
Abstract
Several studies have reported that miR-885-5p was dysregulated in a variety of cancers. However, there are few studies on the biological function of miR-885-5p in gastric cancer (GC). In this study, we investigated the biological function and underlying mechanism of miR-885-5p in GC. Quantitative real-time PCR was used to examine the expression of miR-885-5p in GC. Bioinformatics analysis was used to predict the target of miR-885-5p and confirmed using the luciferase reporter assay. Wound-healing and Transwell assay were conducted to evaluate the biological function of miR-885-5p and malic enzyme 1 (ME1). Western blotting was used to assess molecular changes. Hepatic and lung metastasis models were constructed and used to verify the role of miR-885-5p. We found that the expression of miR-885-5p was significantly downregulated in GC. Overexpression of miR-885-5p inhibited invasion and metastasis of GC in vivo and in vitro, while inhibition of miR-885-5p has the opposite result in vitro. ME1 is a direct target of miR-885-5p, overexpressed in GC, associated with poor prognosis. Overexpression of miR-885-5p negatively regulates ME1 and causes changes in downstream molecules Vimentin and Fibronectin. Our research found that miR-885-5p plays a tumor suppressor gene and could potentially serve as a biomarker and therapeutic target in GC.
Introduction
Gastric cancer (GC) is the fourth prevalent cancer and the third cause of cancer-related death worldwide, resulting in over one million new cases in 2018 and 783,000 mortality (Bray et al., 2018). More than half of new GC cases worldwide occur in East Asia; although standard surgery and adjuvant treatment strategies have been improved and widely used, most patients are often diagnosed at an advanced stage with a high probability of metastasis and recurrence. Therefore, the survival rate is low, with the 5-year survival rate in China being only 35.9% (Allemani et al., 2018).
MicroRNAs (miRNAs) are single-stranded noncoding RNAs with 20–24 nucleotides, which play essential roles in the post-transcriptional level by binding to mRNAs 3′-untranslated region (3′-UTR) and cause degradation or destabilization using the RNA-induced silencing complex. Growing evidence showed that dysregulation of miRNAs could act as a tumor-promoting factor or tumor suppressors, affecting development, differentiation, and metastasis of cancer (Ye et al., 2018). These regulatory effects, in turn, make miRNAs potential biomarkers and therapeutic targets for cancer (Kanwal et al., 2017; Elghoroury et al., 2018).
Lately, many studies revealed that miR-885-5p plays the role of tumor suppressor in various cancers, including hepatocellular carcinoma (HCC) (Zhang et al., 2016), osteosarcoma (Liu et al., 2019), and neuroblastoma (Afanasyeva et al., 2011). In HCC, miR-885-5p significantly inhibited cell proliferation, migration, invasion, and metastasis (Zhang et al., 2016; Xu et al., 2019). Besides, there is evidence that miR-885-5p suppresses tumor growth, migration, and invasion by interfering β-catenin in osteosarcoma (Liu et al., 2019). Moreover, Afanasyeva et al. discovered that miR-885-5p interfered with cell cycle and vitality in neuroblastoma, thereby playing a tumor suppressor role. Recently, we discovered that the expression of miR-885-5p is notably downregulated in GC. However, there are few studies on the biological function of miR-885-5p in GC. The only study on miR-885-5p in GC was the study by Li et al. who found that miR-885-5p as an oncogene regulates the growth, clone formation, and metastasis of GC cells through the target gene YPEL1 (Li et al., 2019b).
Malic enzyme 1 (ME1), also known as cytosolic NADP (+)-dependent malic enzyme and malate dehydrogenase, is a cytosolic protein responsible for oxidative decarboxylation of malate to pyruvate that leads to NADPH production and essential for glutamine metabolism and lipogenesis (Wise and Ball, 1964; Frenkel, 1975; Al-Dwairi et al., 2012). Research on the relationship between ME1 and human cancer has achieved some results recently. Jiang et al. discovered that downregulation of ME1 reciprocally induces Tumor protein 53 (TP53) activation and promotes TP53-dependent tumor progression and metastasis (Jiang et al., 2013). Son and Ying also discovered ME1 regulated by KRAS in multiple cancers (Ying et al., 2012; Son et al., 2013). Besides, the expression of ME1 was upregulated in many human cancers, including GC, breast cancer, colon and rectal cancers (CRC), HCC, lung cancer, laryngeal squamous cell carcinoma, and oral squamous cell carcinoma (OSCC) (Simmen et al., 2020). ME1 promotes growth and invasion in OSCC, HCC, and GC cells (Wen et al., 2015; Lu et al., 2018), but attenuates migration and invasion in nasopharyngeal carcinoma (Zheng et al., 2012). ME1 levels are also used as prognosis and predictive markers in multiple human cancers, including GC, breast cancer, CRC, HCC, lung cancer, and OSCC (Simmen et al., 2020).
Materials and Methods
Bioinformatics analysis
TCGA (The Cancer Genome Atlas) database was used to download miRNA-Seq data, and differential expression analysis was performed using R packages (DESeq2 and EdgeR). The relationship between miR-885-5p expression and the clinicopathological characteristics of GC was analyzed using the online tools cBioPortal and LinkedOmics. The binding site of miR-885-5p in the 3′-UTR was predicted by miRanda, TargetScan, and starBase databases.
Clinical tissues
All the 40 pairs of GC tissues and adjacent nontumor tissue (NAT) samples were collected consecutively from the Department of Gastrointestinal Surgery, Provincial Hospital Affiliated to Shandong University between October 2019 and December 2019 with an average age of 60, including 31 males and 9 females. The samples have been clinically and histopathologically diagnosed according to the WHO criteria, including 20 early-stage and 20 advanced-stage tumors. Informed consent was obtained from the patients for all parts of this study and was approved by the Medical Ethics Committee of Shandong Provincial Hospital, approval number 2019000121. All surgical tissues were quickly deposited in liquid nitrogen after the resection. Total RNA from clinical tissues was extracted and stored at −80°C for further experimental use.
Cell culture and transfection
The human GC cell lines MGC-803, SGC-7901, BGC-823, HGC-27, and human gastric epithelial cell line GES-1 were acquired from the Chinese Academy of Sciences (Shanghai, China). All cells were cultured in RPMI-1640 (Gibco, Gaithersburg, MD) medium with 10% FBS (Gibco) in a pathogen-free incubator at 37°C with 5% CO2. For overexpression and inhibition in vitro experiments, miR-885-5p mimics, mimics control, miR-885-5p inhibitor, and inhibitor control were synthesized by GenePharma (Shanghai, China). The plasmids containing ME1 (pcDNA3.1) and the empty vector were purchased from Genomeditech (Shanghai, China). All cell transfections were performed using Lipofectamine 3000 reagent (Thermofisher, Waltham, MA), following the manual instructions. The efficacy was verified by quantitative real-time PCR (qRT-PCR) assay. Transfected cells were collected after 24–48 h for further experimental analysis.
RNA extraction and qRT-PCR
Total RNAs from surgical tissues and cell lines were isolated with Total RNA Extraction Reagent (Vazyme, China) according to the manual instructions and qualified by NanoDrop 2000 (Thermofisher). Total RNAs were reverse transcribed (RT) to synthesize first cDNA using miRNA cDNA Synthesis Kit by stem-loop (Vazyme, China) and HiScript III RT SuperMix for qPCR (Vazyme, China) and quantified the expression levels using miRNA Universal SYBR qPCR Master Mix and ChamQ Universal SYBR qPCR Master Mix (Vazyme, China) following the manual instructions.
The qRT-PCR primer sequences were as follows: miR-885-5p-RT (5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAGAGGC-3′), miR-885-5p-F (5′-GCGCGTCCATTACACTACCCT-3′), miR-885-5p-R (5′-AGTGCAGGGTCCGAGGTATT-3′); U6-F (5′-GCTTCGGCAGCACATATACTAAAAT-3′), U6-R (5′-CGCTTCACGAATTTGCGTGTCAT-3′); ME1-F (5′-GGATTGCACACCTGATTGTG-3′), ME1-R (5′-TCTTCATGTTCATGGGCAAA-3′); GAPDH-F (5′-AGAAGGCTGGGGCTCATTTG-3′), GAPDH-R (5′-AGGGGCCATCCACAGTCTTC-3′).
Luciferase reporter assay
The wild type of ME1 3′-UTR sequence (ME1-WT) containing miR-885-5p binding sites and the mutant ME1 (ME1-MT) containing mutant binding sites were constructed and cloned in the pmirGLO vector. The miR-885-5p plasmids and reporter plasmids were cotransfected into MGC-803 cells using Lipofectamine 3000 (Thermofisher). Luciferase Reporter Assay System (Promega, Madison, WI) was used to evaluate the bioluminescence activity after 48 h of transfection; all assays were performed at least thrice.
Immunohistochemistry
Immunohistochemistry assay was performed using Paraffin-embedded GC and matched NATs. After the slides were deparaffinized with xylene, graded ethanol solutions were used for rehydration. Tissue slides were then infiltrated in 10 mM citrate buffer and cooled naturally after 10 min in the microwave for antigen retrieval. After the endogenous peroxidase was inactivated, the antigen was blocked with 10% serum; afterward, they were treated with the ME1 antibody (16619-1-AP, 1:200; Proteintech) at 4°C overnight. Those slides were incubated with secondary antibody and washed, followed by staining with diaminobenzidine and hematoxylin. The tissue section digital scanning system was used to scan and image. Three random fields were selected for each slice to perform the semiquantitative analysis. Each field of view was scored for staining intensity and percentage of positive cells, and the overall score was the multiplication of staining intensity score and cell positivity score. (Staining intensity score: 0 for no staining, 1 for pale yellow, 2 for pale brown, and 3 for brown. Cell positivity score: 0 to 5% is 0 points, 6% to 25% is 1 point, 26% to 50% is 2 points, 51% to 75% is 3 points, and >75% is 4 points).
Wound-healing assay
To perform the migration assay, we plated GC cells to six-well plates, cultured until they were at 70% confluence, then transfected them with miR-885-5p mimics and mimics control. Transfected cells were starved overnight then scratched with 200 μL pipette tips to create the wound. Unattached cells were washed away by phosphate-buffered saline (Hyclone, Logan, UT) and were photographed by a microscope at a magnification of 40 × . The wounded monolayers were cultured for an additional 24 h then photographed again with a microscope at 40 × magnification. The migration distance was assessed with ImageJ software (NIH, Washington, DC). Briefly, the average scratch width can be obtained by dividing the area by the height. Subtract the scratch's width at the end of the experiment from the scratch's width at 0 h to get the distance of cell migration. The data from three independent replicates were statistically analyzed using the migration index, which is the migration distance of the test group divided by the migration distance of the control group.
Transwell assay
Transwell assays were undertaken using Transwell chambers (3422; Corning, NY). In brief, in migration assays, a total of 2 × 105 transfected cells resuspended in 250 μL serum-free medium were seeded into the upper chamber, while the lower chamber was filled with 650 μL medium containing 10% FBS to induce the migration. In invasion assays, the upper chambers were coated with 70 μL diluted Matrigel (BD, Franklin Lakes, NJ) and incubated at 37°C for at least 4 h. A total of 2 × 105 transfected GC cells resuspended in 250 μL serum-free medium were seeded into the upper chamber, while the other chamber was filled with 650 μL medium containing 30% FBS. After 24–48 h at 37°C, cells that have not passed through the upper layer of the membrane were wiped gently by cotton swabs. Invaded cells that penetrated the membrane were fixed using 4% formaldehyde solution then stained in 0.1% crystal violet solution. The fixed cells were observed and photographed at a magnification of 100 × or 200 × , and then the number was counted in six random fields. All assays were conducted in triplicate.
Western blotting
Radioimmunoprecipitation assay (RIPA) buffer (high) (Solarbio, China) was used to extract total proteins. After concentration was measured, a total of 20 μg proteins were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene fluoride (PVDF) membranes (0.45 μm; Millipore). Membranes were soaked in 5% skimmed milk in Tris-buffered saline (pH7.4), containing 0.05% Tween 20 (TBST, Servicebio, China). Afterward, membranes were incubated overnight at 4°C with primary antibodies, including anti-ME1 antibody (ab97445, 1:1000; Abcam, China), Anti-Fibronectin antibody (ab2413, 1:1000; Abcam), anti-Vimentin antibody (mAb #5741, 1:1000; CST, China), and anti β-actin antibody (60008-1-Ig, 1:3000; Proteintech). After washing thrice by TBST, membranes were treated by secondary antibody (#7074, #7076, 1:3000; CST) for 2 h at room temperature. At last, the membranes were washed by TBST in triplicate, subsequently treating the membranes with ECL reagent (Millipore Sigma) for chemiluminescence detection. Images were photographed using Amersham Imager 680 (GE, Boston, MA), and optical density was analyzed using ImageJ software (NIH). All Western blotting experiments were repeated at least thrice and were assessed for reproducibility.
Lentivirus production and animal experiments
To establish the hepatic and lung metastasis models, miR-885-5p-expressing stable GC cell lines were constructed. In brief, SGC-7901-Luc and BGC-823-Luc cell lines were constructed using pGMLV-CMV-Luc lentivirus luciferase reporter (Genomeditech). For the liver metastasis experiment, six 6-week-old male nude mice (Charles River, China) were divided into two groups of three each. After anesthesia, a total of 1 × 106 miR-885-5p expressing SGC-7901 cells or miR-ctrl expressing SGC-7901 cells in 120 μL serum-free medium were injected into the spleen. Similar to the hepatic metastasis experiment, six 6-week-old male nude mice were divided into two groups of three each, and a total of 1 × 106 miR-885-5p expressing SGC-7901 cells or miR-ctrl expressing SGC-7901 cells in 120 μL serum-free medium were injected into the tail vein for the lung metastasis experiment. Approximately 28 days later, hepatic and lung metastasis bioluminescence signals were received by Bioluminescence Imaging System (PerkinElmer, Waltham, MA). The mice were then sacrificed after the last imaging, at which time the livers and lungs were harvested. All animal experiments were repeated thrice, and the procedures were approved by the Animal Care Committee of Shandong Provincial Hospital, approval number 20200017.
Statistical analyses
SPSS Statistics 23 (IBM, Armonk, NY) and GraphPad Prism 8 (GraphPad Software, San Diego, CA) were used to perform all statistical analyses. Different values among each group were analyzed using Student's t-test or one-way analysis of variance. The analysis of clinicopathological parameters was performed by Chi-square test or Fisher's exact test. All data in this study were acquired from at least three independent experiments and presented as the mean ± standard deviation. p < 0.05 was considered statistically significant.
Results
miR-885-5p expression is notably downregulated in GC tissues and cell lines
To explore the expression of miR-885-5p in GC, we first used TCGA database and identified that miR-885-5p was notably downregulated in GC tissues (Fig. 1A). To validate this result, we applied RT-qPCR to examine the expression levels of miR-885-5p in GC, including human cell lines MGC-803, SGC-7901, BGC-823, HGC-27, as well as normal gastric cell line GES-1. Moreover, 40 pairs of matched tissues from GC patients were also examined. The results demonstrated that miR-885-5p was notably downregulated in human GC cell lines compared to that in GES-1 (Fig. 1B); similar results were also obtained between human GC tissues and NATs (Fig. 1C).
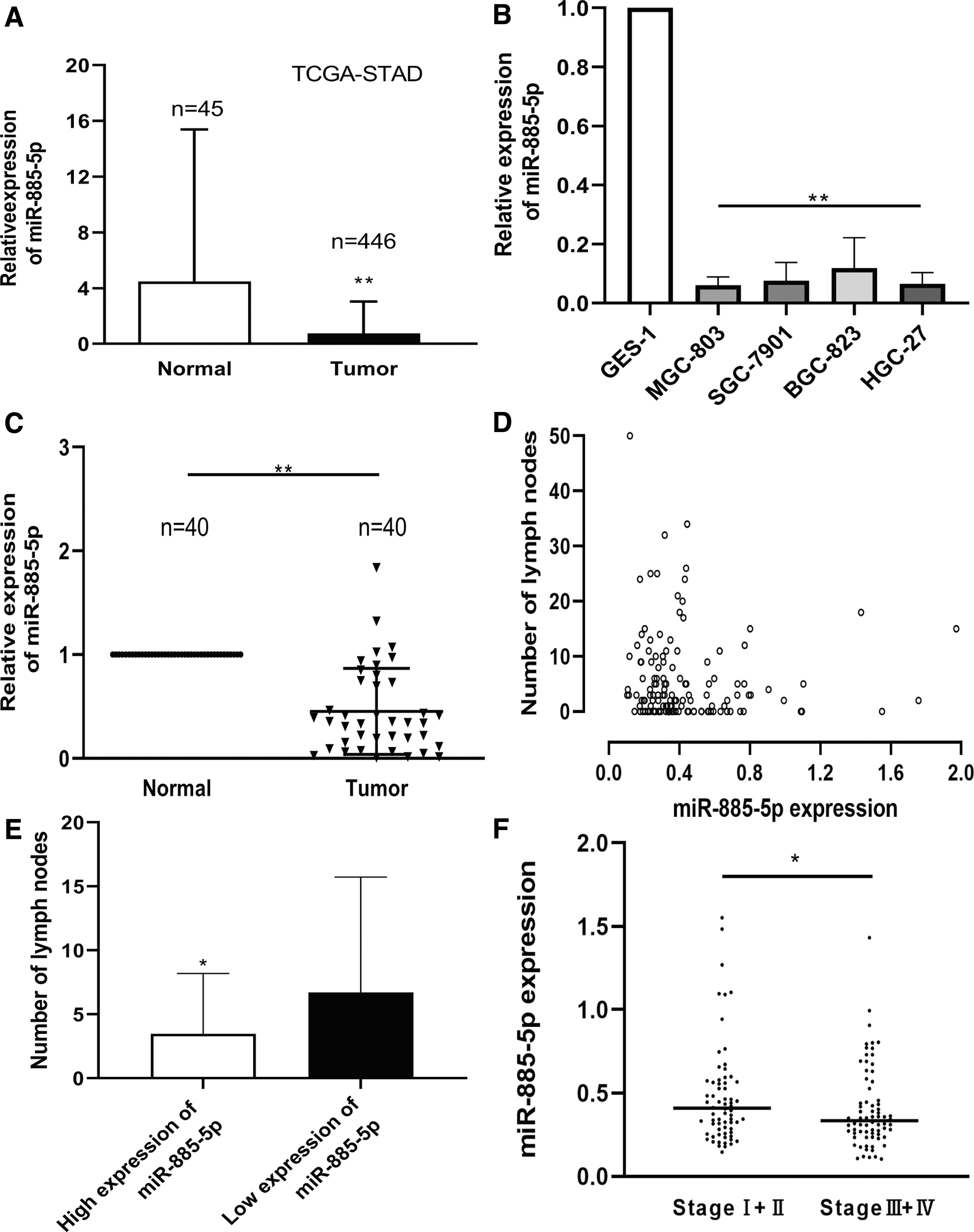
miR-885-5p is downregulated in GC tissues and cell lines.
miR-885-5p expression correlated with tumor invasion and lymphatic metastasis in GC patients
To investigate the biological role of miR-885-5p, we conducted a correlation analysis between miR-885-5p expression and clinicopathological characteristics in tissues from 40 GC patients (Table 1 and Supplementary Table S1). The result implied that the expression of miR-885-5p was highly correlated with the tumor invasion, lymphatic metastasis, and pathological stage (all p < 0.05). Nevertheless, gender, age, venous invasion, tumor size, and historical differentiation were not closely related to the expression of miR-885-5p (Table 1). Through TCGA database, we obtained the number of positive lymph nodes identified by H&E staining together with the expression of miR-885-5p. The results, presented in scatterplot and column chart, are consistent with the findings of our study (Fig. 1D, E). Similar to lymphatic metastasis, the clinical stage data from TCGA further support our findings (Fig. 1F).
Correlation Between miR-885-5p Expression Levels and Clinicopathologic Parameters in 40 Patients with GC
ME1 directly targeted by miR-885-5p
TargetScan, miRanda, and starBase databases were used to find targets of miR-885-5p. After the intersection of these three databases, there were 271 candidate genes. By reviewing the publications, we discovered that among the genes negatively correlated with miR-885-5p expression, ME1 was closely associated with GC. Therefore, we considered ME1 as a potential target (Fig. 2A). To confirm this prediction, we constructed the wild type of ME1 3′-UTR sequence (ME1-WT) containing miR-885-5p binding sites and mutant ME1 (ME1-MT) containing mutant binding sites and cloned them in the pmirGLO vector. Luciferase assay indicated that miR-885-5p suppressed the luciferase activity containing 3′-UTRs of ME1-WT, compared with controls on MGC-803 cell (Fig. 2B). To further confirm this result at the protein level, Western blotting experiments were conducted. After transfection with miR-885-5p, ME1 was inhibited, and overexpression of ME1 reversed the inhibition of miR-885-5 in GC cells (Fig. 2C). Correspondingly, inhibition of miR-885-5p upregulated ME1 expression, while simultaneous knockdown of ME1 inhibited this effect (Fig. 2D). All these effects demonstrated that miR-885-5p directly targets ME1 and inhibits its expression in GC cells.

ME1 is a direct target of miR-885-5p.
ME1 is upregulated in GC cell lines and tissues associated with poor prognosis
After we confirmed that ME1 is a downstream molecule regulated by miR-885-5p, we detected the relative mRNA expression of ME1 in GC. Compared with GES-1, ME1 was overexpressed in GC cell lines (Fig. 3A). Correspondingly, the same results were also obtained in GC tissues and NATs (Fig. 3B). Immunohistochemical analysis in patient-matched tissues also revealed that ME1 expression in GC tissues was higher than NATs (Fig. 3C and Supplementary Table S2). Furthermore, we used TCGA database for correlation analysis, which demonstrated that the expression of ME1 and miR-885-5p was negatively correlated (Fig. 3D and Supplementary Table S3). Kaplan–Meier survival analysis revealed that GC patients with high ME1 expression had a worse survival rate, suggesting that ME1 is associated with poor prognosis (Fig. 3E).

ME1 is upregulated in GC tissues and cell lines and is associated with poor prognosis.
miR-885-5p inhibits migration and invasion in GC cells
To better evaluate the exact biological effects of miR-885-5p in GC, we transfected miR-885-5p and its control into GC cells. The migration assay suggested that the migration ability of miR-885-5p overexpressed cells was inhibited compared to cells transfected with controls (Fig. 4A and Supplementary Table S4). Migration and invasion experiments using Transwell chambers also confirmed this conclusion; with or without Matrigel, GC cells transfected with miR-885-5p showed less migration and invasion than those transfected with control (Fig. 4B). In contrast, miR-885-5p inhibited cells exhibited stronger migration and invasion capability (Fig. 4C). We also conducted a rescue experiment, overexpressed, and silenced ME1 expression in GC cells. Evidence suggested that overexpression of ME1 could block the miR-885-5p's inhibitory effects on migration and invasion in GC cells (Fig. 4B). Correspondingly, silencing ME1 can offset the miR-inhibitor induced migration and invasion (Fig. 4C). To further uncover the mechanism of those phenomena, we applied Western blotting to quantify related protein changes. Results of protein expression indicated that overexpression of miR-885-5p suppressed the expression of Fibronectin and Vimentin in GC cells, while inhibiting the expression of miR-885-5p had the opposite effect (Fig. 5A, B). Moreover, the effects of ME1 and si-ME1 on regulating those genes were also analyzed by Western blotting (Fig. 5C, D).

miR-885-5p inhibits migration and invasion of GC cells.
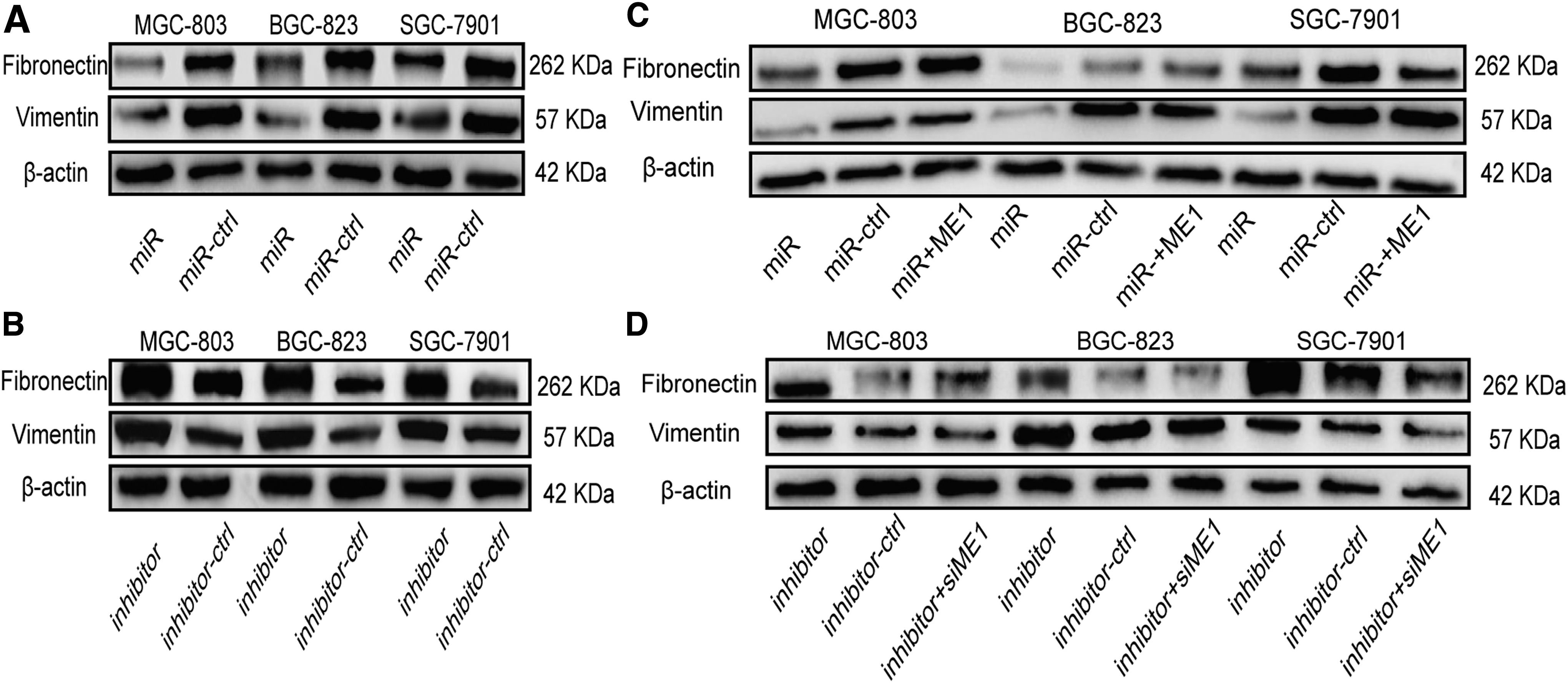
miR-885-5p and ME1 regulate Fibronectin and Vimentin expression in GC cells.
miR-885-5p inhibits GC metastasis in vivo
To evaluate whether miR-885-5p can inhibit the metastasis in vivo, hepatic and lung metastasis models were constructed (Fig. 6A). We used luciferase-labeled GC cells that stably expressed miR-885-5p and injected them into the spleen and tail vein of nude mice, respectively, and compared the metastasis ability with their negative controls, and all experiments were repeated thrice. Replicate experimental results showed that GC cells expressing miR-885-5p clearly show less metastasis in the liver and lung (Fig. 6B, C). In addition, metastatic nodules stably expressing miR-885-5p still showed lower levels of ME1 expression compared to controls (Fig. 6D). Altogether, our results suggest that miR-885-5 exerts potential inhibition of metastasis and invasion in vitro and in vivo by targeting ME1.

Effects of miR-885-5p in vivo.
Discussion
GC is one of the most prevalent and lethal cancers; the high mortality rate is mainly due to its distant metastasis, so it is promising to identify and intervene with the molecules involved in its metastasis. As a post-transcriptional regulatory molecule, miRNA's intricate effects and mechanism in malignant tumors have been studied extensively (Ambros, 2004; Calin and Croce, 2006).
miR-885-5p was dysregulated and serves as an oncogene or tumor suppressor among many malignant tumors (Yang et al., 2017; Li et al., 2020). However, almost no research exists on the biological function of miR-885-5p in GC. The only study on miR-885-5p in GC was the study by Li et al., who found that miR-885-5p was highly expressed in GC, as an oncogene regulates the growth, clone formation, and metastasis through the target gene YPEL1 (Li et al., 2019b), a finding that is contrary to our study. We discovered that miR-885-5p was notably low expressed at the cell and tissue level by comparing GC cells with GES-1 and comparing GC tissues with NATs. We predicted the target gene by bioinformatics methods and validated that ME1 was the direct target of miR-885-5p through luciferase assay. Subsequently, evidence revealed that ME1 was overexpressed in GC at the pretranscriptional and protein level through cell and tissue experiments, and the expression of ME1 and miR-885-5p was negatively correlated in TCGA database. Furthermore, we transfected GC cells with mimics and inhibitors. miR-885-5p overexpression in cells leads to a decrease of ME1 at protein levels. These results proved that miR-885-5p serves as an upstream controller of ME1. In the phenotypic experiment, upregulation of miR-885-5p or silencing of ME1 inhibited migration and invasion, while overexpression of ME1 or suppression of miR-885-5p promoted migration and invasion in GC. We also obtained the same results from animal experiments. Therefore, as a tumor suppressor, miR-885-5p can straightly regulate the expression of ME1 and inhibit the progression of GC in vitro and in vivo.
Furthermore, TCGA database demonstrated that the number of lymph node metastases and pathological stages were statistically significant with the expression of miR-885-5p. Moreover, in our own clinicopathological characteristics analysis, in addition to lymphatic metastasis and pathological stage, tumor invasion was also considered to be statistically significant. Through the verification of experiments, our findings might be used as a novel complement to the complex interaction between miRNA and GC, which can potentially act as a new target for GC treatment.
Vimentin and Fibronectin were also found to be related to the miR-885-5p-ME1 axis in our study. As an essential molecule in the epithelial–mesenchymal transition (EMT) process, Vimentin was related to the invasion and metastasis process in multiple cancers and associated with prognosis and survival in GC (Otsuki et al., 2011; Satelli and Li, 2011; Yin et al., 2018; Paolillo and Schinelli, 2019). Wen et al. proved that ME1 induces the EMT process and is related to the expression of Vimentin in HCC, which was similar to our results in GC (Wen et al., 2015). Fibronectin as a marker of EMT also promoted migration and invasion in cancers, including GC (Chen and Zheng, 2017; Li et al., 2017, 2019a; Xia et al., 2017; Lin et al., 2019), and indicated poor prognosis in GC (Sun et al., 2020).
Conclusion
In summary, our research found that miR-885-5p plays the role of tumor suppressor gene, inhibiting invasion and metastasis by regulating ME1 in GC, and could potentially serve as a biomarker and therapeutic target in GC.
Ethics Statement
Informed consent was obtained from the patients and was approved by the Medical Ethics Committee of Shandong Provincial Hospital, approval number 2019000121. The procedures of animal experiments were approved by the Animal Care Committee of Shandong Provincial Hospital, approval number 20200017.
Footnotes
Authors' Contributions
Conceptualization, Z.J. and L.L.; Funding acquisition, L.L.; Investigation, Z.J. and H.C.; Methodology, Z.J.; Supervision, S.Z.; Validation, H.C. and S.Z.; Writing—original draft, Z.J.; Writing—review and editing, L.L. All authors contributed to the article and approved the submitted version.
Disclosure Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding Information
This work was supported by the Key Research and Development Program of Shandong Province [2019JZZY010104, 2019GSF108146] and the Special Foundation for Taishan Scholars Program of Shandong Province [ts20190978].
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.