
Abstract
Baicalin-aluminum regulates the gut microbiome of piglets with diarrhea. However, whether it affects poultry gut microbiome composition and function remains unknown. In this study, we used metagenomic sequencing to explore the effects of baicalin-aluminum on gut microbiome changes in poultry when compared with animals administered colistin sulfate. Our data showed that important gut microbiome components consisted of Ruminococcaceae, Subdoligranulum, Bifidobacterium, Bifidobacterium pseudolongum, and Pseudoflavonifractor when broilers were administered baicalin-aluminum compared with colistin. At the species level, Lactobacillus salivarius, Bacteroides uniformis, Oscillibacter unclassified, Bacteroides fragilis, Ruminococcus torques, and Subdoligranulum unclassified abundance were significantly upregulated upon baicalin-aluminum treatment when compared with colistin administration. In addition, Gene Ontology (GO) enrichment analysis indicated that functional differentially expressed genes, which were in the top 30 GO enrichment terms, were associated with metabolic processes, catalytic activity, and cellular processes. Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis demonstrated that ABC transporters, oxidative phosphorylation, and phosphotransferase systems were the dominant signaling pathways in the baicalin-aluminum group when compared with the colistin group. Taken together, our data indicated that baicalin-aluminum modified broiler gut microbiome composition. These observations enhance our physiological insights of baicalin-aluminum-mediated functions in the broiler microbiome and potentially provide a novel therapy to manage both animal and human health.
Introduction
Currently, antibiotic resistance is a major threat to animal and human health, one of the contributors is high antibiotic levels used in global animal production (Kumar et al., 2020). Extensive antibiotic-use means antibiotics accumulated in the environment, and the generation and spread of resistance micro-organisms and their resistance genes to humans, animals, and environment (Yue et al., 2020). Once these antibiotics enter the body, they affect intestinal microbiota, which may lead to flora imbalance and gut-based diseases (Lopetuso et al., 2015). Thus, controlling antibiotic resistance is urgently required in the agricultural sector.
Colistin sulfate is widely used in the poultry industry to treat severe infections, and is considered the last-hope treatment against multidrug-resistant Gram-negative bacteria in the mid-1990s (El-Sayed Ahmed et al., 2020). With continuous colistin sulfate use, antibiotic resistance has now become critical for colistin sulfate resistance genes identified in pigs, poultry, and humans; for example, chromosome-encoded tigecycline resistance gene, tet(X14) in Empedobacter stercoris from pig feces (Cheng et al., 2020), and mcr in Klebsiella pneumoniae from patients and chickens (Sun et al., 2020). In addition, colistin sulfate appears to alter human intestinal microbiota and the antibiotic resistome in simulated human intestinal microbiota (Li et al., 2021). Similarly, colistin sulfate, in early mouse life, modified the gut microbiome and increased disease incidence in the spontaneous autoimmune insulin-dependent diabetes mouse model (Candon et al., 2015). Since implementation of a policy banning antibiotic addition to feed, the redevelopment of new antibiotic substitutes has gained considerable traction.
Baicalin was isolated from the Scutellaria baicalensis Georgi (Lamiaceae) root (Xu et al., 2020), and has demonstrated antibacterial (Wang et al., 2019), antivirus (Yang et al., 2020), antioxidation (Jia et al., 2021), and anti-inflammation characteristics (Fang et al., 2020). Previous research indicated that baicalin ameliorated neuropathology in a repeated cerebral ischemia-reperfusion injury mouse model, by remodeling the gut microbiota (Liu et al., 2020). Baicalin alleviated intestinal inflammatory and oxidative damage through inhibiting NF-κB and increasing mTOR signaling (Liao et al., 2020). Aluminum has been reported to have antimicrobial activity against Gram-negative bacteria (Gugala et al., 2019), thus baicalin-aluminum was synthesized by reaction of baicalin and alums from our laboratory and our previous data demonstrated that baicalin-aluminum could treat diarrhea of piglets and changed the gut microbiome composition of piglets with diarrhea (Fu et al., 2019). However, whether baicalin-aluminum modifies the broiler gut microbiome remains unclear.
In this study, we explored the effects of baicalin-aluminum on broiler gut microbiota composition and function, when compared with broilers supplemented with colistin sulfate. Also, microbiome Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways in broilers treated with or without baicalin-aluminum were investigated. Our results provide increased insights into the baicalin-aluminum effects on broiler gut microbiome composition, and the potential role of baicalin-aluminum in animal health.
Materials and Methods
Drugs and experimental design
This study was conducted in strict accordance with China Regulations for the Administration of Affairs Concerning Experimental Animals 1988 and the Hubei Regulations for the Administration of Affairs Concerning Experimental Animals 2005. The animal experiments and animal care were approved by Animal Care and Use Committee of Wuhan Polytechnic University, Hubei Province, China (EM949, November 4, 2020). Fifteen 14-day-old white feather broilers, weighing 0.3–0.4 kg, were obtained from Yichang MUKANG animal husbandry Co., Ltd. (Yichang, China) for study.
The synthesis method for baicalin-aluminum, and extract use were previously reported (Fu et al., 2019). Colistin sulfate was purchased from Livzon Group Fuzhou Fuxing Pharmaceutical Co., Ltd. (Fuzhou, China). The broilers were reared in a poultry facility with standardized conditions on a 20:4 h light:dark cycle under 19–28°C. Fifteen female broilers were randomly divided into three groups: a normal control group, a colistin sulfate group, and a baicalin-aluminum group. Broilers in the normal control group were fed with a basic diet. In the colistin sulfate group, broilers were fed a basic diet supplemented with 20 g colistin sulfate/ton, and the baicalin-aluminum group was given 0.5% baicalin-aluminum extract in the basic diet. After 14 days feeding, the fresh stool samples were collected and immediately snap-frozen in liquid nitrogen, and stored at −80°C for use. Stool samples from each group (each group five samples) were obtained for metagenomic sequencing.
Extraction of DNA from stool and DNA sequencing
DNA extraction and purification from stool was performed according to a previous report (Feye et al., 2020). In brief, stool was stored in liquid nitrogen. Genomic DNA was isolated using the Qiagen Qiamp Fast DNA Stool Mini Kit (Qiagen, Germany) according to manufacturer's instructions. DNA purity and quality were assessed using a Nanodrop instrument (Thermo Scientific). DNA library construction was conducted according to manufacturer's protocols (Illumina, San Diego, CA) (Yan et al., 2020), and library quality was verified using a Qubit dsDNA HS assay kit (Invitrogen, Waltham, MA). We conducted 2 × 150 base pair (bp) paired-end metagenomic sequencing on the Illumina NovaSeq platform.
Metagenomic analysis
Raw reads were filtered to discard sequences with a quality value <20, flanking sequences, and sequencing reads with lengths <45 bp, to generate quality sequences. These sequences were mapped to host genomes to derive clean reads using Bowtie 2 (version 2.3.1) (Meera et al., 2020). Gene predictions were performed using MetaGeneMark (
Statistical analysis
All statistical analysis was determined by using R software. All comparisons were carried out pairwise for every group. The Wilcoxon rank sum test and correlation analyses were used to evaluate all data and p-values at ≤0.05 were deemed statistically significant. PCoA was determined with weight-UniFrac distance matrix by QIIME. All significance determination was at p ≤ 0.05.
Results
The effects of baicalin-aluminum on the broiler gut microbiome
Using metagenomic sequencing analysis, structural changes in the broiler gut microbiome were explored when animals were fed colistin and baicalin-aluminum. We identified 1096928331 clean reads from 15 stool samples, with 73128555 ± 8810935 sequences per sample (Table 1).
Statistical Summary Analysis of the Sequencing Data
PCA was used to investigate relationships between the different samples; most samples from the baicalin-aluminum, colistin, and control groups were present in a mix distribution, which were not gathered into one single cluster (Fig. 1). PCoA was used to explore the microbiome composition of different groups based on evolutionary distance. PCoA demonstrated that control, colistin, and baicalin-aluminum groups exhibited significantly different changes (Fig. 2), and the microbiome composition of the colistin and baicalin-aluminum groups displayed distinct clusters (Fig. 2). In addition, the overall broiler gut microbiome was modified by colistin and baicalin-aluminum supplementation (Fig. 2).
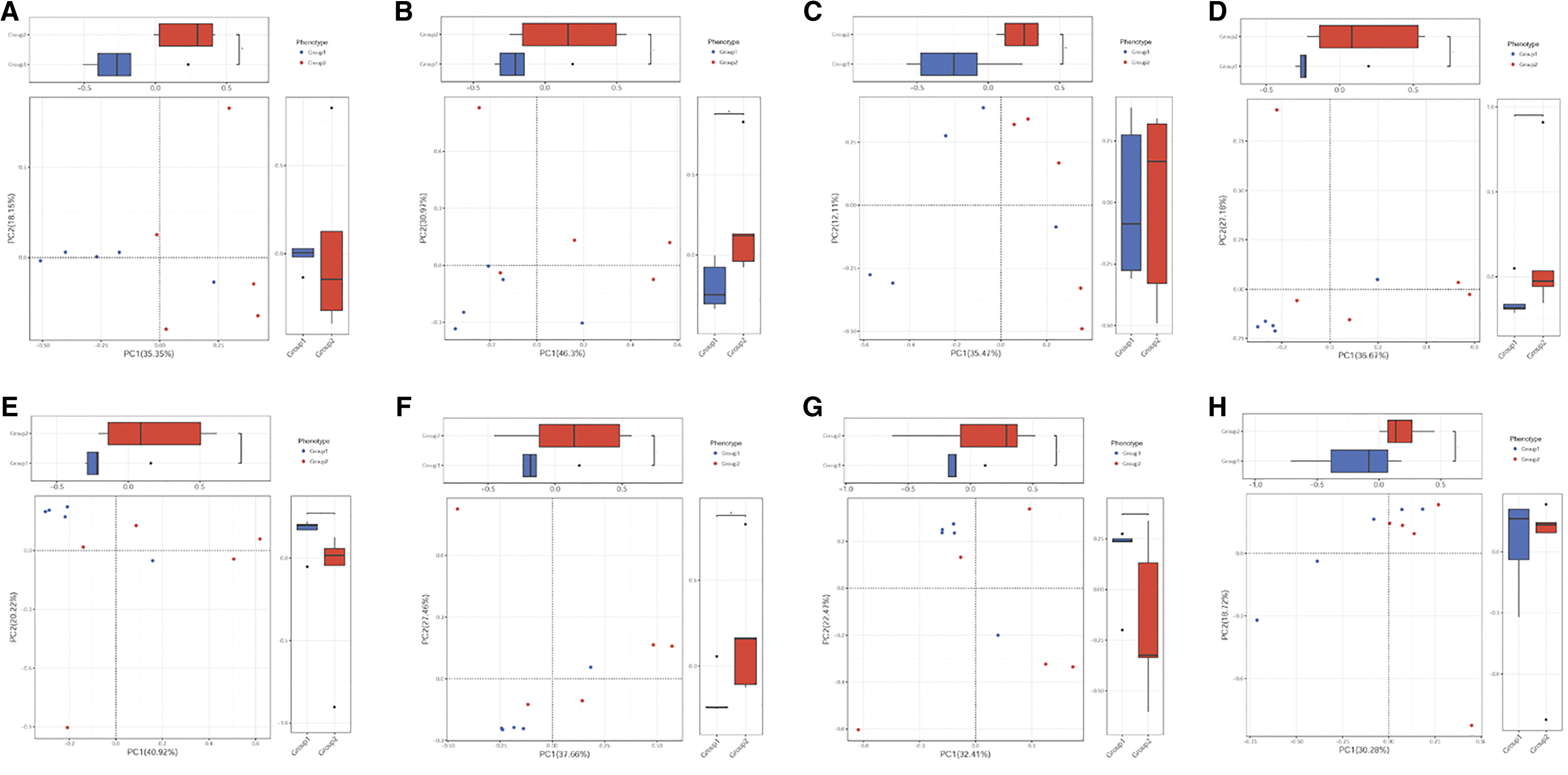
PCA on the effects of baicalin-aluminum on broiler gut microbiome composition. PCA of gut microbiome changes of ARDB

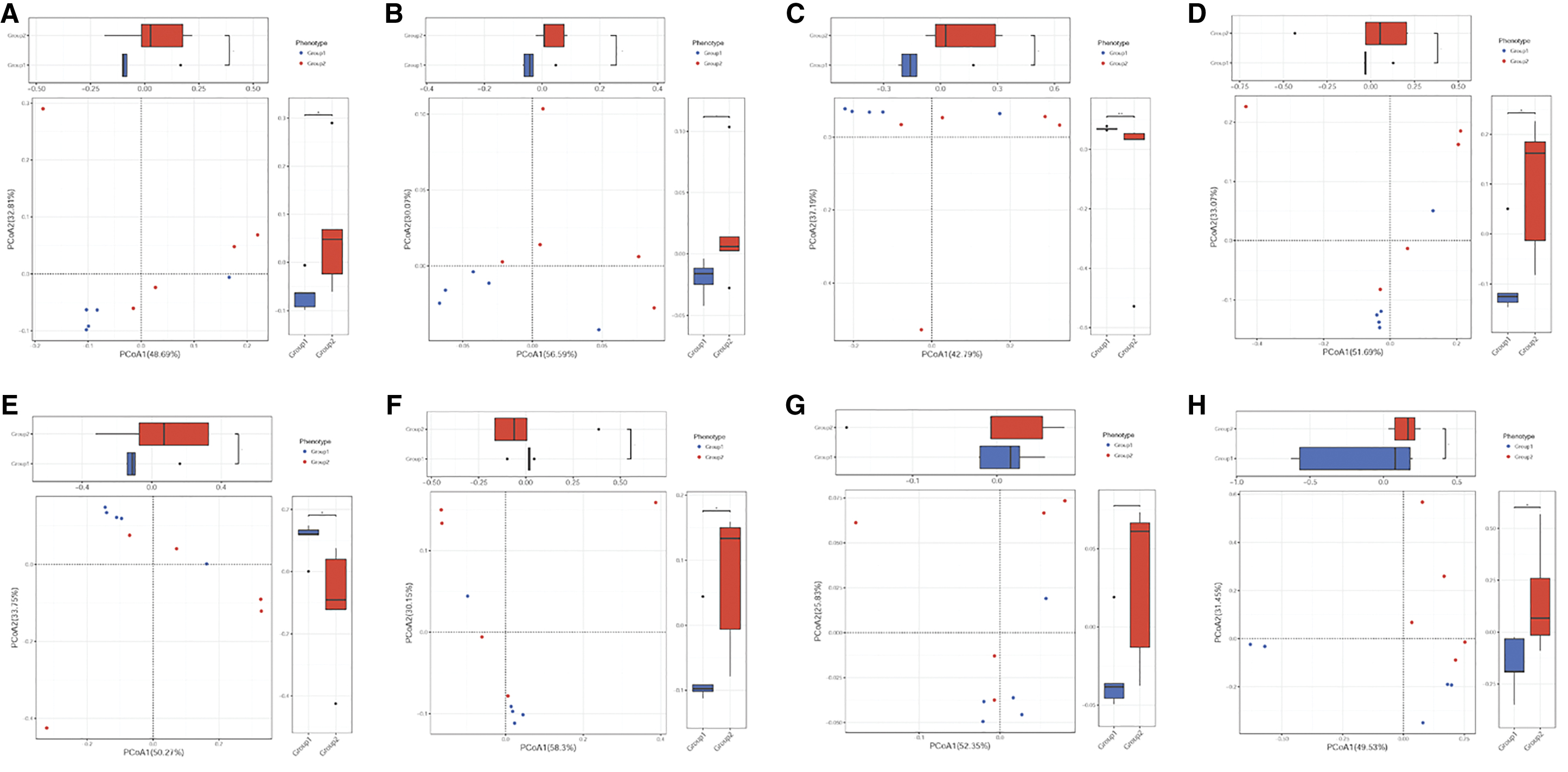
PCoA on the effects of baicalin-aluminum on broiler gut microbiome composition. PCoA of gut microbiome changes of ARDB

The effects of colistin and baicalin-aluminum on gut microbiome phylotypes
From the species community structure diagram of broilers feeding with colistin, Lactobacillus johnsonii, Weissella paramesenteroides, Lactococcus lactis, and Streptococcaceae were the main microbiome components when compared with the control group (Fig. 3A, B and Supplementary Tables S1 and S2). However, when broilers were administered baicalin-aluminum, the important gut microbiome components were Ruminococcaceae, Subdoligranulum, Bifidobacterium, B. pseudolongum, and Pseudoflavonifractor when compared with the colistin group (Fig. 3C, D, Supplementary Tables S3 and S4). These data indicated that baicalin-aluminum regulated gut microbiome composition (Fig. 3).

Species community structure diagram of broiler gut microbiome using linear discriminant analysis effect size (LEfSe).
Furthermore, broiler taxonomy analysis at genus, phylum, and species levels were also determined (Fig. 4). Genus level analyses demonstrated that Flavonifractor and Ruminococcaceae noname levels were decreased in the colistin group when compared with the control group (Fig. 4A and Supplementary Table S5). However, Pseudoflavonifractor and Subdoligranulum were increased in the baicalin-aluminum group when compared with the colistin group (Fig. 4D and Supplementary Table S6). Species level analyses indicated that Pseudoflavonifractor capillosus, S, unclassified, and Ruminococcaceae bacterium D16 abundance were reduced when the broilers were fed colistin (Fig. 4C and Supplementary Table S7), whereas S. unclassified, P. capillosus, and B. pseudolongum were significantly increased and L. johnsonii, W. paramesenteroides were significantly decreased in broilers administered baicalin-aluminum when compared with colistin (Fig. 4F and Supplementary Table S8).
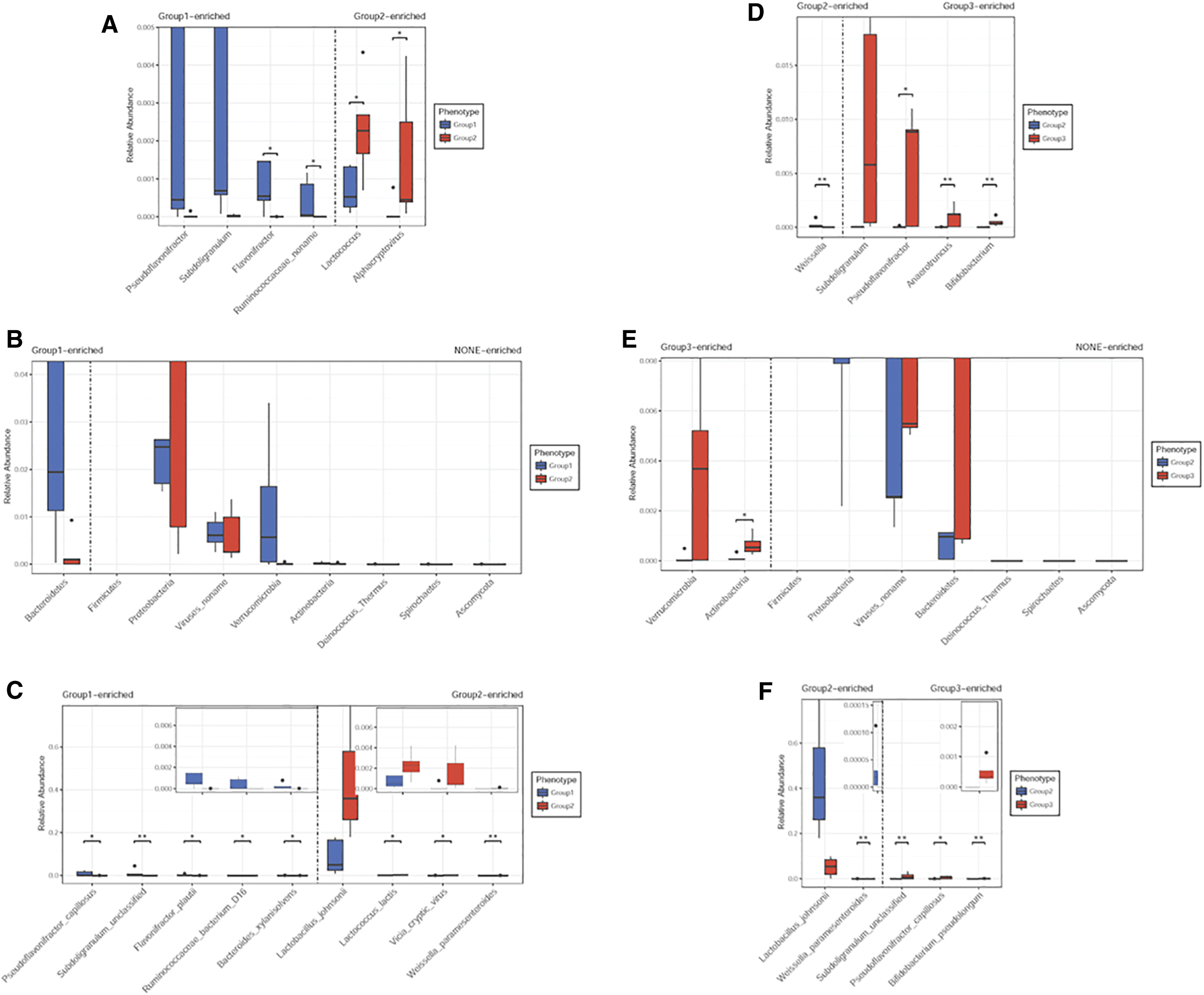
Detection of genus
We also generated a taxonomy comparison heatmap based on genus, phylum, and species levels. These data showed that Escherichia and P. noname were significantly increased at the genus level in the colistin group when compared with the control group (Fig. 5A and Supplementary Table S9), whereas in broilers fed baicalin-aluminum, these bacteria were significantly decreased, and Bacteroides, Oscillibacter, and Subdoligranulum were increased when compared with the colistin group (Fig. 5D and Supplementary Table S10). At the species level, L. salivarius, O. unclassified, and B. fragilis were reduced, and L. johnsonii, Escherichia coli, and P. noname unclassified were increased upon colistin treatment when compared with the control group (Fig. 5C and Supplementary Table S11). When broilers were fed baicalin-aluminum, L. salivarius, B. uniformis, O. unclassified, B. fragilis, R. torques, and S. unclassified levels were significantly increased when compared with the colistin group (Fig. 5F and Supplementary Table S12).

Heatmap of genus
Baicalin-aluminum modifies gene expression in the broiler microbiome
To explore gut microbe changes to colistin and baicalin-aluminum supplementation, GO and KEGG enrichment analysis were conducted. The data indicated that in broilers fed colistin, the metabolic process, binding, and single-organism process, which the differentially expressed genes were enriched in, were dominant when compared with the control group (Fig. 6A and Supplementary Table S13), whereas the metabolic process, catalytic activity, and cellular process were the main when the broilers were treated with baicalin-aluminum compared with the colistin group (Fig. 6B and Supplementary Table S14). KEGG analyses demonstrated that ABC transporters, carbon metabolism, glycolysis/gluconeogenesis, and the citrate cycle (TCA cycle) were the main signaling pathways when broilers were administered colistin when compared with the control group (Fig. 6C and Supplementary Table S15). However, when broilers were fed baicalin-aluminum, the dominant signaling pathways were ABC transporters, oxidative phosphorylation, the phosphotransferase system, and ribosome when compared with the colistin group (Fig. 6D and Supplementary Table S16).

DAVID analysis of GO and the top 30 signaling pathways enrichment terms.
Discussion
We observed that baicalin-aluminum significantly altered the broiler gut microbiome when compared with colistin and control groups. Previous research demonstrated that baicalin-copper modulated the gut microbiota, inflammatory responses, and hormone secretion in deoxynivalenol challenged piglets (Zha et al., 2020b). Baicalin-copper also could alleviate the growth impairment triggered by deoxynivalenol and enhanced antioxidant capacity and nutrition absorption of piglets (Zha et al., 2020c). Baicalin zinc inhibited oxidative stress and enhanced nutrition absorption in deoxynivalenol challenged pigs (Zha et al., 2020a). Our previous data showed that baicalin-aluminum regulated gut microbiome structure in piglets with diarrhea (Fu et al., 2019). However, whether baicalin-aluminum affected the broiler gut microbiome was unexplored. To address this, we observed that baicalin-aluminum modified the broiler gut microbiome and this change would prevent or control infectious diseases or metabolic disorders, which may serve as a new therapeutic to control poultry disease.
In this study, five broilers were used in each group for metagenomic sequencing because of restriction of sequencing price. In the previous study, 16 s rRNA gene amplification method was used to determine the gut microbiomes change (Fidler et al., 2020). In this study, metagenomic sequencing method was utilized to comprehensively reflect the gut microbiomes composition. After baicalin-aluminum supplementation, Subdoligranulum levels were significantly increased when compared with the colistin group. Previous research reported that Subdoligranulum, as a beneficial bacterial genera, produced short-chain fatty acids essential for human health (Zhang et al., 2015). It was previously shown that vegan diets could prevent hypertension and human metabolic disease, which may improve Subdoligranulum abundance (Jia et al., 2020). It was also reported that Subdoligranulum was an optimal probiotic strain for spore-forming butyrate producer functions in the cecal microbiota of 7-day-old chickens (Polansky et al., 2015), and the core microbiota for global commercial swine production (Holman et al., 2017). Subdoligranulum also reduced nonstarch polysaccharides in the broiler cecum, thereby eliminating intestinal pathogens and improving poultry growth performance (Xia et al., 2019). Thus, baicalin-aluminum appeared to regulate gut microflora composition, thereby improving gut function. And our results provide the foundation to modify the poultry gut microbiota in the next research for potential health benefits.
The gut microbiome is a key factor in maintaining intestinal barriers and promoting immunological tolerance. L. salivarius is a core member of the animal gut microbiome and is a promising probiotic (Lee et al., 2017). L. salivarius enhanced intestinal health and alleviated inflammation responses in IPEC-J2 cells induced by enterotoxigenic E. coli (ETEC) K88, through the NF-κB signaling pathway (Qiao et al., 2020). L. salivarius BP121 also prevented cisplatin-induced acute kidney injury by inhibiting uremic toxins (Lee et al., 2020). Similarly, L. salivarius LA307 attenuated skin inflammation in mice by modulating other gut microbiota (Holowacz et al., 2018). In addition, L. salivarius inhibited Streptococcus mutant growth, virulence, biofilm formation, and gene expression (Wasfi et al., 2018). Thus, based on this evidence, L. salivarius could be considered a probiotic for animal and human health (Chaves et al., 2017). In our study, L. salivarius abundance was significantly increased when broilers were treated with baicalin-aluminum, thus baicalin-aluminum effects may be mediated by L. salivarius, potentially relieving inflammatory responses and enhancing antimicrobial properties. However, these mechanisms require further investigation.
B. uniformis is a putative bacterial species, and was associated with isoflavone genistein degradation in human feces (Renouf and Hendrich, 2011). Evidence has suggested that B. uniformis ameliorated metabolic and immunological dysfunctions in high-fat-diet-induced obese mice (Gauffin et al., 2012). Also, it protected against metabolic disorders and obesity in Wistar rats (Gómez Del Pulgar et al., 2020). The administration of B. uniformis delayed nonalcoholic fatty liver disease progression through adiponectin (Zhou et al., 2018). Our results suggested that baicalin-aluminum increased beneficial gut bacteria such as B. uniformis, which may rebalance gut microbiome composition.
In this study, we found that ABC transporters was the main signaling pathway in the baicalin-aluminum group compared with the colistin group by KEGG analysis. The genes of vcaM, nikA, urtC, msrA, urtA, and lapB were involved in the ABC transporters and their expression levels were downregulated. Previous research showed that VCAM was associated with inflammatory responses infected by C. muridarum (Nagarajan et al., 2020). NikA is critical for metal acquisition of Brucella survival during infection processes (Budnick et al., 2018). msrA was thought to be related to virulence and resistance of Staphylococcus aureus (Roberts et al., 2020). Thus, we speculate that baicalin-aluminum could inhibit some important genes expression resulting in antimicrobial function, but the specific mechanism needs further study.
Taken together, our data demonstrated that baicalin-aluminum regulated broiler gut microbiome composition. We inferred that our research found the new characteristic of baicalin-aluminum on enhancing the abundances of the probiotic bacteria thereby realizing its anti-inflammation responses and antimicrobial functions, which might provide a novel therapy to maintain animal and human health. In the future, we will evaluate the function of baicalin-aluminum in the poultry chickens and the safety to animals and the environment.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the National Key R&D Plan (grant no. 2018YFD0500301).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Supplementary Table S7
Supplementary Table S8
Supplementary Table S9
Supplementary Table S10
Supplementary Table S11
Supplementary Table S12
Supplementary Table S13
Supplementary Table S14
Supplementary Table S15
Supplementary Table S16
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.