
Abstract
Previous studies have shown that lysine-specific demethylase 1 (LSD1) could regulate cell cycle progression through demethylation. The 3′domain of HOX transcript antisense RNA (Hotair) combined with the LSD1/CoREST/REST complex helps LSD1 target the corresponding gene. However, its role in mice's myocardial regeneration is still unclear. The heart from neonatal mice shows strong myocardial regeneration ability, but this ability disappears 7 days after birth. Our study shows that the myocardial tissue highly expresses Hotair and Lsd1 within 1 week after birth, consistent with the myocardial regeneration time window. Knockdown Lsd1 or Hotair expression by RNA interference could inhibit myocardial regeneration and cardiomyocyte proliferation. Our results suggest that Hotair-mediated demethylation of LSD1 may play an important role in myocardial regeneration in neonatal mice.
Introduction
Despite the continuous extensive improvement in clinical treatment, cardiovascular disease is still an increasingly serious health problem (Lu and Thum, 2019; Timmis et al., 2020). According to World Health Organization (WHO)'s estimates, the increased cardiovascular risk may double the victim number by the year 2050, including obesity and diabetes.
Lost cardiomyocytes can only be superseded by scar tissue, will affect the contractility of the remaining myocardium, cause compensatory hypertrophy at the very beginning, then dilatation of the myocardial wall, ineluctably leading to heart failure (Garbern et al., 2020; Pianca et al., 2020). Except for some lower vertebrates, adult mammals do not have this regenerative capacity. Intense myocardial regeneration can occur in newborn mice, but this ability will be lost at 7 days after birth (Porrello et al., 2011). The cardiomyocyte turnover rate is nearly 1% every year but decreased to 0.45% over the age of 75 (Bergmann et al., 2009).
Long noncoding RNAs (LncRNAs), longer than 200 nucleotides in length, regulate gene expression through transcription and posttranscriptional levels (Li et al., 2017; Mas and Huarte, 2020). Recently, several studies highlight the important role of LncRNA-mediated regulation in endogenous cardiac regeneration (Chen et al.,, 2017; Li et al., 2018a).
The lncRNA HOX transcript antisense RNA (Hotair), which acts as a molecular scaffold, decreases histone 3 lysine 4 dimethylation (H3K4me2) by recruiting and binding with lysine-specific demethylase 1 (LSD1)/CoRest/REST complex (Chu et al., 2014). Hotair has been recognized as an adverse prognostic factor in different cancer types (You et al., 2014). It is also considered to have a heart-protective effect. Hotair could protect myocardial cells from hypoxia-induced apoptosis by reducing the ratio of Bax to Bcl-2 (Gao et al., 2017; Zhou et al., 2019). However, little is known about the role of Hotair in mammalian cardiomyocyte proliferation and myocardial regeneration.
The purpose of this study was to discover related regulators of myocardial cell proliferation and myocardial regeneration, thereby improving the early prognosis of heart failure. Our study found that the high expression levels of Hotair and LSD1 in mouse myocardial tissues are consistent with the mouse myocardial regeneration time window. Downregulation of Hotair or Lsd1 can inhibit myocardial regeneration and myocardial cell proliferation, and affect H3K4me2 expression levels. Therefore, we speculate that Hotair may regulate cardiomyocyte proliferation and myocardial regeneration by regulating the demethylation of LSD1.
Materials and Methods
Experimental animal and ethics statement
Six male and 6 female Institute of Cancer Research (ICR) mice (8 weeks) were purchased from SPF (Beijing) Biotechnology Co, Ltd. All mice were housed and bred in the animal facility at the Institute of Radiation Medicine, Peking Union Medical College, and animal experiments were performed following the Institutional Animal Care and Use Committee guidelines (approval no. IRM-DWLL-2019113). There were six animals/group in each experiment.
Cardiomyocyte isolation
Cardiomyocyte isolation and culture were followed by reported protocol (Li et al., 2020). Briefly, ICR mice were anesthetized with 5% isoflurane and sacrificed by decapitation. Small pieces of ventricular myocardium (1–2 mm3) were dissociated with trypsin (0.2%) and collagenase (0.1%). The cell suspension was preplated in the cell culture dish at 37°C for 120 min. The cardiomyocytes were obtained from the cell suspension after being centrifuged with 200 × g for 8 min and culture with complete DMEM/F12 culture medium.
Real-time quantitative PCR and Western blotting
Ventricular muscle tissues were obtained from mice at 1, 4, 7, 14, and 21 days after birth (atrial tissue and excess blood were removed). Tissue RNA was extracted by RNAiso (TaKaRa, China), and then subjected to real-time quantitative PCR (qPCR) after reverse transcription into cDNA. qPCR was conducted on Roche Light Cycler 480 (Switzerland). Relative mRNA was determined with the comparative CT (2−▵▵ct) method. Primer sequences are listed in the Supplementary Table S1.
Extraction of tissue protein was with radio-immunoprecipitation assay lysis plus protease inhibitor cocktail (MedChemExpress) and determination of protein concentration by the Bicinchoninic Acid Protein Assay method. The protein was isolated on 10% polyacrylamide gel electrophoresis, the protein was transferred to polyvinylidene fluoride membrane, and blocked by 5% skim milk powder. Membranes were incubated overnight at 4°C with mouse anti-GAPDH antibody (ab8245, 1: 2,000; Abcam, UK); rabbit anti-KDM1/LSD1 antibody (ab129195, 1: 20,000; Abcam); rabbit anti-histone H3 (dimethyl K4) antibody (ab7766, 1: 1,000; Abcam); and rabbit anti-histone H3 (dimethyl K9) antibody (ab32521, 1: 1,000; Abcam). After, incubation with goat anti-rabbit secondary antibody (ab6721, 1:10,000; Abcam) and goat anti-mouse secondary antibody (ab205719, 1:10,000; Abcam) for 1 h at room temperature. Proteins were visualized according to the BeyoECL Moon protocol (P0018FS; Beyotime, China).
Apical resection
Apical resection (Apr) was performed on mice in stage P0 (within 24 h of birth) following the previously reported protocols (Porrello et al., 2011). Briefly, left parasternal thoracotomy was performed at the fourth intercostal space and the cardiac apex was resected (15%). At 21 days, cardiac function was measured with a Vevo 2100 Imaging System (Visual Sonics, Toronto, Canada).
Transfection
shRNA guide sequence cloned into pLKO.1 backbone (Addgene; Plasmid no. 10878) according to the protocol from Addgene. Small hairpin sequences are listed in Supplementary Table S2. Briefly, shRNA expression vectors were cotransfected with the psPAX2 (Addgene; Plasmid no. 12260) and pMD2.G (Addgene; Plasmid no. 12259) with Lipofectamine™ 2000 transfection reagent (Thermo Fisher Scientific, Waltham, MA). Lentiviruses were concentrated with ultracentrifugation for 120 min at 19,500 rpm using a Beckman SW28 ultracentrifuge rotor at 4°C.
On the second day after mouse Apr, lentivirus (10 μL, 1 × 108 IU/mL) was injected to the fifth intercostal space of the left margin of the sternum delivering short hairpin RNAs targeted to Hotair, LSD1, or scramble.
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide cell proliferation assay
Cell proliferation was measured by a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTS) assay (Promega) according to the provided procedures and absorbance at 490 nm was collected with Cytation 5 (BioTek).
Cell cycle and apoptosis analysis
After washing with precooling phosphate-buffered saline, the cardiomyocytes were fixed with 70% ethanol at 4°C overnight and then stained with Propidium Iodide (PI) staining solution plus with RNase (Cell Signaling Technology). The cell cycle was analyzed with a FACSCalibur instrument (San Diego, CA). A Sigma-Aldrich Annexin V-FITC Detection Kit was applied for the cell apoptosis according to the instruction manual and analyzed using the same Flow cytometer mentioned above.
Immunofluorescence detection
On the second day of Apr, mice were injected intraperitoneally with 5-bromo-2′-deoxyuridine (BrdU) labeling at a dose of 50 mg/Kg, and cardiac tissue was taken 5 days after continuous injection. Tissue sections were incubated overnight at 4°C with mouse anti-BrdU antibody (RT 1081, 1:200; huabio, China), rabbit anti-Ki67 antibody (ER 1802-31, 1:100; huabio), and rabbit anti-PH3 antibody (ab5176, 1: 1000; Abcam), followed by incubation of the corresponding secondary antibodies at room temperature for 2 h. Image acquisition was performed with fluorescence microscope after sealing with Antifade Mounting Medium with DAPI (P0131; Beyotime, China).
Statistical methods
SPSS 22.0 software was used to analyze the data, and the results were expressed as mean ± standard deviation (
Results
Time-dependent expression of Hotair and LSD1 in the heart tissue of mice after birth
Interestingly, our results showed that the mRNA level of Hotair and Lsd1 significantly decreased in the myocardial tissue at P14 (14-day-old) (Fig. 1A), while the protein expression was also impeded and H3K4me2 level increased after 14 days of birth (Fig. 1B). On the second day after Apr, lentivirus was injected into the fifth intercostal space of the left margin. The mouse was sacrificed at day 21 and ventricular muscle tissues were obtained for further study.
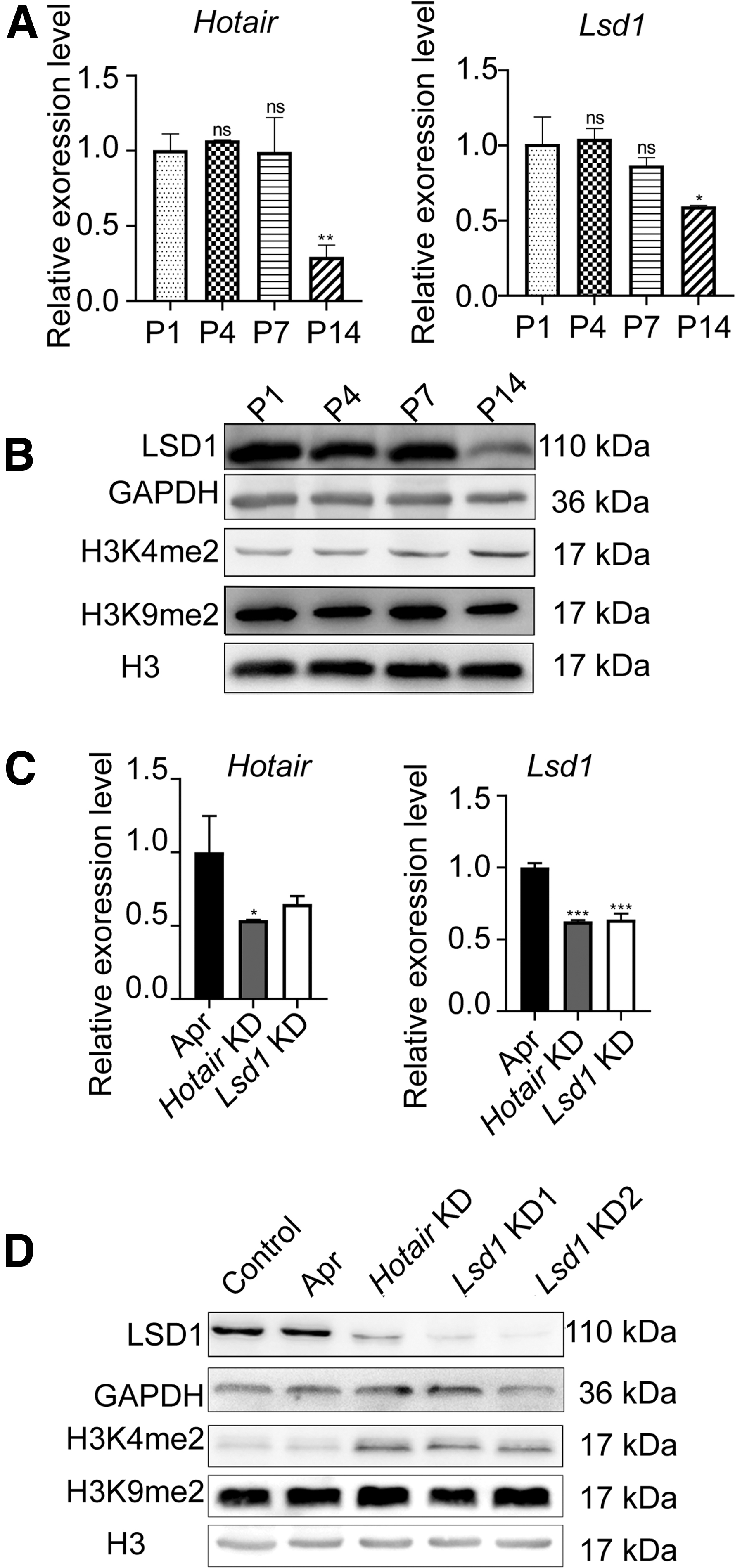
Expression of Hotair and LSD1 in the myocardial tissue of mice.
We did successfully knockdown (KD) Hotair expression in the Hotair KD group by RNAi (Fig. 1C). The expression of Lsd1 in the myocardial tissue of the Hotair KD and Lsd1 KD group was significantly lower than the control group (p < 0.001) (Fig. 1C). In the Hotair KD group, LSD1 protein expression was downregulated and the H3K4me2 level became higher than Apr and control group in myocardial tissues (Fig. 1D). We selected the RNAi sequence of Lsd1 KD2 in further study.
Knocking down Hotair or LSD1 inhibits neonatal myocardial regeneration
We did further KD Lsd1 or Hotair expression with RNAi. We first confirmed the regenerative ability of the neonatal heart within 21 days. Figure 2A shows the schematic diagram of this study. We also show a photo of Apr in Figure 2B. The left ventricular ejection fraction (EF%) and left ventricular short-axis contraction rate (FS%) values in the Apr group did not show a significant difference with Control and sham group (Fig. 2C, D). EF% and FS% of the Hotair KD group and Lsd1 KD group were significantly different from the Apr group (p > 0.05) (Fig. 2C, E). Figure 2E showed that the systolic blood pressure (SBP) and diastolic blood pressure (DBP) value of the LSD1 KD group was significantly lower (p < 0.01, p < 0.05) than the Apr group.
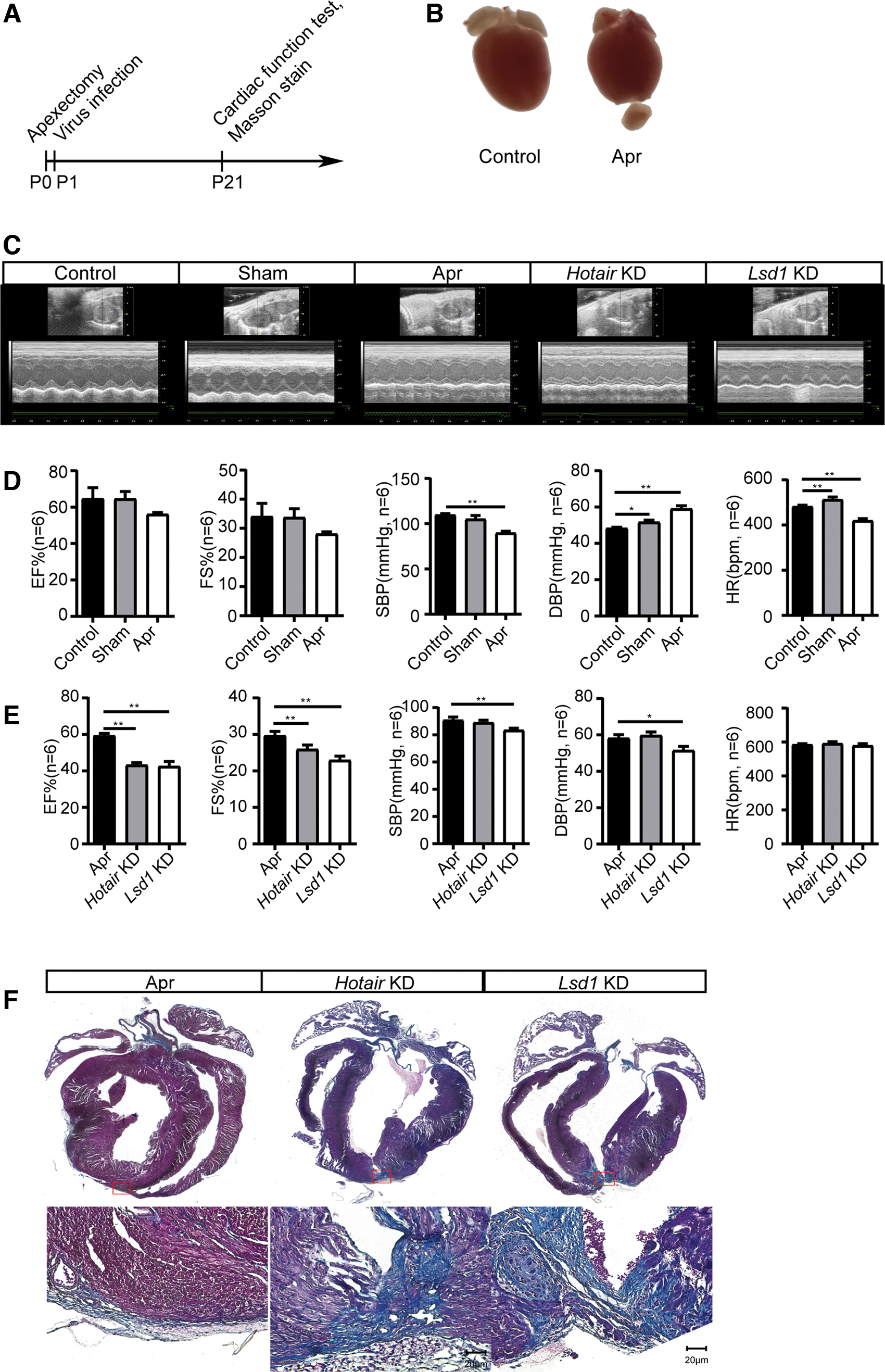
Knocking down Hotair or Lsd1 inhibits neonatal myocardial regeneration.
Masson staining showed that the apical scar tissue area was formed in the Hotair KD and Lsd1 KD group, but fewer in the Apr group (Fig. 2F).
Inhibition of Hotair or LSD1 expression can inhibit cardiomyocyte proliferation in vivo
To further illustrate the mechanism, we performed immunofluorescence staining with PH3, Ki67, and BrdU. Figure 3A shows the schematic diagram of this study. Compared with the control group, the number of pH3-, Ki67-, and BrdU-positive cells in the Apr group increased significantly; whereas the number of pH3- and BrdU-positive cells in the Hotair KD and Lsd1 KD group was lower than in the Apr group; (Fig. 3B). Interestingly, we could find KI67-positive cells in the Lsd1 KD group fewer than in the Apr group; but in the Hotair KD group and control group, we almost could not find KI67-positive cells (Fig. 3B).

Inhibition of Hotair or LSD1 expression can inhibit cardiomyocyte proliferation in vivo.
KD of the expression of Hotair or LSD1 could cause S phase arrest
Next, we isolated the cardiomyocytes from the neonatal ICR mice and did successfully KD the expression of Lsd1 or Hotair by RNA interference (Fig. 4A). We tested the Lsd1, H3K4me2, and H3K9me2 levels by the Western blotting. LSD1 protein expressions were downregulated in the Hotair KD group and Lsd1 KD group (Fig. 4B). The expression levels of H3K4me2 had upregulated in the Hotair KD group and Lsd1 KD group (Fig. 4C). The result of the MTS assay showed that either KD of the expression of Lsd1 or Hotair could impede cardiomyocyte proliferation (Fig. 4D). We found that the cell cycle was arrested at the S phase after KD of the Lsd1 or Hotair expression (Fig. 4E, F).
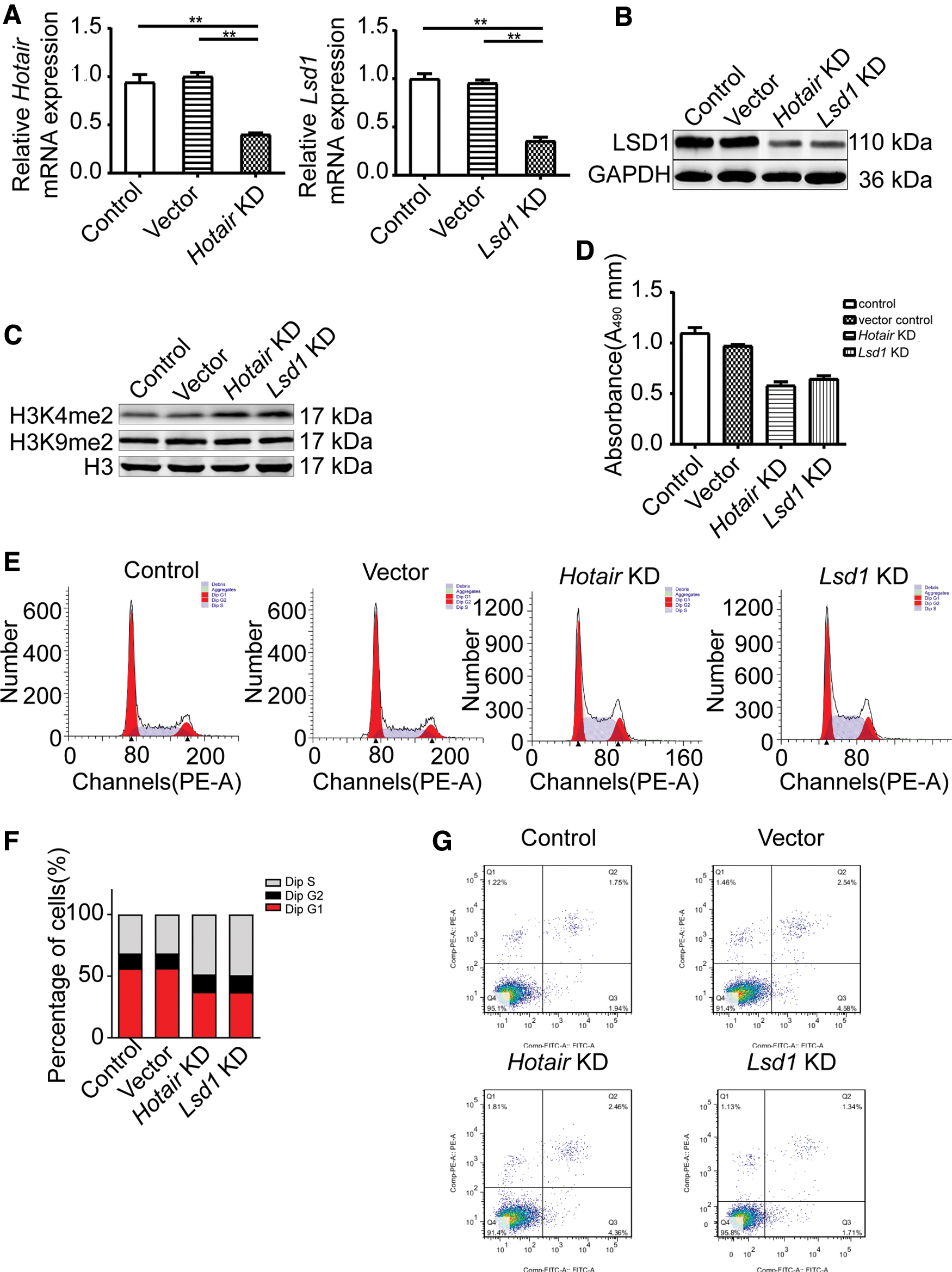
KD the expression of Hotair or Lsd1 could cause S phase arrest.
Interestingly, downregulating the expression of LSD1 or Hotair in neonatal cardiomyocytes do not affect the result of cell apoptosis in vitro (Fig. 4G).
Discussion
Regenerative treatment strategies are expected to improve heart function and prevent the exacerbation of heart failure. The mechanisms and cellular pathways involved in cardiomyocytes' endogenous regeneration still need to be explored.
Several research groups demonstrated that regulating endogenous regeneration was modulated by signal pathways, such as Hippo (Lin et al., 2014) NRG1 (D'Uva et al., 2015), etc. Recently, lncRNA has proved to play an important role in cardiac development, differentiation, lineage commitment, and heart regeneration through epigenetic regulation (Afify, 2019). The antisense lncRNA of silent information regulator factor 2-related enzyme 1 (Sirt1) could form a lncRNA/mRNA complex to improve the stability of the mRNA, and increase the protein level of sirt1 that could promote cardiac regeneration (Li et al., 2018a). AZIN2-SV, a lncRNA that interacts with miR-214, reduced the number of proliferating cardiomyocytes through the PTEN/PI3K/Akt pathway (Li et al., 2018c). Cardiomyocyte regeneration-related lncRNA could directly bind to miR-199a-3p and suppress heart regeneration (Chen et al., 2018). Endogenous cardiac regeneration-associated regulator markedly stimulated cardiac regeneration and promote the recovery of heart function after myocardial infarction (Chen et al., 2019). Accumulating research findings have shown that LncRNA is involved in the biological processes of cardiac regeneration. In our study, we found that Hotair may also regulate myocardial cell proliferation and myocardial regeneration.
Hotair, discovered by Howard Chang's group, has been intensively investigated as an oncogenic lncRNA in the field of cancer. Hotair, a promising therapeutic target, highlights an immense role in tumor growth, invasion, apoptosis, metastasis, cancer stem cell differentiation, and drug resistance (Qu et al., 2019). Hotair could also bind to the GA-rich motif, leading to PCR2-mediated trimethylation of histone H3 Lys 27 (H3K27me3) (Imai-Sumida et al., 2020). The protective effects of Hotair have been reported by Gao et al. (2019) Hotair, which functioned as a molecular sponge of miR-34a, could improve cardiac function in diabetic cardiomyopathy. Meanwhile, Hotair is reported as a potential therapeutic target for myocardial infarction by sponging miR-519d-3p (Zhang et al., 2019). The plasma concentration of Hotair could be a biomarker for human acute myocardial infarction (Gao et al., 2017). Hotair also involved in cardiac-associated sepsis, regulating myocardial TNF-α production through the NF-κB pathway (Wu et al., 2016). Hotair inhibition could provoke oxidative stress-induced injury through Hotair/miR-125/MMP axis in H9C2 cells (Li et al., 2018b). In cardiac fibroblasts, Hotair induces upregulation of fibrosis-related genes and promoted proliferation and migration of cardiac fibroblasts through Wnt signaling (Pan et al., 2018). However, Yang and Lu (2016) also reported that Hotair could promote hypoxia-induced ischemic infarct through NOX2 production upregulation; but little is known about the role of Hotair in cardiac regeneration. In the present study, we found Hotair highly expressed within 1 week after birth, consistent with the myocardial regeneration time window. Hotair plays a strong role in the regulation of endogenous regeneration in the neonatal mouse.
Hotair acts as a molecular scaffold for the assembly of the repressor complex (LSD1/CoREST/REST complex) consisting of LSD1. To look for the molecular mechanism underlying the neonatal myocardial regeneration, we further tested whether LSD1 also plays a role in the regeneration processes.
As a histone demethylase, LSD1 decreases trimethylation of histone H3 Lys 4 (H3K4me3) on target genes, leading to efficiently repress gene expression. LSD1 plays an important role in heart development partly through the control of E-cadherin phosphorylation (Nicholson et al., 2013). Activation of LSD1 also has a cardioprotective effect in ischemic disease (Wang et al., 2018). Interestingly, our data show that LSD1 expression strongly decreased after 14 days of birth, and KD of the LSD1 expression could impair the neonatal myocardial regeneration. These data strongly suggest that LSD1 could be a key gene responsible for the myocardial regeneration process.
Therefore, we have shown evidence that Hotair and LSD1 could regulate myocardial cell proliferation and myocardial regeneration. Hotair-mediated activation of the LSD1 signaling pathway may be a potential strategy to promote cardiac regeneration.
Footnotes
Authors' Contributions
The work presented in this study was carried out in collaboration among all authors. Q.F., M.Q., and S.Z. carried out the laboratory experiments. G.F. carried out analysis of the data. S.W. worked on animal breeding. B.Y. and L.Z. carried out the study design and article preparation. B.Z. performed cardiomyocyte isolation, culture, cell cycle, and apoptosis. Q.W. performed the immunofluorescence detection assay. All authors have contributed to, see, and approved the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Youth Project of the Tianjin Natural Science Foundation (16JCQNJC12100, 15JCYBJC25400); Tianjin Outstanding Youth Science Foundation (17JCJQJC46200); and the National Natural Science Foundation of China (82070687).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.