performed ancestry analyses considering individual admixture proportions as covariates during the regression analyses to account for population stratification (Hellwege et al., 2017).
Information on hypertension status (yes/no) was available for 1129 samples (512 hypertensive), for which we developed a logistic regression model adjusting for age, sex, body mass index (BMI), total cholesterol (TC), high-density lipoprotein (HDL), glucose, and the native ancestral component. ACE2 is located on the X chromosome, hence sex, in addition to altered blood lipids, can be a confounding factor. Therefore, we also performed genotype–phenotype associations, stratifying by sex and excluding all patients with dyslipidemia defined as low-density lipoprotein (LDL) >190 mmg/dL, HDL <40 mg/dL, triglyceride (TG) >130 mg/dL, and TC >240 mg/dL (Liu et al., 2018). Associations were developed by generating linear regression models using PLINK, defining SBP/DBP as the dependent variable, and adjusting by clinical and genetic covariates relevant to each model (Supplementary Table 1). The output, including effect sizes and statistical significance, was computed using the least absolute shrinkage and selection operator (LASSO) approach (Tibshirani, 2011). Comparisons between normotensive and hypertensive individuals stratified by genotype were also assessed (Supplementary Table S2).
Clinical Characteristics of the Study Population
Statistically significant.
Statistical differences were determined using the Mann–Whitney U test, all others were determined using the chi-square test, according to the frequency distribution of data.
Counts refer to the information available per clinic.
BMI, body mass index; DBP, diastolic blood pressure; IQR, interquartile range; SBP, systolic blood pressure.
Allele frequency differences may reflect different geographic ancestries, which (if not considered) may cause spurious genotype–phenotype associations. Therefore, admixture proportions were used as covariates in the logistic regression model to account for population stratification (Supplementary Table 1).
We were able to retrieve information on actual SBP and DBP for 636, or 321, samples after excluding those with altered blood lipid levels (dyslipidemia). SBP/DBP measurements were taken in duplicate by a certified nurse before and after a medical appointment, values were averaged and utilized in regression analyses. A list of nonredundant covariates was defined using an elastic net approach that relies on LASSO to identify covariates relevant to each model (Tibshirani, 2011; Sanchez-Ibarra et al., 2018). We developed linear regression models for SBP, adjusting for the heart rate (HR), waist circumference (WST), age, glucose levels, and LDL-cholesterol. DBP modeling included TG, HR, and WST as covariates; each model was stratified by sex with inclusion or exclusion of samples with altered blood lipids.
Results
Clinical characteristics of the study population are presented in Table 1, males and females were comparable for age and BMI, but showed significant differences for glucose levels, HR, SBP, and DBP. Admixture analyses showed a wide heterogeneity of the population. On average, we observed 47% of native ancestry (range: 0.0001–0.99997) and 31% of Caucasian ancestry (range: 0.0001–0.96092), in addition to 12% and 10% of Asian and African ancestral components (Supplementary Fig. S1).
Allele frequency of ACE2 variants
ACE2 genotypes were available for 1231 samples, allele and genotype frequencies were reported for females as diploid and for males as haploid. For variants, rs2048683 and rs4240157, we observed a 20% higher frequency of the minor allele (MAF) in males versus females, and for the other two variants, frequencies were comparable between the sexes (Table 2). Next, we compared MAF values between Mexican Mestizos and major continental populations, including African (AFR), American (AMR), East Asian (EAS), European (EUR), and South Asian (SAS) populations. The ACE2 rs4240157 variant showed the lowest MAF in Mestizos (17.7%) compared to all populations except EAS, for which we observed a threefold and twofold higher allele frequency versus Africans and Europeans. Similarly, the MAF of variant rs2048683 was lowest in Mestizos (15.6%) when comparing to all populations except EAS. For variants, rs2285666 and rs2106809, we observed an opposite trend, the MAF was highest for EAS, followed by SAS. Mestizos showed a significantly higher MAF when compared with AFR and EURs, but lower when compared with EAS and SAS. The most frequent variants in Mestizos were rs2285666 (39.5%) and rs2106809 (39.8%) and values were comparable with people of the Americas (AMRs) from the 1000 G database. In summary, for MAF observations, one could group variants rs2285666 and rs2106809, showing a similar MAF compared to AMR and 60% higher MAF compared to EUR. In contrast, variants rs2048683 and rs4240157 showed a twofold and 60% lower MAF in Mexican Mestizos compared with EUR and AMR (Table 2). Allele frequencies did not differ between normotensive and hypertensive individuals (Supplementary Table S2).
Allele and Genotype Frequencies of ACE2 Variants
ACE2 variants: rs4240157 G>A, rs2048683 A>C, rs2285666 A>G, and rs2106809 G>A. AFR, African; AMR, Native American; EAS, East Asian; EUR, European; SAS, South Asian; Mex. Mestizo, Mexican Mestizo from this study. A1: allele 1 as reported by the NCBI and depicted here, A2: allele 2 as reported by the NCBI and depicted here.
ACE2, angiotensin-converting enzyme 2.
Genotype–phenotype associations
Information as yes/no on hypertension status was available for 1129 samples. Exploratory logistic regression analyses identified no association between the four ACE2 SNVs and hypertension status after adjusting by age, sex, BMI, TC, HDL, glucose, and the native ancestral component (p > 0.221). Therefore, we sought to investigate associations between ACE2 variants and continuous variables, SBP and DPB, stratifying by sex and lipid levels.
For 636 samples, we retrieved actual SBP and DBP values and 321 of these presented normal blood lipids, that is, they did not show dyslipidemia (n = 321, males = 138 and females = 183; lipid profile: LDL <190 mmg/dL, HDL >40 mg/dL, TG <130 mg/dL, and TC <240 mg/dL). First, we performed regression analyses for SBP and DBP as continuous variables considering patients with a healthy lipid profile. ACE2 variants, rs4240157 and rs2048683, were significantly associated with SBP (p-value = 0.012 and p-value = 0.018) and DBP (p < 0.001, Table 3 and Supplementary Fig. S2). Interestingly, after stratifying by sex, rs2048683 was associated with DBP in females (p < 0.028) and rs4240157 with SBP in males (p < 0.021, Table 4). In both cases, the alternative allele was related to higher levels of SBP or DBP similar to the association observed when considering males and females together (p < 0.012, Table 3). In summary, two of four ACE2 SNVs were associated with SBP and DBP, and when stratifying by sex, these two SNVs remained significant, with one in females and the other one in males. A trend toward statistical significance in males and females for DBP and SBP was still observed for rs4240157 and rs2048683, respectively (p < 0.072, Tables 3 and 4 and Supplementary Fig. S2).
ACE2 Variants Associated with Systolic and Diastolic Blood Pressure in All Individuals
Statistically significant.
LDL <190 mmg/dL, HDL >40 mg/dL, TG <130 mg/dL, and TC <240 mg/dL. Association considering individuals with dyslipidemia and with normal lipid profiles.
HDL, high-density lipoprotein; LDL, low-density lipoprotein; SE, standard error; TC, total cholesterol; TG, triglyceride.
ACE2 Variants Associated with Systolic Blood Pressure and Diastolic Blood Pressure in Males Versus Females
Statistically significant.
Association considering individuals with a healthy lipid profile.
Association considering all individuals regardless of their lipid profile. Associations in individuals with a healthy lipid profile.
To corroborate if these associations were independent of altered lipid levels, we included all samples with available SBP/DBP data in the regression models (n = 636), including those with dyslipidemia. ACE2 variant rs4240157 remained statistically significant for DBP (p = 0.035) and a trend toward statistical significance for SBP was also noted (p = 0.054, Table 4). After gender stratification, this polymorphism remained significantly associated with DBP and SBP in males only (p < 0.050, Table 4).
Discussion
Dissection of population variation of ACE2 is still incomplete for several admixed populations and its association with blood pressure has not been fully assessed. Genotype data from GSA-24, v1.0, provided information on four ACE2 intron variants we report their allele and genotype frequencies in males and females as this gene is located on chromosome X. These four variants have not been classified by ClinVar as medically relevant. Nevertheless, research between 2018 and 2020 has reported clear associations between these variants and diabetes, lipids, and cardiovascular disease in Asian populations (Liu et al., 2018; Pan et al., 2018). Unfortunately, ACE2 nonsynonymous genetic variants that may offer information for SARS-CoV-2 infection were not available from this array. We explored phenotype–genotype associations for hypertension, SBP, and DBP. The few differences in allele frequencies observed were mostly for those variants associated with blood pressure, while rs2048683 and rs4240157 also showed MAF differences between males and females and when compared with people of the Americas (AMRs). These differences may be explained by population stratification for admixed individuals with several ancestral components (Hellwege et al., 2017), which have been consistently reported for other relevant gene variants in Mexican Mestizos (Bonifaz-Pena et al., 2014; Romero-Hidalgo et al., 2017).
Exploratory binary associations between ACE2 variants and hypertension status showed no statistical significance for any of the four variants analyzed. The effect of ACE2 on blood pressure has been reported and justified by assuming that ACE2 degrades the vasoconstrictor, angiotensin II, and thus counteracts its effects. In addition, it has been assumed that genetic variation that decreases or impairs ACE2 activity may contribute to cardiovascular disease, but the molecular mechanism behind these observations is far from understood. A direct lack of association between ACE2 genetic makeup and hypertension was earlier supported by Crackower et al. as the ablation of ACE2 did not alter blood pressure homeostasis in experimental models (Boehm and Nabel, 2002; Crackower et al., 2002); instead, ACE2 was shown to influence heart development and function. Studies from different regions in China have reported contradictory results for ACE2 polymorphisms and their relationship with essential hypertension and gender differences, pointing also to population stratification, sex, and ethnic differences (Fan et al., 2007; Li et al., 2015; Luo et al., 2019). More recently, ACE2 variants have been associated with cardiac features, DBP, and SBP, showing sex dimorphism (Unger et al., 2015; Zhang et al., 2018), but these studies are pending validation in admixed populations, and sex differences ought to be confirmed with consideration of potential confounding variables such as altered lipid levels.
In this study, we identified ACE2 rs4240157 and rs2038683 associated with SBP and DBP in a sex-dependent manner. These variants are in partial linkage disequilibrium (R 2 = 0.76, p < 0.0001) and their functional impact is not yet clear for cardiovascular disease, heart development, or SARS-CoV-2 infection.
ACE2 rs4240157 was consistently associated with SBP and DBP, but only with SBP in males. Similarly, Patel et al. (2012) investigated the impact of several ACE2 SNVs, including the one investigated here, rs4240157, on hypertension, increased ventricular mass, and cardiac function and found a genetic association more prevalent in men (Patel et al., 2012). We did not have specific cardiac function data to further support this genetic–blood pressure association, but this ACE2 intronic variant located between exons 4 and 5, together with rs5879922, rs4646156, and rs233575, forms a haplotype that has been recently associated with left ventricular mass, septal wall thickness, hypertrophy, cardiovascular disease in diabetic patients (Liu et al., 2018), and increased risk of essential hypertension and dyslipidemia in Chinese males (Pan et al., 2018). When our modeling included samples with dyslipidemia, ACE2 rs4240157 remained significantly associated with DBP, but again, only with SBP in males after gender stratification. Our observations on rs4240157 confirmed previous reports on the association of this variant with SBP and DBP predominantly in males. We identified that this association, ACE2 rs4240157-SBP, in males could be not only independent of altered lipid levels but also apparent for DBP when considering males and females together (Table 3 and Supplementary Fig. S2).
ACE2 variant rs2048683 lies between exons 14 and 15 and there are fewer than five publications on this variant. It has been recently associated with cardiovascular risk and hypertension-related organ damage in the Chinese population (Luo et al., 2019). This variant is in partial LD with rs2106809, which increases the risk for hypertension and reduces the antihypertensive response to ACE inhibitors (Fan et al., 2007). In this study, ACE2 rs2048683 was associated with SBP and DBP, but after stratification by sex, it remained significant for DBP in females only. Interestingly, this association did not hold when adding samples with an altered lipid profile to the regression model; that is, in individuals with a healthy lipid profile, the association was significant, but not for women showing nonhealthy blood lipids. The latter observation points again to sex differences putatively mediated by lipid levels. For example, female mice have shown that despite obesity, they are protected from hypertension due to high circulating Ace2 levels influenced by estrogen and diet (Shoemaker et al., 2019). The relationship between ACE2 and dyslipidemia has also been shown by Pan et al. (2018) with heterogeneity among genetic variants. It is possible that rs2048683 or other ACE2 polymorphisms in women are less relevant than their lipid profile or that women may have higher ACE2 levels despite genetic mutations driving the antiobesity effects of ACE2 by generation of Ang1–7 and browning of white adipose tissue (Kawabe et al., 2019).
In this regard, we also noted that the statistical impact of BMI and age in our models was log3-fold more significant in females compared with males. Similarly, a positive correlation was observed for age and SBP in females, but not in males (R 2 = 0.354, p < 0.001), suggesting that age and lipid composition in females are key determinants of cardiovascular health, supporting the notion that lipid profiles correlate with female healthy aging and cardiovascular protection, features not necessarily present in men (Gonzalez-Covarrubias et al., 2013). Our analyses did not include cardiac function data, but these genetic associations with SBP or DBP hint toward a potential role of specific heart features, which ought to be investigated.
In summary, we report that ACE2 rs4240157 and rs2038683 polymorphisms are associated with SBP in males and DBP in females, respectively, rs4240157 being independent of the presence of altered lipid levels. Our observations confirm sex differences in ACE2 genetic associations with SBP and DBP potentially related to the antiobesity and antiatherogenic effects of ACE2 (Thomas et al., 2010).
We did not identify statistical associations between SBP/DBP and variants rs2285666 and rs21088609. The former is located four bases before the codon CCG on exon 16, which can bear a common mutation to codon, TTC (glutamic acid). This variant has been associated with increased cardiovascular risk in patients with diabetes (Liu et al., 2018) and heart physiology in Caucasian hemizygous men (Lieb et al., 2006), but its association with arterial hypertension has been controversial (van der Merwe et al., 2008). Variant rs21066809 is listed by the Pharmacogenetics Knowledge Base (PGKB) as a risk variant for the efficacy of the ACE inhibitor, captopril (level 3) (Fan et al., 2007). Sexual dimorphism has been consistently reported for this polymorphism alone or in combination with ACE rs1799752, a 50-base in/del also listed by the PGKB as a level 2 mutation related to captopril efficacy. Together, both SNVs may increase hypertension risk by 1.6-fold and DBP in females by 2.3-fold (Fan et al., 2007). It is possible that other confounders such as treatment compliance or comorbidities, not officially registered in this study, are masking the effects of these genetic variants in our population.
Hypertension is a major burden to life expectancy, over nine million people die per year in the world because of hypertension and its complications (Oparil et al., 2018). Risk and treatment success for hypertension are multifactorial and current endeavors have characterized up to 5% of the 30–50% heritability of this trait. Future studies should prospectively investigate functional assays and full sequencing of ACE2 and candidate genes to aid in identifying the clinical impact of common and rare variations on hypertension and ACE2-related diseases. This study may be of relevance to address, in part, the epidemiology, population differences, drug targets, and prognosis of cardiovascular disease where ACE2 is involved, including the current COVID-19 pandemic. It is possible that together with clinical factors there is a genetic predisposition underlying COVID-19 incidence and severity. Research by the COVID-19 Host Genetics Initiative is already tackling the investigation into human variation and its relationship with COVID-19.
Footnotes
Acknowledgments
The clinical support from the COSFAMM team and the technical assistance of Cintia Guzman are gratefully acknowledged.
Disclosure Statement
No competing financial interests exist.
Funding Information
The present study was supported by funding from Conacyt-FOSSIS, grant No. 272795, to Vanessa Gonzalez-Covarrubias. Conacyt and UNAM granted a stipend for graduate students, Elba Rodriguez-Padilla and Tomas Texis-Valencia.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.