
Abstract
Chemoresistance is one of the major obstacles encountered in ovarian cancer (OC) therapy. Long noncoding RNA PART1 has been reported to be involved in the tumorigenesis of several types of cancers. However, the biological role of PART1 in the chemoresistance of OC is still unclear. In this study, it was found that the expression levels of PART1 and CHRAC1 were increased and miR-512-3p expression was decreased in cisplatin (DDP)-resistant OC cell lines. The depletion of PART1 enhanced the DDP sensitivity of DDP-resistant OC cells, as indicated by the inhibition of cell proliferation, migration, and invasion, and promotion of cell apoptosis. In the upstream mechanism exploration, we discovered that PART1 was induced by YY1 transcription factor. Moreover, it was identified that miR-512-3p was a target of PART1, and PART1 regulated the DDP resistance of OC through miR-512-3p. In addition, we screened the candidate genes of miR-512-3p., and confirmed that CHRAC1 was the downstream gene of miR-512-3p. Furthermore, the knockdown of CHRAC1 inhibited proliferation, migration, and invasion, and accelerated apoptosis of DDP-resistant OC cells, which was counteracted after the inhibition of miR-512-3p. Finally, we observed that PART1 regulated the expression of CHRAC1 through miR-512-3p. In conclusion, we demonstrated that YY1-induced PART1 accelerated DDP resistance of OC through miR-512-3p/CHRAC1 axis, suggesting PART1 may be a promising therapeutic target for DDP-resistant OC patients.
Background
Ovarian cancer (OC) is one of the most common gynecological malignancies, with ∼239,000 new diagnosed cases and 140,000 deaths per year (Banno et al., 2014; Zhao et al., 2020). Owing to the lack of effective screen methods (Ozols et al., 2004), the majority of OC patients were diagnosed at advanced stage, and the 5-year-survival rate of OC patients is only about 15–30% (Dillman et al., 2015). Nowadays, surgery combined with chemotherapy is the primary therapeutic method for OC, and cisplatin (DDP) is considered as a frontline chemotherapeutic agent (Tan, 2016). However, the acquisition of DDP resistance is one of the major obstacles for OC therapy in the clinic (Wang et al., 2018). Thus, it is of great significance to elucidate the underlying molecular mechanism of DDP resistance in OC.
Long noncoding RNAs (lncRNAs) are a type of noncoding RNAs longer than 200 nucleotides (nts) in length (Zhang et al., 2019), and have been demonstrated to play a vital role in a variety of biological processes, including cell proliferation, differentiation, apoptosis, and autophagy (Zhao and Hong, 2021). In addition, lncRNAs-mediated drug resistance in diverse cancers has been widely discussed in recent years (Chen et al., 2017). For instance, lncRNA NCK-AS1 promoted drug resistance of osteosarcoma cells to DDP by regulating the expression of miR-137 (Cheng et al., 2019). Silence of lncRNA NNT-AS1 restrained drug resistance of DDP to cervical cancer cells through regulating miR-186/HMGB1 axis (Liu et al., 2020). PART1 was reported to function as a vital contributor to the tumorigenesis of various cancers, such as breast cancer (Wang and Xu, 2020), hepatocellular carcinoma (Zhou et al., 2020), and colorectal cancer (Zhou et al., 2020), which can be an independent prognostic factor for survival. In addition, recent studies indicated that PART1 induced chemoresistance in esophageal squamous cell carcinoma (Kang et al., 2018), non-small cell lung cancer (Chen et al., 2020), and breast cancer (Zhang et al., 2020). Nevertheless, the involvement of PART1 in the DDP resistance of OC has not been clarified.
MicroRNAs (miRNAs) are a class of small noncoding RNAs with a length of 20–22 nts (Xiao et al., 2018). Emerging evidence has revealed the crucial regulatory role of miRNAs in the chemoresistance of OC. For example, the overexpression of miR-335 suppressed progression and promoted chemotherapy sensitivity of OC (Liu et al., 2018). miR-142 restrained DDP resistance and proliferation of OC by regulating SIRT1 (Gao et al., 2018). miR-199a contributed to the DDP sensitivity of OC through regulating ITGB8 (Cui et al., 2018). miR-512-3p has been shown to have cancer-inhibiting effects in several cancers, such as prostate cancer (Huo et al., 2020), breast cancer (Mohamadzade et al., 2021), and non-small cell lung cancer (Zhu et al., 2015). However, the function of miR-512-3p implicated in the DDP resistance of OC remains largely unclear.
In this study, we mainly focused on the biological function of PART1 in DDP resistance of OC, and the results demonstrated that YY1-induced PART1 regulated the sensitivity of OC to DPP through miR-512-3p/CHRAC1 axis. These findings might provide a novel treatment strategy for DDP-resistant OC patients.
Materials and Methods
Cell lines and culture
Human OC cell lines (CAOV3 and A2780) were obtained from American Type Culture Collection (Manassas, VA) and cultured in RPMI-1640 medium (Invitrogen; Thermo Fisher Scientific, Inc.), including 10% FBS at 37°C with 5% CO2. DDP-resistant OC (CAOV3/DDP and A2780/DDP) cells were established by continuous exposure to progressively increasing DDP concentrations for >6 months. DDP-resistant OC cells were cultured in RPMI-1640 medium containing 2 μg/mL DDP and 10% FBS at 37°C in a humidified incubator with 5% CO2. This study was approved by Ethics Committee of the Fourth Affiliated Hospital of Jiangsu University.
Cell transfection
The short hairpin (sh) RNAs targeting PART1 (shPART1; 5′-UCGCGGUAGCUGAAUCAUGCC-3′) and its negative control (shNC; 5′-GAUACGUUCAAUAGGGAUCCU-3′), miR-512-3p mimics (5′-GACAGUCCUCUGCAAAGAGUA-3′) and its negative control (NC mimics; 5′-UAGUUGAAGCUUGACCGUAGU-3′), and miR-512-3p inhibitor (5′-GAUCGGACUGUUAGCUAGCUU-3′) and its negative control (NC inhibitor; 5′-GAUCGAUCGCCAGCCAGGUCC-3′) were obtained from GenePharma (Shanghai, China). The overexpression of PART1 or CHRAC1 plasmid (PART1 or CHRAC1) was conducted by inserting the full-length PART1 or CHRAC1 sequences into pcDNA3.1 vector. All plasmids and oligonucleotides were transfected into CAOV3/DDP and A2780/DDP cells using Lipofectamine 2000 (Invitrogen, Carlsbad, CA).
Reverse transcription quantitative real-time PCR
Total RNA from OC cells was extracted with TRIzol reagent (Invitrogen). Then cDNAs were generated by Prime Script RT reagent kit (Takara, Dalian, China). The PCR was carried out by SYBR Premix Ex Taq II (TaKaRa) on ABI 7500 real-time PCR system (Applied Biosystems, USA). The relative gene expression was calculated using the 2 −ΔΔCT method. GAPDH and U6 were used as internal controls for PART1, YY1, CHRAC1, and miR-512-3p. The following primers were used for analysis: PART1: 5′-AAGGCCGTGTCAGAACTCAA-3′ (Forward), 5′-GTTTTCCATCTCAGCCTGGA-3′ (Reverse); miR-512-3p: 5′-AAGTGCTGTCATAGCTGAGGTC-3′(Forward), 5′-TTCTCCGAACGTGTCACGTTT-3′ (Reverse); CHRAC1: 5′-GTTAGGCTTGACGGGGTCTCGTCG-3′ (Forward), 5′-GTAATACACGTGCACATCATAGAC-3′ (Reverse); GAPDH: 5′-GGTGGTCTCCTCTGACTTCAA-3′ (Forward), 5′-GTTGCTGTAGCCAAATTCGTTGT-3′ (Reverse); U6: 5′-CGCTTCGGCAGCACATATACTA-3′ (Forward), 5′-CGCTTCACGAATTTGCGTGTCA-3′ (Reverse).
Dual luciferase-reporter assay
The YY1 target sequences (wild-type or mutant) within PART1 promoter were synthesized and inserted into the pGL3-basic vector, and co-transfected into DDP-resistant OC cells with the YY1 overexpression plasmid. Besides, wild-type and mutant sequences of PART1 and CHRAC1 were cloned into the pGL3 vector. Subsequently, these constructed vectors were co-transfected with NC mimics and miR-512-3p mimics into DDP-resistant OC cells. The relative luciferase activities were measured using a dual luciferase-reporter assay system (Promega Corporation, Madison, WI, USA).
CCK-8 assay
For IC50 determination, the transfected cells (1 × 104 cells/well) were seeded into 96-well plates and treated with a range of concentrations of DDP (0, 2, 4, 6, 8, and 10 μg/mL) for 48 h. Afterward, cells were incubated with 10 μL CCK-8 reagent (Beyotime, Haimen, China) for 4 h. The absorbance at 450 nm was measured on a microplate reader. The IC50 value was calculated according to the relative survival curve. To evaluate the cell viability, transfected CAOV3/DDP and A2780/DDP cells (1 × 104 cells/well) were seeded into 96-well plates and treated with 4 μg/mL DDP, followed by CCK-8 assay to evaluate cell viability at indicated time point (0, 24, 48, 72, and 96 h).
Colony formation assay
CAOV3/DDP and A2780/DDP cells were plated in six-well plates and incubated in RPMI-1640 with 10% FBS. After 14 days, the cells were fixed and stained with 0.1% crystal violet. colonies with >50 cells were counted manually.
Chromatin immunoprecipitation assay
The EZ ChIP™ Chromatin Immunoprecipitation Kit (Millipore, Billerica, MA) was used for chromatin immunoprecipitation (ChIP) assay. The OC cells were cross-linked with 1% formaldehyde and were sonicated on ice to create 200–500 bp fragments. Subsequently, the chromatin was incubation with anti-YY1 antibody or IgG as a negative control overnight at 4°C. The precipitated chromatin DNA was recovered and analyzed by reverse transcription quantitative real-time PCR (RT-qPCR).
TUNEL assay
After cells were fixed with 4% paraformaldehyde and permeabilized in 0.1% Triton X-100, cells were then incubated with the TUNEL reaction mixture. Finally, the fluorescent reaction was counterstained with DAPI to stain the nucleus for 10 min at room temperature. Images were captured from four fields of view with a fluorescence microscope.
Wound-healing assay
CAOV3/DDP and A2780/DDP cells (5 × 104 cells per well) were cultured in serum-free medium in six-well plates. After the cells reached 90% confluence, a 200-μL pipette tip was used to generate scratches. The wound width at 0 and 24 h was observed under a light microscope.
Transwell assay
CAOV3/DDP and A2780/DDP cells in serum-free DMEM were placed in the upper chamber precoated with Matrigel (BD Biosciences), and DMEM supplemented with 10% FBS was added into the lower chamber. After incubation for 48 h at 37°C, cells in the lower chamber were fixed with 4% methanol and stained with 0.1% crystal violet. Finally, the invaded cells were counted using a light microscope.
RNA immunoprecipitation
RNA immunoprecipitation (RIP) assay was performed using EZ-Magna RIP Kit (Millipore). The cells were lysed and incubated with A/G magnetic beads conjugated with Ago2 and IgG. The RNA was extracted from the immunoprecipitation complex and the enrichment levels of PART1 and miR-512-3p were measured using RT-qPCR.
Western blot
The isolation of total protein was performed by using RIPA buffer (Beyotime, Shanghai, China). The 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was applied to separate the protein samples, which were then transferred onto polyvinylidene fluoride (PVDF) membranes. Then, the membranes, blocked for 2 h with 5% fat-free milk, were incubated with primary antibodies against YY1 (ab109228; Abcam, USA) or GAPDH (ab8245; Abcam, USA). After washing, the membranes were incubated with secondary antibody (Abcam, USA). Finally, enhanced chemiluminescence detection (ThermoScientific, Waltham, MA, USA) was used to visualize the protein signals.
Statistical analysis
All the experiment had been repeated at least three times and the data were shown as the mean ± standard deviation. The Student's t-test or analysis of variance followed by Tukey's post hoc test was used to analyze the difference between two groups or among the multiple groups. p < 0.05 was considered as statistically significant.
Results
PART1 was highly expressed in in DDP-resistant OC
Based on the TCGA database, PART1 was highly expressed in OC samples (Fig. 1A). To determine PART1 expression pattern in DDP-resistant OC cells, DDP-resistant CAOV3 cell line (CAOV3/DDP) and DDP-resistant A2780 cell line (A2780/DDP) were established. The IC50 value of DDP in CAOV3/DDP and A2780/DDP cells was significantly increased compared with their parental cells (Fig. 1B, C). In addition, PART1 expression was strikingly upregulated in DDP-resistant OC cells compared with that in the corresponding parental cell lines (Fig. 1D). Collectively, the expression of PART1 was significantly upregulated in DDP-resistant OC cell lines.
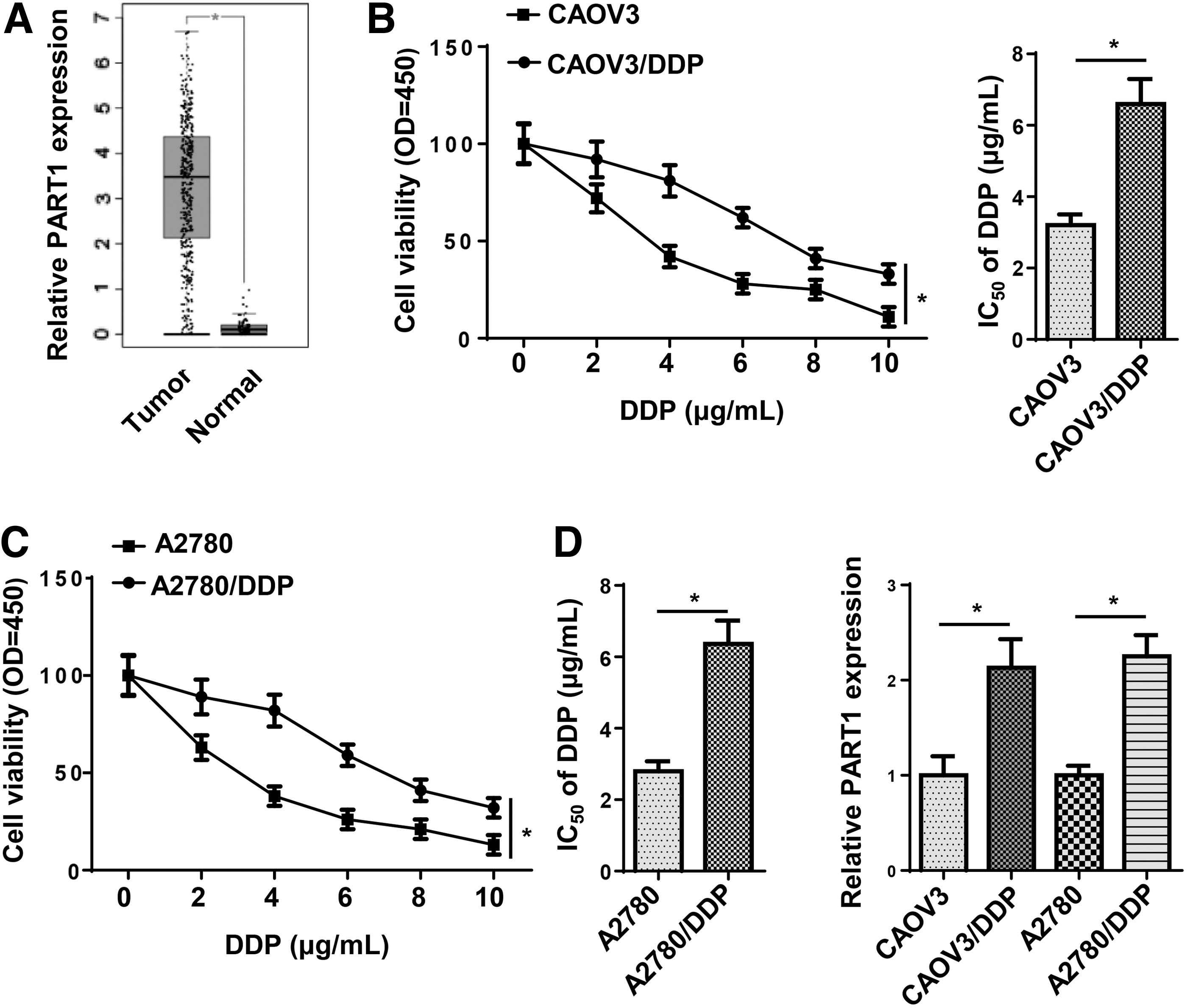
PART1 was highly expressed in in DDP-resistant OC.
Knockdown of PART1 inhibited the DDP resistance of OC
To investigate the biological role of PART1 in DDP-resistant OC cells, shNC or shPART1 was transfected into CAOV3/DDP and A2780/DDP cells. RT-qPCR indicated that the expression of PART1 was significantly downregulated in DDP-resistant OC cells transfected with shPART1 (Fig. 2A). CCK-8 and colony formation assays revealed that PART1 silence remarkably inhibited the proliferation of CAOV3/DDP and A2780/DDP cells (Fig. 2B, C). Moreover, the interference of PART1 suppressed the migration and invasion and increased apoptosis of DDP-resistant OC cells (Fig. 2D–F). These results demonstrated that depletion of PART1 enhanced chemosensitivity of DDP-resistant OC cells.
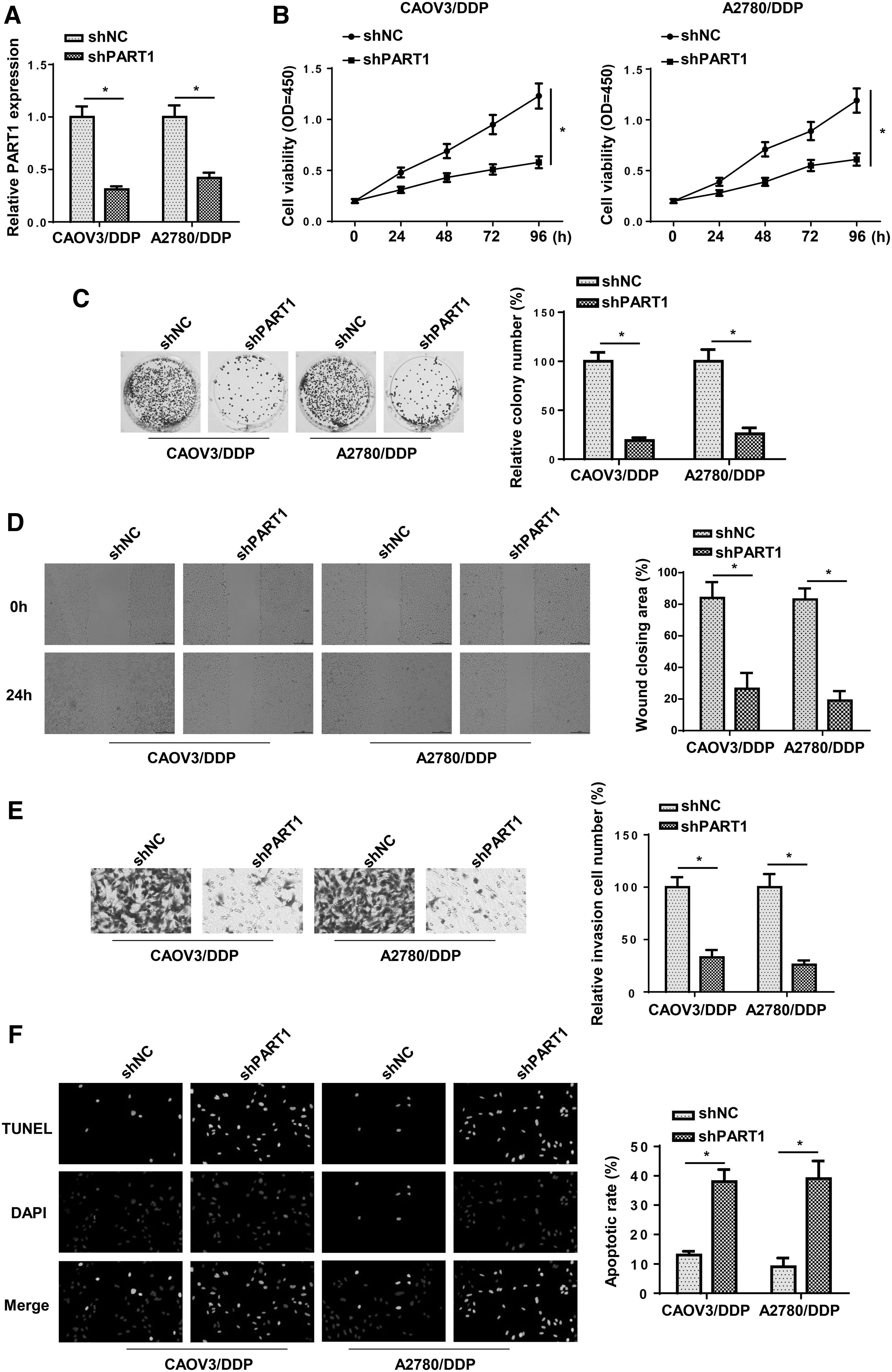
Knockdown of PART1 inhibited the DDP resistance of OC.
YY1 upregulated the expression of PART1 in OC cells
Based on the earlier results, we demonstrated the PART1 knockdown decreased the chemoresistance of OC cells. Therefore, we further investigated upstream regulatory mechanism of PART1 in OC. Increasing studies reported that YY1 transcription factor was involved in the regulation of lncRNA expression [13]. Hence, we wondered whether YY1 could upregulate PART1 at a transcription level. Through JASPAR and UCSC, we obtained the binding motif of YY1 to PART1 promoter (Fig. 3A). Then RT-qPCR demonstrated that YY1 mRNA levels were significantly upregulated or downregulated in OC cells transfected with pcDNA3.1-YY1 or shYY1 (Fig. 3B). Meanwhile, we found that the expression of PART1 was increased by the overexpression of YY1 and decreased by YY1 knockdown (Fig. 3C). In addition, ChIP assay revealed there was a strong binding affinity of YY1 and PART1 promoter in site 2 region (Fig. 3D). Subsequently, we mutated the site 2 of YY1 in PART1 promoter and performed luciferase reporter assay. The results indicated that the luciferase activity of PART1 promoter-WT was strikingly increased by YY1 upregulation, whereas that of PART1 promoter-Mut showed no significant changes (Fig. 3E). In addition, RT-qPCR and western blot showed that the mRNA and protein levels of YY1 was highly expressed in DDP-resistant OC cells compared with that in OC cells (Fig. 3F, G). In sum, transcriptional factor YY1 induced the upregulation of PART1 expression in OC cells.
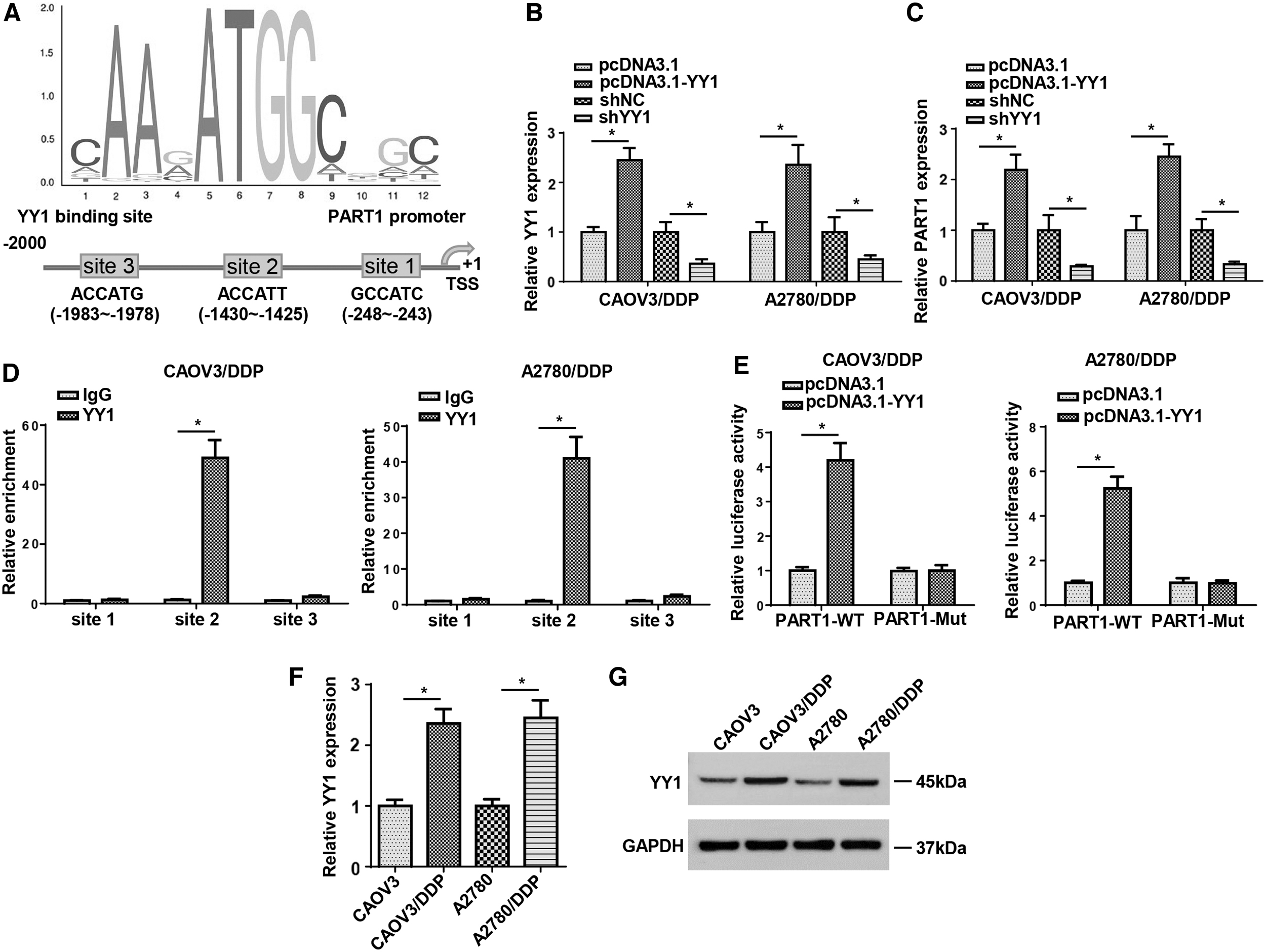
YY1 upregulated the expression of PART1 in OC cells.
PART1 acted as a molecule sponge for miR-512-3p
Accumulating studies indicated that lncRNAs exerted regulatory function through interacting with miRNAs. Therefore, starBase was performed to screen the potential targets of PART1. Among the predicted miRNA, miR-512-3p was reported as a tumor suppressor in various cancers (Zhu et al., 2015; Dou et al., 2020). To validate the relationship between PART1 and miR-512-3p, we conducted luciferase reporter assays and found that transfection of miR-512-3p mimics reduced the luciferase activity of the wild-type PART1 reporter gene, but not the mutant PART1 (Fig. 4A, B). In addition, RIP assays indicated that PART1 and miR-512-3p were highly enriched by Ago2, whereas IgG was not significantly enriched (Fig. 4C). Moreover, the level of miR-512-3p was lowly expressed in DDP-resistant OC cell lines (Fig. 4D). Finally, it was observed that the overexpression or knockdown of PART1 significantly decreased or increased miR-512-3p expression in DDP-resistant OC cells (Fig. 4E). These results demonstrated that PART1 negatively regulated miR-512-3p expression by direct interaction.
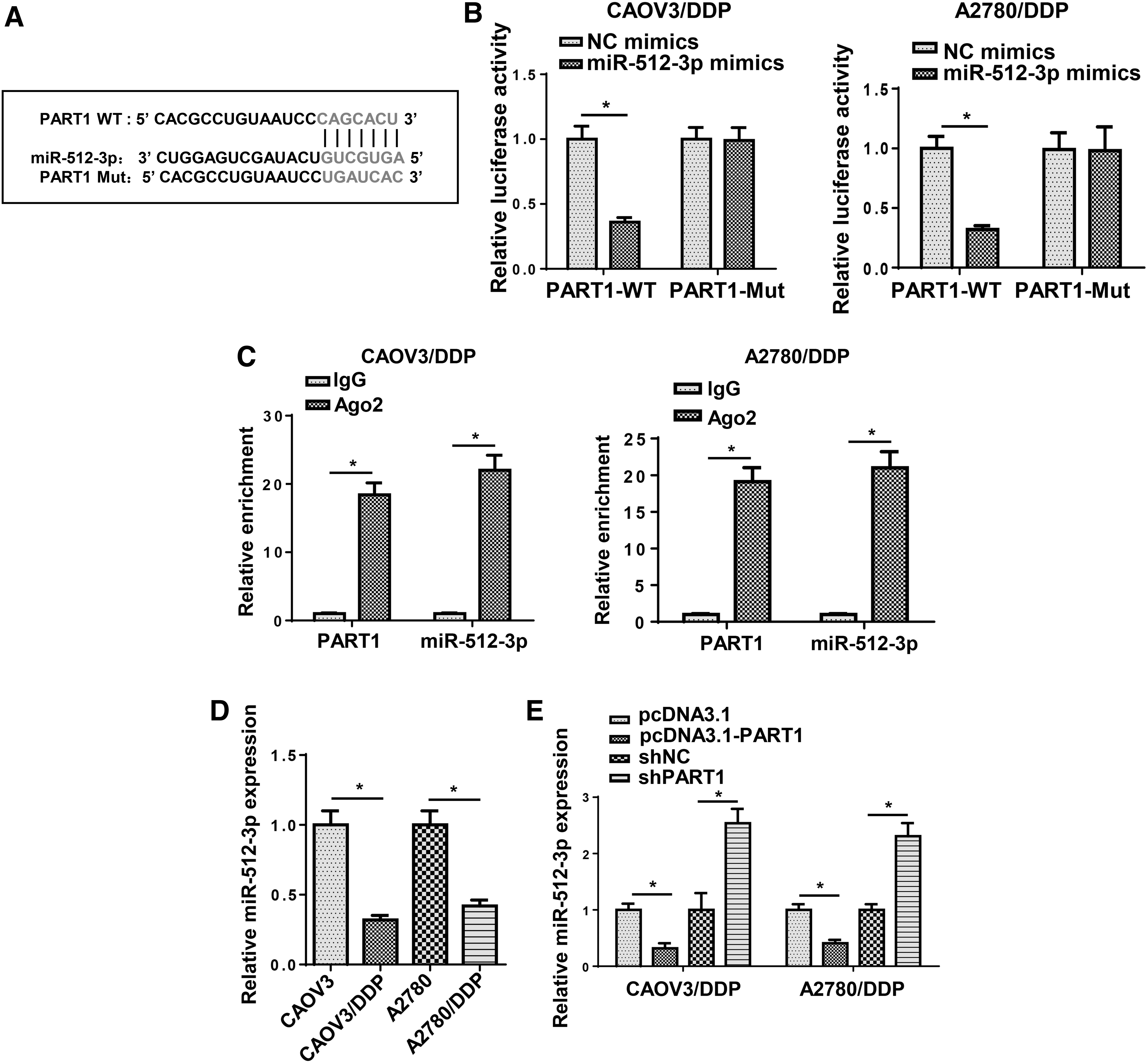
PART1 acted as a molecule sponge for miR-512-3p.
Knockdown of PART1 decreased DDP resistance of OC cells through miR-512-3p
To further explore whether PART1 regulated the DDP resistance of OC through miR-512-3p, functional assays were performed. First, CAOV3/DDP and A2780/DDP cells were transfected with NC mimics, miR-512-3p mimics, miR-512-3p mimics+ pcDNA3.1, and miR-512-3p mimics+pcDNA3.1-PART1. RT-qPCR showed that miR-512-3p expression was increased by miR-512-3p mimics and was further inhibited by PART1 (Fig. 5A). Then, CCK-8 and colony formation assays revealed that miR-512-3p overexpression distinctly decreased cell proliferation, which was abolished after pcDNA3.1-PART1 transfection (Fig. 5B, C). Besides, the addition of miR-512-3p restrained the migration and invasion and facilitated apoptosis of CAOV3/DDP and A2780/DDP cells, which was counteracted after pcDNA3.1-PART1 transfection (Fig. 5D–F). In sum, the earlier findings revealed that PART1 contributed to the DDP resistance of OC through regulating miR-512-3p.

Knockdown of PART1 decreased DDP resistance of OC cells through miR-512-3p.
CHRAC1 was directly targeted by miR-512-3p
Then, starBase showed that CHRAC1 was a potential target gene of miR-512-3p and the binding sequences between miR-512-3p and CHRAC1 are presented in Figure 6A. In addition, TCGA database showed that CHRAC1 was upregulated in OC tissues (Fig. 6B). To validate the binding ability between miR-512-3p and CHRAC1, luciferase reporter assays were performed and the results indicated that miR-512-3p mimics weakened the luciferase activity of CHRAC1-WT in DDP-resistant OC cells, whereas the mutant CHRAC1 exhibited no effect on luciferase activity (Fig. 6C). Furthermore, we found that CHRAC1 expression was higher in CAOV3/DDP and A2780/DDP cells than that in their parental cells (Fig. 6D). In addition, miR-512-3p mimics remarkably decreased the CHRAC1 expression, whereas miR-512-3p inhibitor displayed the opposite results (Fig. 6E). The aforementioned data indicated that miR-512-3p could directly target CHRAC1.
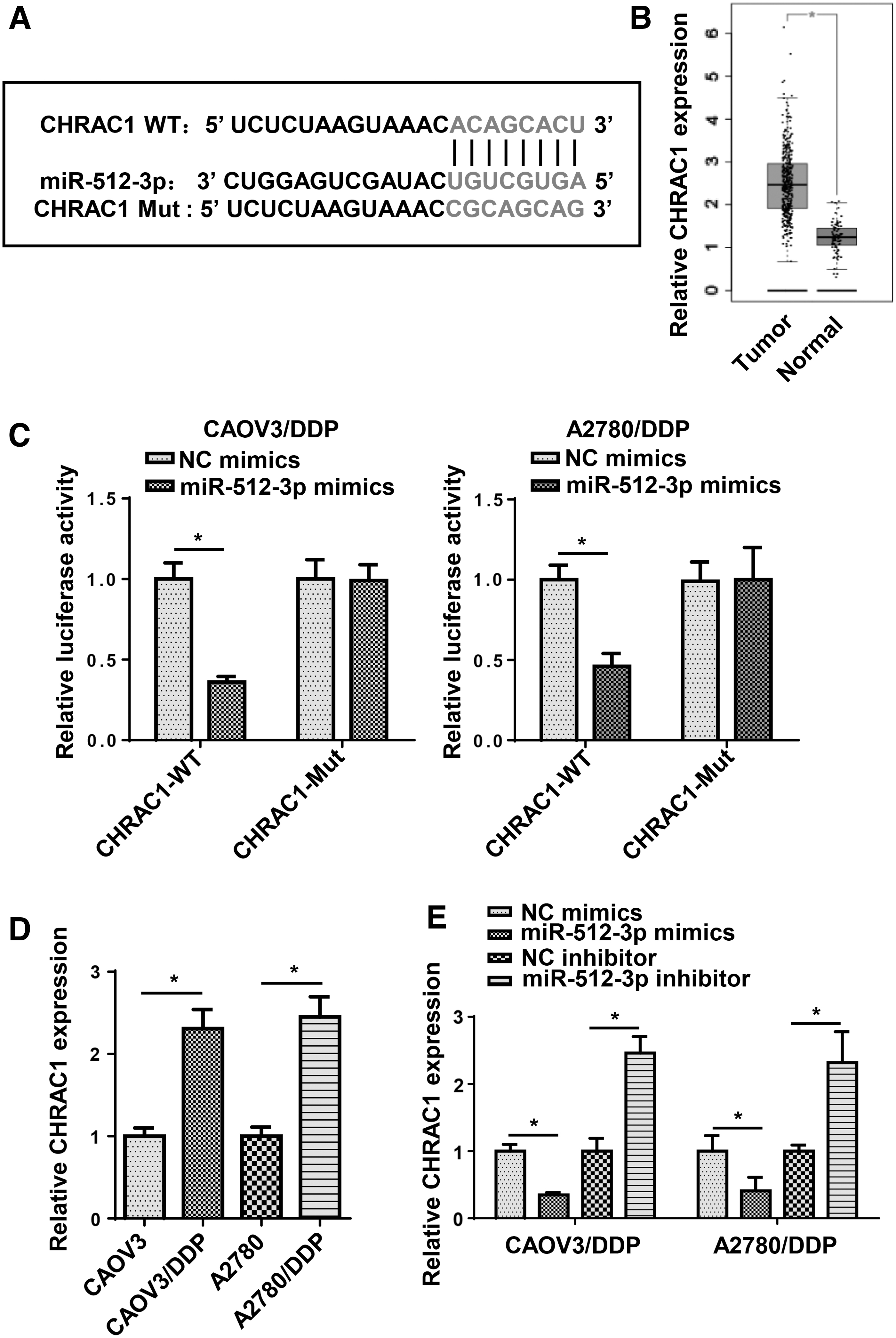
CHRAC1 was directly targeted by miR-512-3p.
PART1 regulated DDP resistance of OC through regulating miR-512-3p/CHRAC1 axis
To further investigate whether CHRAC1 was the downstream regulator of PART1/miR-512-3p axis in DDP-resistant OC, CAOV3/DDP cells were transfected with shNC, shCHRAC1, shCHRAC1+NC inhibitor, and shCHRAC1+ miR-512-3p inhibitor. RT-qPCR revealed that the inhibition of miR-512-3p abrogated the inhibitory effect of CHRAC1 knockdown on CHRAC1 expression (Fig. 7A). Moreover, functional analysis indicated that interference of CHRAC1 reduced cell proliferation, migration, and invasion, and facilitated apoptosis in CAOV3/DDP cells, whereas miR-512-3p inhibition neutralized these effects (Fig. 7B–F). Furthermore, the depletion of PART1 resulted in downregulation of CHRAC1, whereas the suppressive effects could be reversed by inhibition of miR-512-3p (Fig. 7G). All these data indicated that PART1 led to DDP resistance in OC cells through upregulating CHRAC1 expression by sponging miR-512-3p.
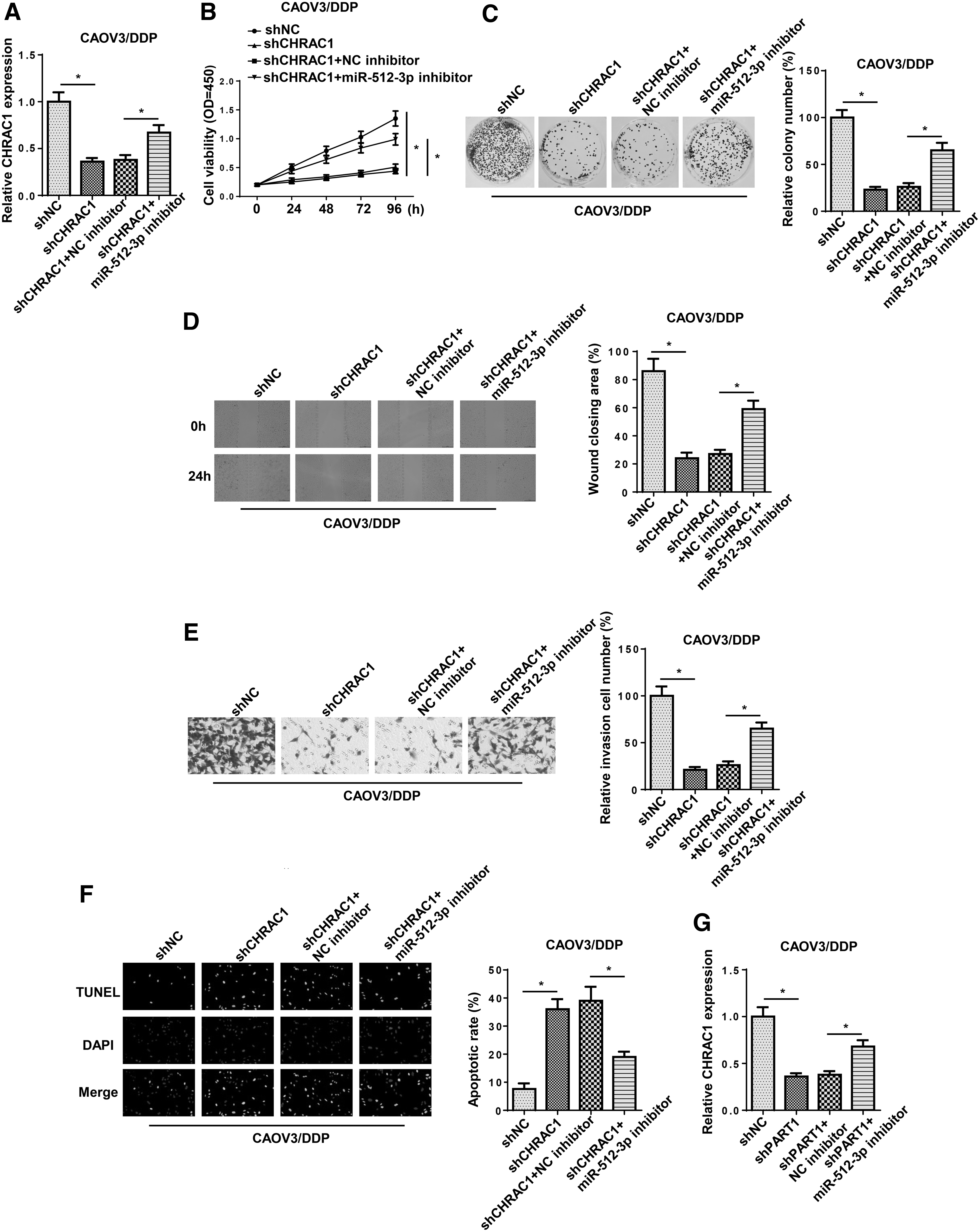
PART1 regulated DDP resistance of OC through regulating miR-512-3p/CHRAC1 axis.
Discussion
Increasing evidence indicates that lncRNAs are critical regulators s in chemoresistance of various tumors, including OC. For example, lncRNA MALAT1 knockdown significantly attenuated DDP resistance and enhanced DDP-induced apoptosis in DDP-resistant OC through inhibiting the Notch1 signaling pathway in vitro and in vivo (Bai et al., 2018). The depletion of lncRNA ANRIL expression decreased HMGA2 expression to facilitate the apoptosis and enhance DDP-sensitivity of OC cells through upregulating let-7a expression (Miao et al., 2019). KB-1471A8.2 significantly inhibited the development and paclitaxel resistance of OC cells by inhibiting CDK4 expression (Zhang et al., 2019). PART1 was confirmed to induce gefitinib resistance in esophageal squamous cell carcinoma and inhibit the efficacy of DDP in breast cancer through diverse regulatory pathways (Kang et al., 2018; Zhang et al., 2020). However, the biological significance of PART1 in DDP resistance of OC is still unknown. In our study, we first determined that PART1 was highly expressed in DDP-resistant OC and cell lines, and the depletion of PART1 distinctly reduced proliferation, migration, and invasion, and promoted the apoptosis of DDP-resistant OC cells, suggesting the inhibitory effect of PART1 in the chemosensitivity of OC cells to DDP.
Previous studies have indicated that transcription factors can cause the dysregulation of lncRNAs in human cancers (Liu et al., 2018; Pan et al., 2018; Zeng et al., 2018). Based on JASPAR and UCSC database, we found YY1 was a potential transcription factor of PART1. Transcriptional factor YY1 has been reported to enhance the expression of lncRNAs in human cancers. For instance, YY1 induced the increase of LINC00673 to facilitate breast cancer progression (Qiao et al., 2019). MCM3AP-AS1 was induced by YY1 and accelerated the progression of lung cancer through regulating miR-340-5p/KPNA4 axis (Li et al., 2020). YY1-mediated ARAP1-AS1 expedited the progression of colorectal cancer by regulating Wnt/β-Catenin pathway (Ye et al., 2019). However, the relationship between YY1 and PART1 in OC is still elusive. Based on mechanism research, YY1 induced PART1 transcription by binding to its promoter. Moreover, the expression of PART1 was positively regulated by YY1 in OC cells. These results revealed that YY1 was a transcriptional activator of PART1 in OC.
In another mechanism exploration, lncRNAs have been widely reported to act as competing endogenous RNAs (ceRNAs) to regulate the expression of target genes and participate in the tumorigenesis of various cancers (Qi et al., 2015; Chan and Tay, 2018). For example, MT1JP inhibited the progression of gastric cancer by acting as a ceRNA against miR-92a-3p and upregulating FBXW7 expression (Zhang et al., 2018). lncRNA UCA1 increased resistance to DDP by competitively binding miR-143 to regulate FOSL2 expression in OC (Li et al., 2019). In our study, miR-512-3p was predicted and validated as a downstream target of PART1. Previous reports have revealed the role of miR-512-3p in multiple malignant tumors. For example, Duan et al. (2020) reported that miR-512-3p was diminished in breast cancer cells, and the addition of miR-512-3p facilitated chemosensitivity, impeded growth, and metastasis of breast cancer cells. Chen et al. (2010) indicated that miR-512-3p accelerated Taxol-induced hepatocellular carcinoma cell apoptosis. In our research, we found that the addition of miR-512-3p promoted DDP sensitivity of OC, whereas this effect was neutralized after the transfection of PART1 overexpression plasmid. In sum, these results indicated that PART1 regulated the chemoresistance of OC by directly interacting with miR-512-3p.
To investigate the downstream mechanism of miR-512-3p, bioinformatics analysis was used to screen the candidate genes, and TCGA database indicated that CHRAC1, one of the potential downstream targets, was highly expressed in OC samples. CHRAC1 was confirmed to promote cell viability, and inhibiting apoptosis in breast cancer (Mahmood et al., 2014). In this study, it was observed that miR-512-3p could negatively regulate the expression of CHRAC1. Besides, the knockdown of CHRAC1 suppressed proliferation, migration, and invasion, and facilitated the apoptosis of DDP-resistant OC cells, which was neutralized after miR-512-3p inhibitor transfection. Furthermore, we demonstrated that PART1 regulated the expression of CHRAC1 through miR-512-3p, which identified a novel lncRNA-mediated ceRNA network in the chemoresistance of OC.
In summary, our study demonstrated that YY1-induced PART1 enhanced DDP resistance in OC through CHRAC1 axis by sponging miR-512-3p. These findings might provide a novel therapeutic strategy for overcoming DDP resistance in OC therapy.
Footnotes
Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
This study was supported by Zhenjiang Social Development Project (No. SH2019062) and Jiangsu Provincial Health Commission Top Talent Scientific Research Project (No. LGY2019027).