
Abstract
The interaction of calf thymus DNA (ct DNA) with anastrozole, which is acknowledged as an antineoplastic drug, has been enquired into in the absence and presence of histone H1, through the means of absorbance, fluorescence, circular dichroism spectroscopy, viscosity, thermal melting, and molecular modeling techniques. In addition, the effects of anastrozole on MCF 7 cell line have been thoroughly investigated. Fluorescence spectroscopy results have indicated that quenching mechanism of ct DNA-anastrozole are known as static quenching procedures, since the Stern-Volmer quenching constant (KSV) seems to face a decrease as the temperature is enhanced; this is a significant evidence for intercalative binding mode of anastrozole with ct DNA. Regarding the ternary system in the presence of H1, the constant of Stern-Volmer quenching was increased as the temperature was heightened. The thermodynamic parameters suggested that the binding could be characterized as exothermic by negative and positive enthalpy and entropy changes in both binary and ternary systems, respectively. It is vital to mention that hydrogen bonds and hydrophobic contributions play significant roles in anastrozole association to ct DNA in the absence and presence of H1. In accordance to the absorption spectroscopy and melting temperature curve outcomes, the binding mode of anastrozole with ct DNA in absence and presence of H1 was indicative of intercalative and nonintercalative bindings, respectively. The viscosity results as binary and ternary systems, which have been elucidated from a sensitive viscometer, have confirmed the fluorescence spectroscopy determinations. The intercalation of anastrozole to ct DNA seemed to be significantly related to an induced reduction in MCF-7 cell proliferation. The molecular modeling results have suggested that anastrozole could bind to H1 in ct DNA-H1 complex in ternary systems, which supports the conclusions that have been obtained from experimental data.
Introduction
Cancer stands as one of the rationales of death throughout the developed countries (Steiner et al., 2016). Although a lot of researches are available on this topic, the reducing rate in cancer deaths is relatively low (Bonneterre et al., 2000). In accordance to anticancer therapy, the specific interaction of molecule with the genomic DNA may be effective in preventing further information retrieval from DNA and thus lead to the arrest of cell division. The photo physical behavior that is linked to DNA binding could be employed for site-directed cleavage of the DNA backbone in regard to photodynamic therapy of tumors and other diseases (Bhadra and Kumar, 2011; Abdulrahman et al., 2019).
Deoxyribonucleic acid is responsible for carrying the main genetic information and leads the biological synthesis of proteins and enzymes through the processes of duplication, transportation, and translation of genetic information. DNA is acknowledged as the intracellular target for most anticancer, antibiotic, and antiviral drugs (Mohammed et al., 2018). Small molecules can bind to DNA by covalent or noncovalent interactions and cause the alteration or inhibition of DNA function (Han et al., 2014; Shi et al., 2015). The research of drug-DNA interaction is very important for discovering the drug function and designing specific DNA-targeted drugs, since it contains a significant relevance to cancer chemotherapy (Hu et al., 2005; Paul and Kumar, 2012; Tanveer et al., 2020).
Small molecules such as drugs can bind to DNA and (DNA-H1) complex by means of covalent or noncovalent interactions (Qais et al., 2017). Therefore, investigating the binding mechanism of DNA and (DNA-H1) complex with small molecules can function as a key role in comprehending their clinical activities (Tanveer et al., 2019).
Most of the antitumor drugs are focused on targeting the DNA in a cell. Considering the biological implications of anastrozole, its interaction with DNA is an important aspect to be assessed in regard to the suppression of tumor growth (Nawaf et al., 2020a).
Anastrozole is the first aromatase inhibitor to display significant survival advantages in the treatment of postmenopausal women with advanced breast cancer, which was put into progress after the prior tamoxifen treatment (Rudolph and Luger, 2019). Although the majority of breast cancers has expressed the estrogen receptor and depends on estradiol (E2) for their growth, the activity of anastrozole is quiet selective (Milani et al., 2009). Aromatase inhibitors are effective in preventing the breast cancer reverse promotion of new contralateral tumors in postmenopausal women. Estrogen stands as a vital agent in breast cancer carcinogenesis and inducing a decrease in its synthesis can cause the reduction of breast cancer risk. Estrogen generation has been driven by the aromatase enzyme, which can convert androgens to estrogens (Gangadhara and Bertelli, 2009).
The DNA of all eukaryotes is packaged into chromatin by its aggregation with histone proteins. There are five main classes of histones, including the core histones H2A, H2B, H3, and H4, which organize the protein components of nucleosome core particles, along with the linker histone H1 (Babić and Serec, 2020a).
Therefore, the main goal of this article would be to apperceive the mode of interaction between anastrozole with calf thymus DNA (ct DNA) and (ct DNA-H1) complex.
In this work, the binding interaction between anastrozole and ct DNA has been enquired into, in the absence and presence of H1, through the utilization of ultraviolet (UV)-visible absorption spectroscopy, fluorescence emission spectroscopy, resonance light scattering (RLS), circular dichroism (CD), viscosity measurement, and molecular modeling (Tanveer et al., 2018). These particular procedures have been performed for the purpose of obtaining detailed information about the binding interaction of anastrozole with ct DNA and ct DNA-H1 complex such as the specific binding site, binding modes, binding constant, effect of anastrozole on conformation of ct DNA, and the interaction forces, among others. This type of research will be efficient to design new types of agents that can target ct DNA and ct DNA-H1 complex as part of their beneficial performance.
We have investigated the nature and strength of anastrozole binding to ct DNA (single strand and double helix strand), which has provided information on the complexion of anastrozole with ct DNA and helped in elucidating the peculiarities of this biologically important complex formation. Therefore, the achieved information from this study will be quiet helpful in understanding the mechanism of anastrozole interaction with ct DNA and should be useful in the development of potential probes of DNA conformation along with new therapeutic reagents for various diseases.
Experimental
Materials
ct DNA, histone H1, anastrozole, ethidium bromide (EB), and acridine orange (AO) have been purchased from Sigma Chemical Company and used without further purification. All the materials have been prepared in Tris-HCl buffer (pH = 6.8) and kept in refrigerator. The Tris-HCl buffer solution (pH = 6.8) consisted of Tris, ethylene diamine tetra acetic acid (EDTA), and NaOH, which were dissolved in distilled water and adjusted to pH 6.8.
The stock solution of ct DNA that was prepared in buffer resulted in a ratio of ultraviolet absorbance at 260 and 280 nm, indicating that the DNA is sufficiently free from protein (Raman et al., 2012; Karthikeyan et al., 2018). The concentration of ct DNA solution was measured with its absorbance at 260 nm and determined by utilizing the molar absorption coefficient of 6600 M−1·cm−1. The stock solution of anastrozole (5 × 10−4 M) was prepared in dimethyl sulfoxide (DMSO). The final concentration of DMSO in the solution of anastrozole was controlled to stay at a number lower than 10% to avoid the conformational charge of ct DNA and the interaction effect of ct DNA with anastrozole. Double distilled water was applied to prepare all stock solutions for DNA binding studies.
The histone H1 solution was prepared by appending 0.0559 g of histone in Tris-HCl buffer, while the concentration of anastrozole was 0.5 mM, which was provided with 0.0014 g of anastrozole powder that has been dissolved in Tris-HCl buffer.
The MCF7 cells (Iranian type Culture Collection, Tehran, Iran) were maintained in 75 cm2 culture flasks at 37°C and in a 5% CO2 humidified atmosphere. The cells were cultured in the Dulbecco's modified Eagle's medium (DMEM) that contained 10% fetal bovine serum (FBS) and 1% penicillin-streptomycin. DMEM and FBS were procured from Sigma Chemicals, while the antibiotic was purchased from Invitrogen. All the subcultures that had been utilized before passage 15 were routinely performed through the application of 0.25% trypsin/0.1% EDTA.
Methods
The absorption spectral data have been recorded on Jasco V-630 spectrophotometer (Jasco, Tokyo, Japan) that was equipped with a temperature cell holder. The fluorescence emission spectra of solutions have been recorded on a Hitachi F-2500 spectrofluorometer with 1.0 cm quarts cell (Tokyo, Japan) from 300 to 500 nm at λex = 271 nm, at 25°C. These fluorescence emission spectra have been measured as the average of three scans and the appropriate blanks that corresponded to the buffer have been subtracted to remove background.
For the purpose of decreasing the inner filter effect, the fluorescence intensities that had been employed were corrected to absorb the excitation light and reabsorb the emitted light through the usage of the following formula:
where Fcor and Fobs stand as the corrected fluorescence intensity and observed fluorescence intensity, respectively, and Aex and Aem would be the absorption of the system at the excitation wavelength and the emission wavelength, respectively. The measurements have been performed by applying anastrozole ([Anastrozole] = 0.5 × 10−4 M) and appending various concentrations of ct DNA and ct DNA-H1 complex.
CD measurements have been performed on Jasco J-815 with 1.0 cm quartz cell (Japan Spectroscopic Company, Tokyo, Japan) at 25°C, in which the scan range varied from 200 to 350 nm with an interval of 1 nm at a scan rate of 100 nm·min−1. Each spectrum was the average of three scans and corrected through the means of the corresponding buffer blanks.
The RLS spectra have been recorded through a Hitachi model F-2500 spectrofluorometer (Tokyo, Japan)
Viscosity measurements have been performed by the application of a viscometer (Ubbelohde, Shanghai, china), which equilibrated at 25°C ± 0.5°C. The values of relative specific viscosity (μ/μ0)1/3, where μ and μ0 are the specific viscosity contributions of ct DNA in the absence (μ0) and presence of the drug (μ), were plotted against [anastrozole]/[ct DNA] and [anastrozole]/[ct DNA-H1] (Xu et al., 2011).
The comparing procedure of quenching effects regarding single-stranded (ss)-ct DNA and double-stranded (ds)-ct DNA has been performed by adding small aliquots (10 mL) of a concentrated ss-ct DNA or ds-ct DNA solution to anastrozole solution, which contained a constant concentration. All the solutions were allowed to equilibrate for 5 min before the measurements were carried out.
MTT assay
Cytotoxicity assays are widely used in in vitro toxicology studies, while the MTT assay is acknowledged as one of the most common employed techniques for the detection of cytotoxicity or cell viability that follows the exposure to toxic substances (Pawlak et al., 2019). The MTT assay was carried out on MCF7 cell lines, which had been provided from Bu Ali Research Institute, to determine the anticancer activity of anastrozole. The MTT colorimetric assay was performed to detect the cell viability after 24 h of exposure to anastrozole (0.001, 0.1, 0.5, and 1 mM). Culture media were removed and then MTT, (3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide), which had been purchased from Sigma, was diluted in culture media and appended to each well of the plate. Afterward, the plates were incubated at 37°C in the CO2 incubator, for the duration of 4 h. After removing the media, the obtained formazan crystals were stabilized by the addition of DMSO. The absorbance was observed and measured at 570 and 630 nm by the employment of an Epoch spectrophotometer.
Molecular modeling
The molecular docking studies were carried out by means of MGLTools 1.5.6 with AutoGrid 4 and AutoDock 4. The structure of the ct DNA and ct DNA-H1 complex was downloaded from the Protein Data Bank with accession number of 1bna and p02259, respectively (
The DNA was enclosed in a box with a number of grid points in x y z directions, 74, 92, and 76, and a grid spacing of 0.375 Å. Lamarckian genetic algorithms, as implemented in AutoDock, were employed to perform the docking computation. A total of 100 runs were accomplished and the best structure with minimum energy was finally selected.
Results and Discussion
Absorption spectra
The UV-visible absorption spectra of the ct DNA, ct DNA-anastrozole, ct DNA-H1, and (ct DNA-H1) anastrozole systems (Fig. 1) were measured separately in Tris-HCl buffer (pH = 6.8). Figure 1 shows the hyperchromicity and blue shift of ct DNA-anastrozole absorption curve than to ct DNA peak that clearly determines the complex formation of ct DNA-anastrozole. It is generally accepted that the binding of small molecules to the base pairs of DNA through intercalation usually results in hyperchromic (or hypochromic) effect and a blue shift (or red shift) of the DNA absorption band (Guo et al., 2007; Sun et al., 2011). Therefore, anastrozole was likely to interact with ct DNA through some of the binding modes. ct DNA-H1 absorption curves in the absence and presence of anastrozole have been also shown in Figure 1. As can be seen, anastrozole enhances the absorbance of ct DNA-H1 curve, which shows the binding between ct DNA-H1 and anastrozole occurs.

UV-Visible absorption spectra of ct DNA, ct DNA-anastrozole, ct DNA-H1, and (ct DNA-H1) anastrozole in Tris-HCl buffer, pH = 6.8. The concentrations of ct DNA and anastrozole were 0.012 and 0.05 mM, respectively. ct DNA, calf thymus DNA; UV, ultraviolet.
Resonance light scattering
The RLS technique, which measures light scattering signals on a common spectrofluorometer, has greatly altered the situation of light scattering in spectral measurements. The RLS spectra were recorded with a Hitachi model F-2500 spectrofluorometer as the excitation and emission wavelengths varied from 220 to 600 nm with Δλ = 0 nm, at room temperature. Generally, RLS is widely applied to investigate the aggregation, size, shape, and distribution of particles in a solution, which is determined in accordance to the following formula:

where n would be the refractive index of the medium, N stands as the molarity of the solution, λ is the wavelength of the incident and scattered light, V would be the molecular volume, and δn and δk are fluctuations in real and imaginary components of the refractive index in regard to the particle, respectively. RLS technique is capable of providing some intelligence into the process that is responsible for the organization of the complex; therefore, the amount of scattering is directly proportional to the volume of each sphere, and thus, larger aggregation would result in greater scattering (Caijuan et al., 2018).
Figure 2A and B illustrate the RLS spectra of ct DNA and ct DNA-H1 complex upon their interaction with anastrozole, which determines that the enhancement of anastrozole concentrations can regularly increase the RLS spectra and indicate the binding of anastrozole to ct DNA in the absence and presence of H1. On the other hand, any alteration in these particular spectra suggests the conformational changes of ct DNA and ct DNA-H1 that supply the binding of other anastrozole moles. As Figure 2C demonstrates the ΔIRLS against anastrozole concentration, it can be observed that there are two sets of binding sites in ct DNA-H1 complex for binding to anastrozole.

Fluorescence spectroscopic studies
Considering its high sensitivity, fluorescence quenching is known as a useful method to monitor the molecular interactions of chemical and biological systems (Tan et al., 2009; Magdalena, 2018). The fluorescence intensity of a compound can quench as a result of existing different molecular interactions in the experiment such as excited state reactions, molecular rearrangements, energy transfer, ground state complex formation, and collision quenching. Dynamic quenching is generally caused by the collision between fluorophore and quencher, whereas the static quenching is generated by the ground-state complex formation between fluorophore and quencher (Lei et al., 2020).
Figure 3A displays the emission spectrum of anastrozole in the absence and presence of varying concentrations of ct DNA. As it can be perceived from Figure 3A, the maximum emission wavelength of anastrozole was measured to 240 nm as the excitation wavelength was at 271 nm. Upon the addition of ct DNA concentration to the fixed concentration of anastrozole, the intensity of anastrozole emission seemed to decrease regularly and one could observe the red shift of emission wavelength (420–430 nm). Moreover, two isosbestic points were present in the emission curves at 380 and 460 nm, indicating that an equilibrium state exists between ct DNA and anastrozole. It was suggested by the outcomes that an interaction has occurred between the ct DNA and anastrozole.
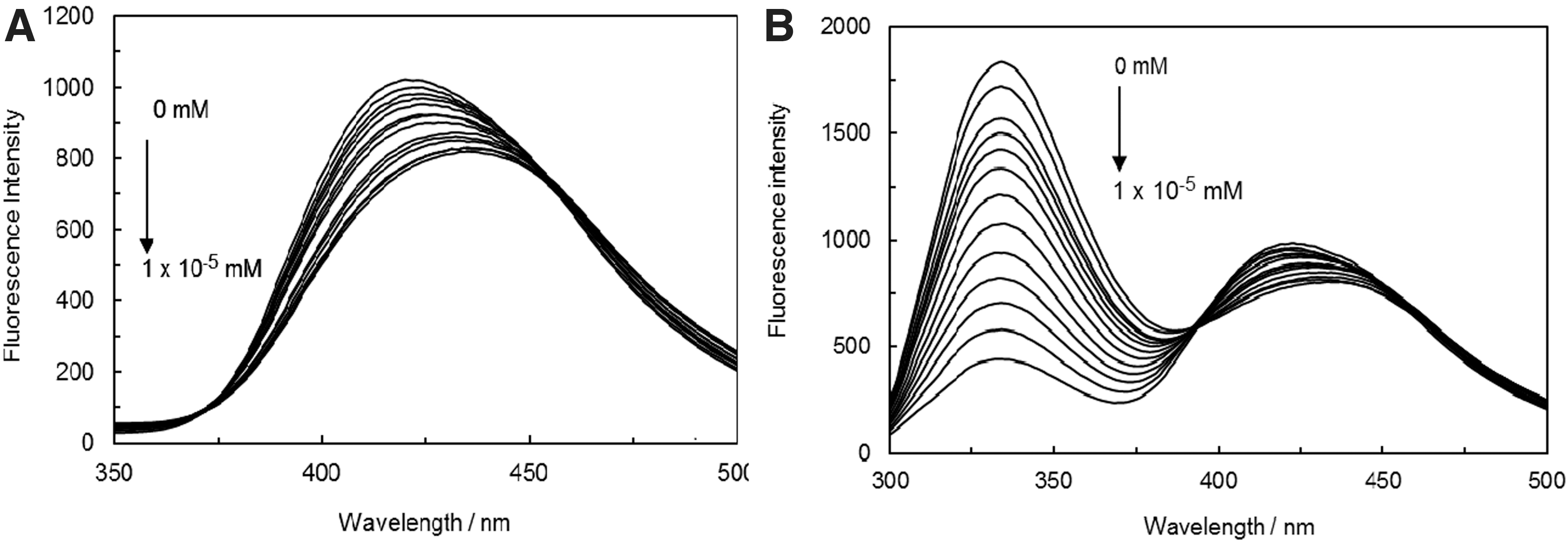
In Figure 3B, we can observe the effect of DNA-H1 solution on anastrozole that has caused a quenching, which indicates the binding of anastrozole along with conformational changes in the ct DNA-H1 complex.
Quenching can occur through different mechanisms, which are usually classified as dynamic and static quenching. Fluorescence quenching is commonly described by the Stern-Volmer equation:
where F0 and F represent the fluorescence intensities in the absence and presence of the quencher, respectively. The kq stands as the fluorophore quenching rate constant, Ksv would be the quenching constant, τ0 is the lifetime of the fluorophore in the absence of quencher (τ0 = 10−8), and [Q] would be the concentration of quencher (Krafcikova et al., 2019).
Dynamic and static quenching can be distinguished by their different dependencies on various temperatures (Chi and Liu, 2011). Regarding the dynamic quenching, the maximum scattering collisional quenching constant of various quenchers is 2 × 1010 L·mol−1·s−1..
Table 1 represents the Ksv values of ct DNA-anastrozole complex in the absence and presence of H1, which were obtained as binary and ternary systems from Stern-Volmer equation at different temperatures. As it can be comprehended from Table 1 throughout the ct DNA-anastrozole complex formation, Ksv values seem to decrease as the temperature is heightened, which suggests that the mechanism of quenching may be static (Fig. 4A). The Ksv values of ct DNA-anastrozole, in the presence of H1 as a ternary system, were observed to decrease as the temperature was heightened, indicating that the behavior of the quenching was dynamic (Table 1 and Fig. 4B). The binding constant (Kb) for the complex formation between ct DNA and anastrozole in the absence and presence of H1 has been measured in accordance to Equation (3), as follows:

Quenching Constants and Binding Parameters for the Interaction of Anastrozole with ct-DNA and (ct-DNA-H1) Complex at Various Temperatures
ct DNA, calf thymus DNA.
Here, F0 and F stand as the fluorescence intensities of the fluorophore in the absence and presence of different quencher concentrations, respectively. In this study, the binding constant of anastrozole to ct DNA, in the absence and presence of H1, has been achieved at various temperatures (refer to Table 1).
The existing interaction forces between small molecules and biomolecules mainly involve hydrogen bonds, van der Waals force, hydrophobic force, and electrostatic interactions. The dominant binding force can be judged through the enthalpy (ΔH0) and entropy changes (ΔS0). The ΔH0, ΔS0, and ΔG0 values in regard to the binding interaction have been calculated by the van't Hoff equation, as follows:
In Equation (5), R would be the gas constant, while the values of ΔH0 and ΔS0 have been obtained from the slope and intercept of the linear van't Hoff plot based on the lnKa versus 1/T (Fig. 5A, B).

Thermodynamic parameters for the interaction of anastrozole with ct DNA, in the absence and presence of H1, are summarized in Table 1. The negative sign for ΔG0 values has revealed that the binding process is spontaneous. Generally, intramolecular forces between macro biomolecules and small molecules include hydrogen bonds and van der Waals, electrostatic, and hydrophobic forces. The Ross's thermodynamic laws state that the functionality of hydrophobic forces becomes quite significant when both ΔH0 > 0 and ΔS0 > 0, whereas in a situation of ΔH0 < 0 and ΔS0 < 0, the main forces are considered to be hydrogen bonding and/or van der Waals forces; however, in the case of ΔH0 < 0 and ΔS0 > 0, the basic molecular forces would be electrostatics (Ross and Subramanian, 1981). The negative values of ΔH0 and ΔS0, for the ct DNA-anastrozole complex as a binary system, have indicated that the binding of anastrozole to ct DNA is driven mainly by hydrogen bond and van der Waals forces. According to the thermodynamic data, the formation of ct DNA-anastrozole complex is preferred by enthalpy and disfavored by entropy; therefore, the perceived negative entropy confirms the intercalative binding mode of anastrozole to ct DNA.
Thermodynamic parameters, in regard to the interaction of ct DNA-H1 with anastrozole as a ternary system, have been listed in Table 1. The positive values of ΔS0 and ΔH0 were frequently taken as proofs for the existence of hydrophobic interactions (Ross and Subramanian, 1981; Bi et al., 2008). The positive value of ΔH0 suggested that the binding process was mainly enthalpy driven through the means of hydrogen bonds (Yuan et al., 2008). Therefore, both hydrophobic interaction and hydrogen bonds might contain vital functionalities in the binding of anastrozole to ct DNA in the presence of H1, while at the same time reveal the groove binding of anastrozole to ct DNA in the presence of H1 as a ternary system. The different values of thermodynamic parameters in binary and ternary systems determine that anastrozole is bounded to both H1 and ct DNA, which uncovers various behavior mechanisms of anastrozole to ct DNA in the binary and ternary systems. Therefore, H1 plays an important role in ct DNA-anastrozole complex formation and causes the changes of the binding site and force type in ct DNA-anastrozole interaction.
EB and AO are the most sensitive fluorescence probes that contain a planer structure, which binds to ct DNA through an intercalative mode (Rich et al., 1984; Fasman, 2013). Although the fluorescence emission of EB and AO seems to increase after an interaction with ct DNA, they end up being decreased in the binding sites of ct DNA that is available for EB and AO, which in return lower the fluorescence intensity of the ct DNA-EB and ct DNA-AO complexes. Figure 6A and B display the fluorescence emission on the spectra of EB in binary and ternary systems, while the AO spectra, in the absence and presence of H1, have been illustrated in Figure 7A and B. As one can perceive from Figures 6A and 7A, through the addition of anastrozole to ct DNA-EB and ct DNA-AO complexes, the fluorescence emission intensity decreases regularly toward the EB and AO fluorescence emission peaks.


The displacement of bound EB and AO that compete for the same site on ct DNA is reflected from a decrease in the intensity, which is due to the addition of competing anastrozole. Figures 6A and 7A show the intercalator mode of anastrozole upon interaction with ct DNA in the absence of H1 as a binary system.
Figures 6B and 7B represent the fluorescence emission on the spectra of EB and AO with ct DNA-H1 complex, and the effect of appending anastrozole to EB and AO has bound ct DNA-H1. There is no significance in regard to EB and AO by anastrozole, which would reflect the absence of an intercalator mode of binding, while the groove binding mode could be associated with ct DNA in the presence of H1.
The ionic strength effect study
In this article, NaCl and KI have been employed for the purpose of controlling the ionic strength of solutions. Regarding the electrostatic binding mode, Na+ and K+ ions have inclined to bind with the phosphate groups of DNA, which resulted in weaker bindings of small molecules with DNA; nevertheless, for the groove binding mode, the minor groove of DNA, after the enhancement of NaCl and KI, seemed to be narrow and deeper as the double-strand helix of DNA converged nonlinearity. It can be determined by Figure 8A and B that the absorbance of ct DNA-anastrozole has altered when the concentrations of NaCl and KI ranged from 0 to 0.5 M and 0 to 0.5 × 10−3 M, respectively. This fact indicates that a significant electrostatic binding interaction has been induced between ct DNA and anastrozole. On the other hand, in the presence of H1, the absorbance of ct DNA-anastrozole complex did not change, which suggests that the hydrophobic forces seemed to be more powerful than the electrostatic interaction. The investigation of ionic strengths on ct DNA-anastrozole, in the absence and presence of H1, has proved that the linker histone plays an important role in the interaction behavior between ct DNA and anastrozole.

CD measurements
CD is a reliable tool for better understanding the conformational changes in DNA upon its interaction with small molecules. It is known that the intercalation of linear or flat aromatic molecules into the double-stranded DNA can induce large chirality alterations and consequently have severed effects on their CD spectra (Fasman, 2013). The CD spectrum of free DNA has displayed a negative band at 247 nm, which has been caused by helicity, while a positive band has been observed at 279 nm due to the base stacking, which is a characteristic of DNA in the right-hand B form (Rafael et al., 2017).
Herein, the negative band at 247 nm has increased, while the positive band at 297 nm has faced a decrease as the anastrozole concentration has been appended to ct DNA (Fig. 9A). This observation indicates that the helicity of ct DNA has decreased with the enhancement of anastrozole concentration. It has also supported the conformational changes of ct DNA upon interaction with anastrozole, while the possibility of the induced interaction was intercalative. On the other hand, the binding of anastrozole to ct DNA in the presence of H1 has revealed a large decrease in the ellipticity of the 240 nm peak (Fig. 9B). The substantial CD perturbation at 240 nm has uncovered a clear and stronger stacking interaction of anastrozole to ct DNA in the presence of H1 as a ternary system. The difference between chirality values and wavelength seems to be dependent on the occurred changes in ct DNA-anastrozole complex in the absence and presence of H1, which clearly designates the significant functionality of H1 in the creation of interaction behavior between ct DNA and anastrozole. On the other hand, comparatively larger CD perturbation at the 247 nm peak at binary and 240 nm peak at ternary systems has revealed that anastrozole is capable of binding to H1 as well.

Viscosity investigation
Viscosity measurements have been performed to further clarify the binding nature of interaction between anastrozole and ct DNA in the absence and presence of H1. A classical intercalation binding can lead to an increase in the DNA viscosity due to the induced lengthening of DNA helix as the space between the adjacent base pair is widened enough to accommodate the binding ligand; on the other hand, the electrostatic or groove binding interactions can typically cause inconspicuous changes in the DNA viscosity (Bi et al., 2006; Ma et al., 2013).
In this study, the relative viscosities of DNA have been plotted versus the binding ratio r (r = [anastrozole]/[ct DNA] and r = [anastrozole]/[ct DNA-H1]). The results that are demonstrated in Figure 10 indicate that the relative viscosity of ct DNA has been obviously measured with the continuous alteration of anastrozole, which further proves the capability of its molecules in bounding to ct DNA through an intercalative binding. As it is displayed in Figure 10, there seemed to be a decreasing change in the relative viscosity of ct DNA upon its interaction with anastrozole in the presence of H1. It must be mentioned that the binding mode of anastrozole with ct DNA in the presence of H1 was a nonintercalative one, which makes it possible to be a groove binding.

Effect of increasing amounts of anastrozole on the viscosity of ct DNA (closed circles) and ct DNA-H1 complex (open circles).
Comparison of the effects of ssct DNA with ds-ct DNA
The ss-ct DNA solution has been prepared in accordance to the literature method (Ye and Ju, 2003). If there had been an electrostatic interaction, the quenching effect on the drug and ds-ct DNA should have stayed the same (interaction with phosphate groups). In addition, if there had been a groove binding, the quenching effect on the drug would have been strengthened when compared to the ds-ct DNA. If there had been an intercalating binding, the effect on the drug would have been reduced in contrast to ds-ct DNA. As it can be observed from Figure 11A and in regard to anastrozole, both ss-ct DNA and ds-ct DNA have the ability to quench the fluorescence intensity, but ds-ct DNA seem, to contain a stronger pull. On the other hand, on a situation that involves the ct DNA-anastrozole complex formation in the presence of H1, the enhancement effect of ds-ct DNA would be weaker compared with ss-ct DNA (Fig. 11B). Therefore, the interaction between ct DNA and anastrozole in the absence and presence of H1 as binary and ternary systems has been intercalative and in the mode of groove binding, respectively.

Melting studies
Monitoring the alterations in the melting temperature (Tm) of DNA stands as an efficient method to recognize the binding mode of small molecules and DNA. Previous investigations have indicated that the intercalation binding can increase the stability of DNA helix and cause an increase in its Tm up to about 5–8°C, while the groove binding is not able to induce any obvious increase in the Tm (Mallappa et al., 2021). Figure 12 illustrates the changes in absorbance at 260 nm in regard to ct DNA, in the absence and presence of anastrozole. As it is shown in Figure 12, the Tm of ct DNA is about 61°C, whereas the Tm of ct DNA-anastrozole system is about 65°C. These melting experiments strongly support the intercalation of anastrozole into the ct DNA helix. In contrast, the Tm of ct DNA-anastrozole complex, in the presence of H1 as a ternary system, is about 65°C. The interaction between ct DNA and anastrozole in the absence and presence of H1 cannot change the Tm value, which suggests that the stability of ct DNA-anastrozole was not altered. Thus, the results have revealed that the binding mode of anastrozole to ct DNA in the presence of H1 is a groove binding.

Melting curves of ct DNA and ct DNA-H1 in the absence and presence of anastrozole (open circles: ct DNA in absence of anastrozole, closed circles: ct DNA-anastrozole, open diamonds: [ct DNA-H1] anastrozole).
Cytotoxic effect of anastrozole on cancer cell lines, in vitro study using MTT assay
The cytotoxic effect of anastrozole has been determined within MCF7 and the analyses have been carried out through the utilization of MTT method for a period of 24 h. As it is displayed in Figure 13, the MCF7 cells have been treated with various concentrations (0.001, 0.1, 0.5, and 1 mM) for 24 h. The most suitable cytotoxic effect is related to the 0.5 mM concentration, which can be inhibited by cancer cells. The values of the inhibitory concentration at 50% effect level (IC50) have been reported to be 25.16.

Plots of cell viability % versus increasing of the anastrozole (0, 0.001, 0.1, 0.5, and 1 mM) displayed after 24 h.
Molecular modeling
Molecular modeling has been successful in illustration of the interaction of ligands with macromolecules. After examining the empirical observations, we have utilized the molecular modeling of laboratory accuracy. The molecular docking technique is an attractive scaffold for better comprehending the drug-DNA interactions for the purpose of rational drug design and discovery (Rohs et al., 2005).
In this study, the docking calculations have revealed the existing interaction between anastrozole and DNA, in which anastrozole approaches both the minor grooves and intercalating sites of DNA. In accordance to the docking results, anastrozole prefers to bind to the minor groove of G-C rich B-DNA and mainly through the means of van der Waals and hydrogen bonding forces. Besides, it penetrates the intercalating site by means of its cyanide functional group. Intermolecular interactions between DNA and anastrozole are demonstrated in Figure 14A. The experimental free energy of binding (ΔG0) between anastrozole and ct DNA has been observed to be −25.76 kJ·mol−1 and the computational outcome seems to be near −34.54 kJ·mol−1 as well, which are comparable. Among 100 docking runs between DNA-H1 and anastrozole, most of the models were able to bond to both histone protein and DNA groove binding site. Apparently, in the presence of histone, anastrozole mainly prefers to bind into pocket, which is made of Ser, Lys, Ile, and Tyr (Fig. 14B). In DNA-H1 clusters with the lowest bonding energy, besides the van der Waals forces, there has been a hydrogen bond between lysine and the N of anastrozole. Also, the chemical structures of anastrozole have been shown in Figure 14C. Experimental results have suggested that ΔG0 = −48.75 kJ·mol−1, while the docking result has come up with −51.20 kJ·mol−1. Based on previous docking and spectroscopic results (Nawaf et al., 2020b; Tanveer et al., 2021), the molecular docking studies should reinforce the experimental ones. The type of interactions (van der Waals forces) and the active sites seem to be parallel in both studies. Although anastrozole does not have a typical planner structure of the intercalators, however, this drug can disturb the helicity of double-strand DNA and make interactions with the nucleotides in the intercalating sites.

Conclusion
The interaction of ct DNA with anastrozole, an antineoplastic medicine, has been enquired into in the absence (binary) and presence of histone (ternary), through the means of absorbance, fluorescence, CD spectroscopy, viscosity, thermal melting, and molecular modeling techniques. Besides, the effect of anastrozole on MCF 7 cell line has been thoroughly investigated.
The binding constants and thermodynamic values for anastrozole interactions with ct DNA and ct DNA-H1 have been investigated through fluorescence spectroscopy. The experiments reinforced the theory of potent electrostatic interactions in the binary and ternary systems. The research was continued by displacement studies, which was demonstrated in the binary system; anastrozole can bind to the intercalator site, while in the presence of H1, it prefers groove binding modes. The viscosity results and CD spectroscopy studies have confirmed the fluorescence spectroscopy determinations. The intercalation of anastrozole to ct DNA seemed to be significantly related to an induced reduction in MCF-7 cell proliferation. The molecular modeling results have suggested that anastrozole could bind to H1 in ct DNA-H1 complex in ternary systems, which supports the nonintercalative binding mode that has been obtained from experimental data. The results of this study can be guidance for future drug design and drug interaction studies.
Footnotes
Authors' Contributions
N.Z.-F., Z.A.-T., and J.C. designed the research study. N.Z.-F., A.S.-R., P.M., N.N., F.H., and H.R.R. performed the research and collected the data. Z.A.-T. and J.C. analyzed the data. N.Z.-F. and Z.A.-T. wrote the initial draft of the article. M.R.S. and J.C. revised the article. All authors discussed the results and contributed to the final article.
Acknowledgment
The authors thank Dr. Ljungburg for the English editing.
Disclosure Statement
The authors declare that they have no conflict of interests with the concern of this article.
Funding Information
The financial support of the Research Council of the Mashhad Branch, Islamic Azad University, and Mashhad University of Medical Sciences is gratefully acknowledged.