
Abstract
Pigs have been extensively used as the research models for human disease pathogenesis and gene therapy. They are also the optimal source of cells, tissues, and organs for xenotransplantation due to anatomical and physiological similarities to humans. Several breakthroughs in gene-editing technologies, including the advent of clustered regularly interspaced short palindromic repeat (CRISPR)/CRISPR-associated 9 (Cas9), have greatly improved the efficiency of genetic manipulation and significantly broadened the application of gene-edited large animal models. In this review, we have not only outlined the important applications of the CRISPR/Cas9 system in pigs as a means to study human diseases but also discussed the potential challenges of the use of CRISPR/Cas9 in large animals.
Introduction
Rodents remain the primary animal model used to study human diseases; however, rodents have a short life span, are small in size, and are not closely related to humans, often limiting the value of their use in in vivo experiments. As pigs are much more similar to humans with regard to their anatomical, physiological, pathological, metabolic, and disease characteristics, these animals are much more suited to the study of the pathogenesis of various diseases and the exploration of gene therapies. Furthermore, they are considered ideal organ donors for xenotransplantation.
The advent of several novel gene-editing technologies has accelerated the application of large animal models. However, previous gene-editing technologies, such as those involving the use of zinc-finger nucleases (ZFNs) and transcription activator-like effector nucleases (TALENs), are associated with a few disadvantages, including complicated operation steps, long carrier construction time, low target efficiency, and high costs, which limit their widespread application in the large animal models (Hickey et al., 2011; Bao et al., 2014).
The discovery of the clustered regularly interspaced short palindromic repeat (CRISPR)/CRISPR-associated 9 (CRISPR/Cas9) technology has generated a great deal of excitement within the gene-editing field as it improves the efficacy and simplifies the operational process of gene editing, thus, this technology has been widely used in the field of gene editing for the establishment of animal models (Wan et al., 2015). In this review, we discuss the recent achievements in CRISPR/Cas9-mediated gene editing in pigs, as well as the challenges of the use of CRISPR/Cas9. Given our focus on porcine genome editing by CRISPR/Cas9 in this review, the sophisticated application of TALEN and ZFN-mediated genome editing in pigs, is not discussed here. A summary of CRISPR/Cas9-derived gene-edited pigs for medical research is presented in Tables 1 and 2.
Overview of Gene Edited Pigs Generated by Clustered Regularly Interspaced Short Palindromic Repeat/Cas9 for Medical Research via Somatic Cell Nuclear Transfer
This table provides an overview of used vectors, doses, method for vector transfection, mutation efficiency, and research purpose for the production of gene-editing pigs by SCNT.
A, PiggyBac-Cas9/2gRNAs(pol) plasmids; B, transgene expression plasmids; C, Cas9 expression plasmids; D, β4GALNT2 sgRNA expression plasmids; E, CMAH gRNA expression plasmids; F, GGTA gRNA expression plasmids; G, PiggyBac plasmids; GHR, growth hormone receptor; hIAPP, human islet amyloid polypeptide; INS, insulin; IRX3, iroquois homeobox 3; NGN3, Neurogenin 3; SCNT, somatic cell nuclear transfer; sgRNA, single-guide RNA.
Overview of Gene-Edited Pigs Generated by Clustered Regularly Interspaced Short Palindromic Repeat/Cas9 for Medical Research via Embryonic Microinjection
This table provides an overview of used vectors, doses, mutation efficiency, and research purpose for the production of gene-edited pigs by embryonic microinjection.
ALB, albumin; IL2RG, Interleukin 2 receptor gamma; MITF, microphthalmia-associated transcription factor; RAG2, recombinant activating gene 2; TYR, tyrosinase.
The Mechanism of the CRISPR/Cas System
The CRISPR/Cas system is essentially a unique set of RNA-guided endonucleases found in ∼40% of bacteria and 90% of archaea that act as an acquired defense system allowing them to resist invasion by foreign genetic elements (Garneau et al., 2010). The CRISPR/Cas system functions in three phases: adaptation, expression, and interference.
In the adaptation step, short pieces of foreign DNA are captured and integrated as “spacer” elements into the CRISPR loci. Then, these integrated CRISPR sequences are transcribed with the help of RNA polymerase and spliced into mature CRISPR RNA (crRNA), which forms a complex with trans-acting crRNA creating the single-guide RNA (sgRNA), which in turn, acts as the guide sequence for directed Cas activity (Jinek et al., 2012; Cong et al., 2013).
In the interference phase, the protospacer adjacent motif in the sequence of the exogenous nucleic acids allows the targeted recognition of the sgRNA-guided Cas (such as Cas9, Cas12, and Cas13) and mediates their interaction, initiating the cleavage of DNA cleavage and creating a double-strand break (DSB) (Chen et al., 2018; Zhao et al., 2018). This process can be achieved exogenously through recombinant proteins and synthetic guides in living cells.
In cells, the DSB will give rise to DNA repair processes, which often result in random indels through nonhomologous end joining (NHEJ); the indels mostly cause frameshift mutations. If a DSB is repaired by the homology-directed repair (HDR) pathway, the donor template DNA with a homologous sequence corresponding to the target region can introduce the desired fragment into the target site (Fig. 1). CRISPR/Cas systems can be classified into three major types (I, II, and III). The most commonly used system is the type II CRISPR-Cas9 system from Streptococcus pyogenes (Cong et al., 2013). The high efficiency and simplicity of this system has facilitated its widespread adoption in genome-targeting experiments.

Schematic of the CRISPR/Cas9 system. The sgRNA-guided Cas9 protein recognizes and cuts target genes. The Cas9-induced DSBs are then repaired by DNA repair processes, which often result random indels through NHEJ, and the indels mostly cause frameshift mutations. If DSB is repaired by HDR, the donor template DNA can result in a specific alteration. CRISPR, clustered regularly interspaced short palindromic repeat; DSB, double-strand break; HDR, homology-directed repair; NHEJ, nonhomologous end joining; PFFs: porcine fetal fibroblasts; sgRNA, single-guide RNA.
The Methods of Generating Gene-Edited Pigs
The previous strategies used to modify the porcine genome mainly include pronuclear microinjection (Golovan et al., 2001; Saeki et al., 2004), retrovirus-mediated transgenesis (Cabot et al., 2001), sperm-mediated gene transfer (SMGT) (Lavitrano et al., 2003; Webster et al., 2005), and transposon-mediated transgenesis (Wu et al., 2013). However, some limitations have hampered their wide application. Pronuclear microinjection, virus-mediated transgenesis, and transposon-mediated gene transfer can only introduce random transgenes and these approaches lack control of the copy number of the integrated gene. For the virus-mediated gene transfer, there are potential carcinogenicity and safety concerns. Although the method of SMGT is simple and low-cost, it is difficult for foreign DNA to penetrate sperm, and it followed with low integration efficiency and poor repeatability (Wu et al., 2008). In 1996, the success of somatic cloning of sheep promoted the rapid development of transgenic research in large animals (Campbell et al., 1996). Soon after, the gene-edited pigs were successfully generated by somatic cell nuclear transfer (SCNT) combining with the homologous recombination (HR) (Polejaeva et al., 2000; Lai et al., 2002). However, the HR has an extremely low efficiency. With the development of ZFNs, TALENs, or CEISPR/Cas9, a high rate of DSB can be induced and that can result in a high rate of specific mutation either in somatic cells or in embryos.
Currently, editing the somatic cells with CRISPR/Cas9 followed by SCNT or direct injection of the CRISPR/Cas9 system into embryos at the single-cell stage is the most common method to obtain genetically modified animals. The advantages of SCNT are that the researchers can precisely screen modified cells with homozygous or heterozygous mutations in vitro, and then the cells can be used in cloning to produce a series of known and characterized offspring (Yum et al., 2018). However, reconstructed embryos obtained by SCNT technology are usually associated with some problems, including low efficiency, early fetal mortality, and developmental defects; this reflects the disadvantages of using SCNT (Betthauser et al., 2000; Onishi et al., 2000; Zhang and Li, 2018).
Another way to generate gene-edited animals is via the direct injection of the CRISPR/Cas9 system into embryos at the single-cell stage. In recent years, several studies have shown that a sgRNA and Cas9 mRNA complexes, or sgRNA/Cas9 protein complexes (RNP), can be injected into embryos to efficiently generate gene-edited animals (Hai et al., 2014; Chen et al., 2019; Camargo et al., 2020). However, the chimeras may occur, and it is difficult to obtain sufficient numbers of single-cell stage embryos for large animals (Lei et al., 2016; Hinrichs et al., 2018). The two major routes for generating gene-edited pigs with CRISPR/Cas9 are shown in Figure 2.
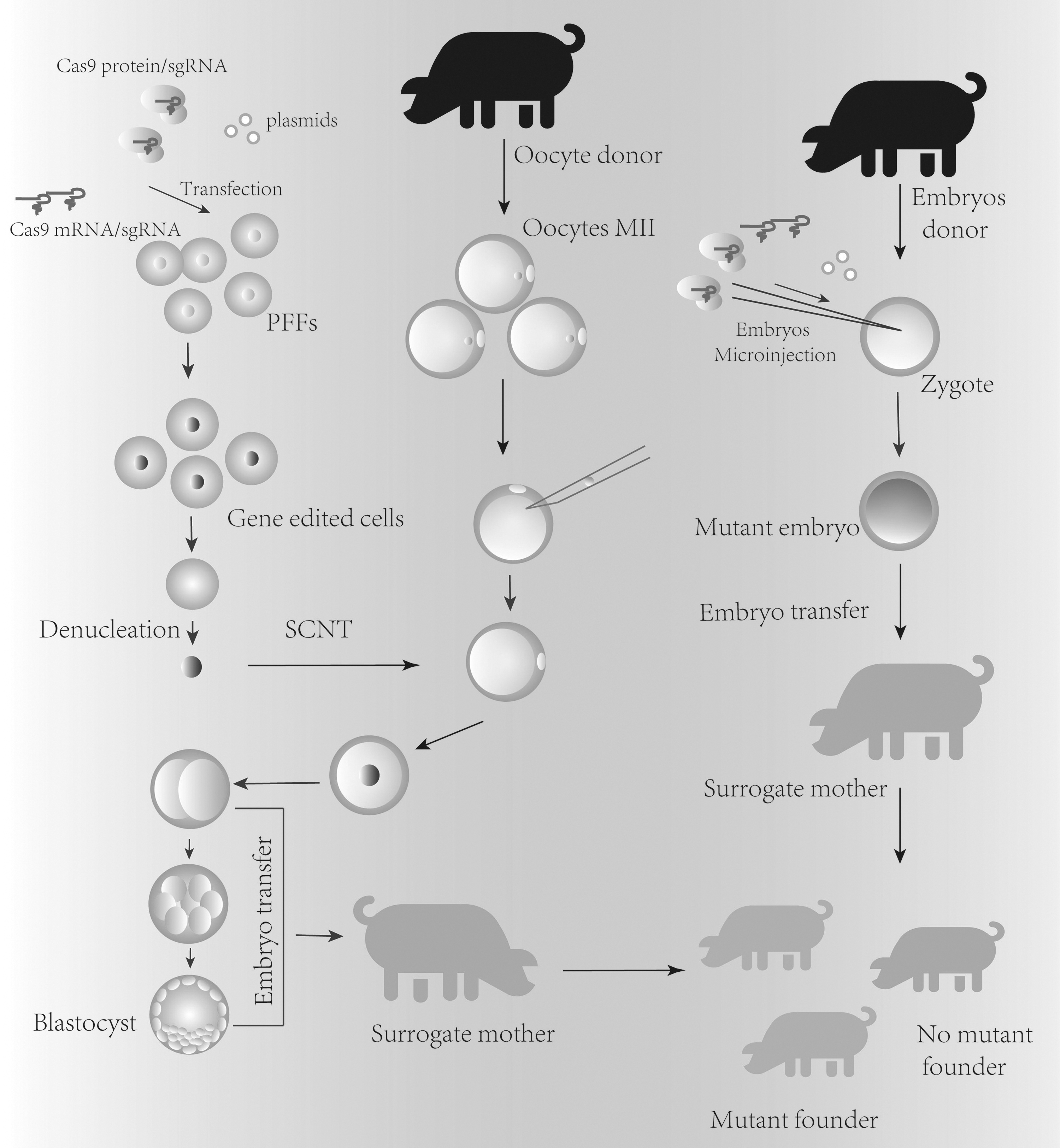
Schematic of the approach to generate gene-edited pigs via CRISPR/Cas9. For the SCNT, first, oocytes in MII phase were harvested, then the gene-edited somatic cells as the nuclear donors are injected into the mature oocytes. The edited embryos are then cultured and implanted in a surrogate mother. For the embryo microinjection, first, the one-cell stage fertilized eggs were injected CRISPR components (Cas9 and sgRNA RNPs, or Cas9 mRNA and sgRNA molecules or plasmids). Then, the edited embryos are cultured and implanted in a surrogate mother. SCNT, somatic cell nuclear transfer.
The Gene-Edited Pig Models for Medical Research
Gene-edited pigs for xenotransplantation
Pigs are the preferred source for potential organ xenotransplants designed to circumvent the shortage of human organ donors. However, immune rejection and the potential risk of porcine endogenous retroviruses (PERVs) are two major hurdles that restrict the development of xenotransplantation protocols.
In 2015, Yang et al. inactivated 62 copies of the PERV genes in porcine PK15 across the genome using CRISPR/Cas9, reducing the risk of human infection with PERVs by more than 1000-fold (Yang et al., 2015). Soon afterward, this team was able to produce PERV-inactivated porcine fetal fibroblasts using CRISPR/Cas9, which they combined with SCNT technology to produce PERV-inactivated pigs (Yang et al., 2015). This study makes a significant contribution toward preventing interspecies transmission of PERVs and addressing the biosafety concerns associated with the clinical application of xenotransplantation.
The hyperacute rejection (HAR) can result in rapid destruction of the graft, usually within minutes to hour (Platt et al., 1991). HAR is mediated by the antibodies that naturally existed in the human. When they bind the xenoantigenic epitopes on porcine endothelial cells that could trigger activation of complement proteins, and further lysis of endothelial cells, it results in the graft failure (Gollackner et al., 2004).
The galactose-alpha-1,3-galactose (αGal, produced by alpha-1,3 galactosyltransferase GGTA1), N-glycolylneuraminic acid (Neu5Gc, produced by cytidine monophosphate-N-acetylneuraminic acid hydroxylase, CMAH), and SDa epitopes (produced by beta-1,4-N-acetyl-galactosaminyltransferase 2, β4GALNT2) are the major xenoantigens which exist in pigs, but not in humans (Byrne et al., 2018). They precipitate both hyperacute and acute rejection in pig-to-human xenotransplantations (Hein et al., 2020).
In 2015, Estrada et al. generated a GGTA1/CMAH/β4GalNT2 knockout pig using CRISPR/Cas9. These pigs were shown to exhibit reduced human IgM and IgG antibody production, suggesting that this model could reduce the antigenicity of pigs for xenotransplantation (Estrada et al., 2015). In 2016, Petersen et al. reported that cytoplasmic microinjection of a CRISPR/Cas9 plasmid produced GGTA1 homozygous mutations in pigs (with a biallelic mutation rate of up to 50%) (Petersen et al., 2016). In 2017, Gao et al. obtained 11 biallelic GGTA1/CMAH mutant Wuzhishan pigs using manual cloning; however, this method was complex and resulted in low live birth rates (2.5%) (Gao et al., 2017).
In 2018, another group produced a GGTA1/CMAH/β4GalNT2 triple gene-knockout (TKO) pig; they proved that the tissues from TKO pigs showed a further reduction in the ability to bind to human IgG/IgM binding in the porcine pericardium and that these pigs could be used to produce heart valve replacements for clinical use (Zhang et al., 2018a). In 2019, Tu et al. used embryo electroporation to produce four biallelic CMAH KO pigs (Tu et al., 2019).
Major histocompatibility complex (MHC) molecules are the main antigens that cause T cell-mediated rejection in xenotransplantation (Sullivan et al., 1997). To alleviate the cellular immune response, in 2019, Sake et al. used CRISPR/Cas9 to successfully deplete the porcine beta-2-microglobulin (β2M) gene, which abrogate SLA class I expression (Sake et al., 2019).
In 2020, to eliminate HAR and T cell immunity simultaneously, researchers generated α-Gal, SLA-I, and SLA-II triple-deficient pigs through depleted GGTA1, β2M, and MHC class II transactivator (CIITA) genes using CRISPR/Cas9 and SCNT. The skin grafts of triple KO pigs significantly improved survival in immunocompetent C57BL/6 mice (Fu et al., 2020).
In 2020, Yue et al. generated GGTA1, CMAH, β4GALNT2 knockout pigs that also overexpressed nine human transgenes (hCD46, hCD55, hCD59, hB2M, HLA-E, hCD47, hTHBD, hTFPI, and hCD39) in an effort to enhance the immunocompatibility and coagulation compatibility between pig and human tissues. Based on this, they inactivated all the PERVs from these knockout and transgenic pigs, improving both the safety and efficacy of any potential xenotransplants (Yue et al., 2020).
Given that one of the goal of producing gene-edited pigs is that use them as human organ donors, since GGTA1 KO (GTKO) pigs were generated, multiple gene-edited pigs based on the GTKO are already being used to prolong the xenograft survival time in nonhuman primates(NHPs).
In 2005, the hearts from GTKO pigs were transplanted into immune-suppressed baboons, which extended the survival of pig hearts in baboons to as long as 179 days (Kuwaki et al., 2005). In 2012, the GTKO/hCD46 transgenic pig hearts were transplanted into baboons and the longest survival for heterotopic cardiac xenografts in baboons was extended to 236 days (Mohiuddin et al., 2012). Subsequently, this group transplanted GTKO/hCD46/hTBM transgenic pig hearts to baboon models administered CD40 antibody-based immunomodulatory regimen, and the longest survival time was extended to 945 days (Mohiuddin et al., 2016).
However, these models demonstrated long-term acceptance of xenografts but did not predict their life-supporting function. In 2018, the xenografted hearts from GTKO/hCD46/hTBM transgenic pig provided a consistent life-supporting of baboons to 195 days (Längin et al., 2018). In their study, two steps were a key to success. First, nonischemic porcine heart preservation was performed. Second, they prevented the detrimental xenograft overgrowth by a drug called temsirolimus. In 2018, Adams et al. transplanted the kidneys from the GGTA1/B4GALNT2 double KO pigs into rhesus monkeys, which were immunosuppressed with T cell depletion, anti-CD154, mycophenolic acid, and steroids. The recipients achieved life-supporting survival to 435 days (Adams et al., 2018).
In 2019, this group reported that depleting the CD4+ T cell of rhesus macaque before pig renal xenografts transplantation could keep a consistent survival over 1 year (Kim et al., 2019). The studies with pig-to-NHP xenotransplantation suggesting that selecting the optimal gene edited pig as the organ source and immunosuppressive regimen strategy in pig-to-NHP models are crucial to further clinical application.
In addition to being used as organ donors for xenotransplantation, the immunodeficient pigs are expected to be used in cell xenografts and as bioreactors for the production of human cells for transplantation.
In 2015, to generate B cell-deficient pigs, Chen et al. used CRISPR/cas9 system to target IgM heavy-chain gene. Three biallelic mutation piglets were produced, and the piglets showed no antibody-producing B cells, the pigs could be further engineered for large-scale production of human antibody (Chen et al., 2015).
Interleukin 2 receptor gamma (IL2RG) gene mutations can cause severe X-linked immunodeficiency disease. In 2016, Kang et al. obtained three IL2RG pigs using microinjection and SCNT. These pigs were characterized by their lack of a thoracic gland, which significantly reduced B cell, T cell, and natural killer cell production. This model can be used to further research on heterogeneous cell regeneration (Kang et al., 2016).
In 2016, Lei et al. generated recombinant activating gene 2 (RAG2)/IL2RG double-knockout pigs using CRISPR/Cas9 injected into embryos (Lei et al., 2016). In 2019, Chen et al., also injected Cas9 mRNA and multiple sgRNAs into cells to obtain tyrosinase (TYR), TYR/IL2RG, and TYR/recombinant activating gene 1 (RAG1) knockouts, recapitulating the typical phenotypes associated with albinism and immunodeficiency (Chen et al., 2019).
In summary, these studies have laid the foundation for the efficient production of heterogeneous organ donor pigs with low immune rejection and may facilitate studies of heterogeneous cell regeneration in immunodeficient gene-edited pigs.
Humanized gene-edited pigs
The use of technologies such as blastocyst complementation to create human organs derived from gene-edited chimeric pigs would provide an alternative approach to overcome the immunologic barriers to xenotransplantation.
In 2017, Wu et al. injected 3–10 human pluripotent stem cells into the wild-type porcine blastocysts, and the chimeric embryos were collected at embryonic day 28. They found differentiated human derivative cells but show limited chimeric contribution to postimplantation pig embryos (Wu et al., 2017a). Then, the group used CRISPR/Cas9 technology to knockout PDX1 to inhibit the development of pancreas in pigs, creating an appropriate platform for the further development of human organs combining interspecies blastocyst complementation in pigs (Wu et al., 2017b).
In 2020, Satyabrata et al. generated ETV2 deficient porcine embryos by CRISPR/Cas9. The ETV2 deficient could lead to embryonic lethal, as the embryos lacked hematoendothelial lineages. Then they injected the human induced pluripotent stem cells (hiPSCs) or hiPSCs overexpressing the antiapoptotic factor BCL2 into ETV2-null blastocysts. The chimeric embryos were transferred to synchronized gilts and analyzed between embryonic day 17 and embryonic day 18. The result shows that all endothelial cells of the embryos were human origin. This is a big step toward generating exogenous organs (Das et al., 2020).
To solve the shortage of human serum albumin, which is a common human blood product, the human albumin (ALB) cDNA construct was inserted into the pig Alb locus using CRISPR/Cas9 combined with embryo microinjection. The researchers used KI pigs to produce recombinant human serum albumin in their serum (Peng et al., 2015).
Pig hepatocytes are widely used in bioartificial liver (BAL) clinical trials. In an attempt to generate pigs expressing humanized liver proteins, Li et al. used CRISPR/Cas9 and SCNT to successfully establish KI pigs expressing human blood coagulation factor VII (F7) and ALB (Li et al., 2019). The pigs could be used as an alternative cell source for BALs, which would provide a promising treatment for patients with acute liver failure.
Using gene-edited pigs to model neurodegenerative diseases
To construct a large animal model of Parkinson's disease, Zhou et al. produced Parkinson disease 2 (PARK2) and PTEN-induced putative kinase 1 (PINK1) homozygous knockout pigs via the CRISPR/Cas9-based editing of fibroblast cells and SCNT. These double-gene knockout pigs did not express the PINK1 protein (Zhou et al., 2015). Simultaneously, Wang et al. injected Cas9 mRNA and sgRNA complexes targeting the PARK2, Parkinson disease 7 (DJ-1), and PINK1 genes into porcine embryos and rapidly obtained three piglets with homozygous mutations (Wang et al., 2016). Zhu et al. also attempted to model PD by generating missense mutations in α-synuclein (SCNA) (Zhu et al., 2018). However, none of the piglets obtained Parkinson's symptoms, possibly because the animals were still relatively young.
Huntington's disease (HD) is a neurodegenerative disease characterized by the selective loss of medium-sized spiny neurons within the striatum. In 2018, Yan et al. established an HD pig model using CRISPR/Cas9 and SCNT (Yan et al., 2018). This model expressed the endogenous full-length mutant huntingtin protein (HTT) and the researchers were able to maintain this trait in both the first- and second-generation animals. The offspring of the HD pigs showed signs of movement, behavioral abnormalities, and premature death, which was consistent with their parental phenotype. Therefore, this HD pig model can be used to study the pathogenesis and treatment of this neurological disease in large mammals.
Using gene-edited pigs to model blood diseases
A defect in the von Willebrand factor (vWF) gene represents a common hereditary hemorrhagic disease. Hai et al. researchers produced 16 piglets with defects in this gene by the injection of Cas9 mRNA and CRISPR sgRNAs into pig embryos, producing a mutation rate of 68.8%. The clotting time of the biallelic mutants was significantly higher than that of the wild type (Hai et al., 2014). Hemophilia B is an X-linked recessive bleeding disorder caused by abnormalities in the coagulation factor IX gene. In 2020, Chen et al. generated porcine F9 (pF9) knockout pigs and human F9 (hF9) knock-in pigs using CRISPR/Cas9 and SCNT. This study not only produces a model for studying the pathogenesis of hemophilia B but also represents the first model available for the evaluation of the genetic correction of hemophilia (Chen et al., 2021).
Using gene-edited pigs to model human cancers
Runt-related transcription factor 3 (Runx3) gene is thought to be a novel tumor suppressor gene, and its deletion has been shown to promote the development of gastrointestinal tumors (Levanon and Groner et al., 2004; De Bruijn and Dzierzak et al., 2017). In 2016, Kang et al. used CRISPR/Cas9 and SCNT to produce five monoallelic Runx3-modified pigs, which can be used to study tumor occurrence and therapy (Kang et al., 2016).
TP 53 (which encodes p53) is one of the most frequently mutated genes in cancer. In 2018, TaniharaI et al. generated TP53-mutant pigs using CRISPR/Cas9 (Tanihara et al., 2018). These mutant pigs exhibited various tumor phenotypes from the age of 16 months.
In 2017, Wang et al. established a pig line with Cre-inducible Cas9 expression. Thus, via the intranasal delivery of the Cre recombinase and sgRNAs using a lentiviral vector that targeted several tumor suppressor genes, including TP53, PTEN, APC, BRCA1, and BRCA2 and the oncogene KRAS in Cas9-expressing pigs, they successfully generated a novel porcine lung cancer model (Wang et al., 2017).
Using gene-edited pigs to model genetic disorders
Microphthalmia-associated transcription factor (MITF) is a key regulator of melanocyte development and acts an important oncogene in melanoma. Mutations in the human MITF gene have been shown to result in both hypopigmentation and deafness. This means that MITF knockout models are a valuable resource in several human conditions. In 2015, researchers obtained two biallelic MITF knockout pigs using the embryonic microinjection. These models were successfully used to simulate human Werner syndrome (Wang et al., 2015).
Mutations in the growth hormone receptor (GHR) gene can cause Laron syndrome, a human autosomal disease. Yu et al. used a dual-sgRNA/Cas9 system and SCNT to generate biallelic GHR mutant pigs with low growth rates and smaller body size. They went on to generate an F2 generation of animals that retained these phenotypic traits, suggesting that that these GHR alleles could be passed to the next generation via germline transmission (Yu et al., 2018). In addition, Hinrichs et al. injected CRISPR/Cas9 into in vitro-fertilized porcine oocytes and obtained pigs with several monoallelic GHR mutations (Hinrichs et al., 2018). The GHR mutant pigs resemble the pathophysiology of Laron syndrome and can be used as a large animal model for mechanistic studies and treatment trials.
Hutchinson-Gilford progeria syndrome (HGPS) is an extremely rare genetic disorder affecting a very small proportion of the human population. Dorado et al. generated a porcine model of HGPS with heterozygous LMNA c.1824C>T site mutations using CRISPR/Cas9-mediated HR. HGPS minipigs exhibited severe growth retardation, lipodystrophy, and cardiovascular disease (Dorado et al., 2019). The HGPS pigs provide a preclinical model to test human-size interventional devices and optimize cardiovascular therapies before advancing to clinical trial.
Homeobox 13 (Hoxc13) is a pathogenic gene involved in hypotrichosis and nail dystrophy. To generate a hairless model for hair regeneration research, Han et al. generated Hoxc13 site mutations in pigs using CRISPR/Cas9-mediated HDR (Han et al., 2017).
Using gene-edited pigs to model metabolic disorders
Increased deposition of human islet amyloid polypeptide (hIAPP) is considered one of the most common causes of type 2 diabetes mellitus (T2DM). As there is no recognized ideal animal model that is fully consistent with the characteristics of human T2DM, in 2019, Zou et al. successfully generated IAPP gene-humanized pigs using CRISPR/Cas9 and SCNT. Subsequent evaluations demonstrated that 3-month-old hIAPP pigs presented with hyperglycemia, decreased glucose utilization, and increased insulin (INS) tolerance (Zou et al., 2019). Their study provided a new model for the pathogenesis and preventive measures of diabetes and its complications
In an effort to develop a large animal model of diabetes mellitus, Cho et al. generated INS gene-deficient pigs using CRISPR/Cas9 and SCNT; however, all three piglets died within 2 days (Cho et al., 2018).
Neurogenin 3 (NGN3) is a transcription factor involved in endocrine pancreatic development. In 2018, Sheets et al. used CRISPR/Cas9 to generate three live biallelic NGN3 knockout pigs who were not able to produce INS (Sheets et al., 2018). This research offers a model in facilitating generation of human islets (endocrine cells) within NGN3-deficient pigs.
The iroquois homeobox 3 (IRX3) gene is known to be relevant to human obesity. Zhu et al. successfully generated IRX3 biallelic cloned pigs with markedly poor viability by CRISPR/Cas9 combined with SCNT; all the piglets died shortly after birth (Zhu et al., 2020).
Using gene-edited pigs to model infectious diseases
Hepatitis E is an acute infectious disease caused by hepatitis E virus (HEV). In 2018, Yugo et al. generated an immunoglobulin heavy chain JH knockout gnotobiotic pig model for HEV infection via the CRISPR/Cas9 system. These HEV-infected JH knockout pigs had significantly enlarged livers. The availability of this novel model will facilitate studies of HEV pathogenicity (Yugo et al., 2018).
Using gene-edited pigs to model cardiovascular disease
Apolipoprotein E (ApoE) and low-density lipoprotein receptor (LDLR) gene mutations play a critical role in the development of atherosclerosis. In 2017, Huang et al. generated six biallelic knockout pigs, ApoE− /−//LDLR− /−, using CRISPR/Cas technologies. These animals were shown to have significantly elevated serum levels of low-density lipoprotein cholesterol (LDL-C), total cholesterol, and apolipoprotein B (Huang et al., 2017). In 2018, Burke et al. used the LDLR-deficient pigs to evaluate the capacity of bempedoic acid to decrease plasma cholesterol and LDL-C levels and attenuate atherosclerosis. This animal model could be used for the preclinical assessment of novel cardiac/cholesterol drugs (Burke et al., 2018).
Challenges Associated with of the Use of CRISPR/Cas9
CRISPR-Cas9 as a newly developed gene-editing tool can simultaneously target multiple genes with high efficiency, and it is easy to use with a low cost, which has enabled the efficient generation of genetically modified pigs, including knock-in and knockout variants. However, there are still some limitations of the gene-editing technology, including the packaging and delivery of CRISPR/Cas9, the low knock-in efficiency, and how to perform precise gene correction to induce effective CRISPR therapy.
The efficient and safe delivery technologies are crucial for the CRISPR/Cas9 systems to effectively enter the target cells and perform their function. Both viral and nonviral methods have been adopted to deliver CRISPR/Cas9 in vitro or in vivo.
Compared with viral vectors, nonviral delivery systems have the potential to be less toxic and immunogenic and there is no size limitation for transgenic DNA. There are three forms of CRISPR/Cas9 that are commonly used to be delivered into cells by nonviral vectors: (1) plasmid-based CRISPR/Cas9; (2) the mixture of Cas9 mRNA and the sgRNA; and (3) the mixture of Cas9 protein and sgRNA.
At present, several nonviral delivery strategies are widely used, such as microinjection (Hai et al., 2014), electroporation (Qin et al., 2015), nanoparticles (Zuris et al., 2015), and hydrodynamic injection (Liu et al., 1999). Each of these approaches has its own advantages and disadvantages.
The electroporation can be used for any cell type with a high transfection efficiency, however, it induces significant cell death (Frangoul et al., 2021).
The Cas9 protein and sgRNA compounds can be directly injected into the one-cell stage zygotes with a rapid action and low off-target effects; however, it requires a sophisticated micromanipulation and comes with low-throughput (Wang et al., 2015).
The nanoparticles are easy to prepare and some of them have entered clinical trials for RNAi therapy, but the delivery efficiency is low (Coelho et al., 2013; Fitzgerald et al., 2014).
Hydrodynamic injection are used to in vivo therapies, in which a high-volume of nucleic acid solution (8–10% body weight) are injected into the tail-vein of rodents, but it remains acute side effects that limited the use in humans (Dul et al., 2017).
Nonviral delivery of CRISPR/Cas9 is used flexibly for in vitro gene editing and generating animal models. However, the viral vectors are regarded as the higher efficacy tools to deliver CRISPR systems for in vivo therapeutic applications (Paulk et al., 2010; Koike-Yusa et al., 2014).
Lentivirus (LV), adenovirus (AdV), and adeno-associated virus (AAV) are the commonly used viral vectors for CRISPR delivery.
The LV-mediated CRISPR/Cas9 delivery systems have been used to generate cancer models on mice and pig (Platt et al., 2014; Wang et al., 2017). However, the LV can randomly integrate viral DNA into host cells and lead to long-term protein expression, which remains the security concerns in clinical application.
As the AdV are not integrated into the host cell genome, they are used for the transient expression CRISPR/Cas9 (Gao et al., 2019).
At present, AAV is the most widely used strategies of delivery CRISPR/Cas9 in vivo for gene therapy and preclinical studies (Rossidis et al., 2018; Villiger et al., 2018). However, only ∼4.5 kb of genome is allowed to be packaged into AAV. The barrier for AAV-mediated CRISPR-Cas9 delivery is the large size of Streptococcus pyogenes Cas9 (SpCas9) protein (∼4.2 kb). To overcome this, the smaller SaCas9 was used to AAV packaging.
The SaCas9-AAV system that has been used acutely infected the animals in vivo to achieve gene editing (Ran et al., 2015; Yin et al., 2017; György et al., 2019). However, another limitation for the AAV-mediated gene therapy was the immune response, as a large proportion of people and animals have preexisting antibodies against the AAV virus. In addition, the researchers indicate that there are preexisting humoral and cell-mediated adaptive immune responses to SaCas9 and SpCas9 both in human and animals (Charlesworth et al., 2019; Li et al., 2020a, b). So the finding to reduce the risks of security for the therapeutic applications should be further explored, such as in combination with immunosuppression therapies.
When the CRISPR system has been delivered efficiently to target cells, more efficient and accurate gene-editing tools are needed to achieve precise gene correction on large animals or human. Compared with CRISPR/Cas9, the cytosine base editor (CBE) or the adenine editor (ABE) can achieve accurate single base replacements (C-to-T, or A-to-G) without the introduction of double-stranded DNA breaks and donor templates (Gaudelli et al., 2017; Kim et al., 2017; Komor et al., 2017). Futhermore, in 2021, Kurt et al. created a new base editor by engineering two base editor architectures that can efficiently induce targeted C-to-G base transversions (Kurt et al., 2021). Recently, some teams have demonstrated the effectiveness of the base editor on pigs.
In 2019, Xie et al. used the BE3 base editor to efficiently induce C-to-T conversions at multiple sites, including Duchenne muscular dystrophy (DMD)/TYR/LMNA, or RAG1/RAG2/IL2RG, in porcine embryos and fibroblasts (Xie et al., 2019). The efficiency of C-to-T conversions in three genes simultaneously at the embryonic level was ∼40–50%, and the single and multiple gene mutant pigs were generated. In addition, they also disrupt the pol genes (porcine endogenous retrovirus) with multiple copies by introducing multiple premature stop codons in porcine embryos and fibroblasts. About 85% copies of pol genes were induced C-to-T conversions. The results of their study showed that introducing stop codons to destroy the genes with multiple copies could avoid DNA damage compared with DSB-dependent knockout methods.
In 2020, Yuan et al. obtained GGTA1, CMAH, and β4GalNT2 deficient pig by precisely converting the base from C-to-T (Yuan et al., 2020). Also, they reported that using the new CBE version (termed AncBE4max) in porcine blastocysts and fibroblasts could increase the efficiency of three genes base editing to 71.4%.
In 2021, Yao et al. used CRISPR/Cas9 with long donor DNA as the repair template to achieve precise correction on a pig model that carries the c.740T>C (L247S) mutation in MITF (Yao et al., 2021). Then they evaluated the efficiency and safety of CRISPR/Cas9 and base editors-mediated correction. Their study showed that the correction efficiency was greatly improved by BES in mutant porcine fibroblast cells and early embryos but higher off-targeting mutations were detected. Thus, the safety should be considered for their potential applications in gene therapy.
As we can see from the current studies, although the efficiency of gene knockout has been high, the efficiency of Knock-in still needs further improvement, including HDR-mediated gene correction (Wang et al., 2019; Zhao et al., 2020). The CRISPR/Cas9-mediated DSB was repaired by NHEJ or HDR. As the NHEJ is faster than HDR and the NHEJ suppresses the HDR process, some researchers use small molecules, such as KU70, KU80, ligase IV inhibitor SCR7, or inhibit the 53BP1 to restrict the NHEJ pathway, or treatment with the factors in favor of HDR such asRS-1 to enhance the HDR pathway (Chu et al., 2015; Maruyama et al., 2015; Canny et al., 2018; Zhang et al., 2018b; Jayavaradhan et al., 2019). In addition, NHEJ is active throughout the cell cycle, whereas HDR occurs largely during S/G2 phases, so regulate the cell cycle or control the delivery time of preassembled Cas9 protein and sgRNA complex to the target cells could increase the frequency of HDR (Lin et al., 2014).
Conclusion
In summary, the rapid development of CRISPR/Cas9 has revolutionized the production of gene-edited large animals and facilitated the biomedical applications in human diseases, xenotransplantation. Nevertheless, the in vivo delivery strategy, biological incompatibility, and cytotoxicity of delivery tools remain significant challenges in CRISPR-mediated gene therapy using large animal models. Moreover, the efficiency of gene correction, such as CRISPR/Cas9-based HDR or precise base transversions, should be facilitated by the increased understanding of DNA repair pathways and the development of novel base editors. These potential gene-editing tools for gene therapy still need long-term preclinical testing in animal models.
Footnotes
Authors' Contributions
M.G. and X.Z. wrote the article. J.B., H.B., and G.Y. designed and revised the article. All authors reviewed the final article.
Disclosure Statement
No competing financial interests exist.
Funding Information
The research program was supported by the National Key Clinical Project. The research leading to these results has also received funding from National Natural Scientific Foundations of China (81770618 and 82070640); Major Science and Technology Project of Sichuan Science and Technology Department (2019TFS0138); Technology Innovation Project of Chengdu New Industrial Technology Research Institute (2018-CY02-00046-GX).