
Abstract
NAD is mainly biosynthesized by the enzymatic action of nicotinamide phosphoribosyltransferase (NAMPT) through the salvage pathway. NAD is indispensable for the proper function and metabolism of all living cells, including cancer cells. Our previous researches revealed that inhibition of NAMPT by miRNA (miR) could suppress NAD levels and thereby hinder the growth and promotion of breast cancer (BC). Therefore, the current study was undertaken to investigate the inhibitory effects of miR-613 on NAMPT and BC cells' survival. Bioinformatics analysis and luciferase reporter assay confirmed that NAMPT 3′-untranslated region is a direct target for miR-613. The expression of miR-613 was noticed to be significantly decreased in both clinical tissue samples and BC cells by real-time PCR. Following transfection with miR-613 mimic, the expression of miR-613 was elevated in the BC cells leading to inhibition of NAMPT expression at both mRNA and protein level as measured by real-time PCR and western blotting, respectively. Inhibition of NAMPT led to a remarkable reduction in the concentration of NAD in the BC cells. The transfection also declined cell viability roughly 40% in MD Anderson-Metastatic Breast-231 (MDA-MB-231) cells. Consistently, the apoptosis rate was remarkably increased, around 65% in these cells as assayed by labeling the cells with Annexin V-fluorescein isothiocyanate (FITC) and Propidium Iodide. Targeting the NAMPT-mediated NAD salvage pathway by miR-613 is a novel approach for managing BC, which is worth further investigation.
Introduction
Despite the massive efforts to screen and manage breast cancer (BC) within the past decade, it still continues to be one of the most serious health concerns worldwide. In this regard, BC is the most commonly diagnosed cancer (11.7% of total cases) in women and is the leading cause of cancer mortality (Sung et al., 2021). These concerning statistics highlight the importance of investigating novel and efficient methods for the management of BC. Suppressing cellular and molecular mechanisms associated with BC's expansion has been proposed as a crucial diagnostic and therapeutic approach (Bolandghamat Pour et al., 2019a; Zang et al., 2020).
NAD is mainly synthesized through the salvage pathway rather than de novo pathway in mammalian cells. It plays a pivotal role in a range of physiological and pathological activities, such as energy metabolism, cell death, and DNA damage repair. Nicotinamide phosphoribosyltransferase (NAMPT) acts as the rate-limiting enzyme in the salvage pathway. Abnormal high expression of NAMPT has been reported in several cancers such as renal, prostate, and BCs. Additionally, it has been claimed that secretion of NAMPT as a proinflammatory or anti-inflammatory cytokine from various cells, such as leukocytes, hepatocytes, and adipocytes, can enhance the proliferation and survival of BC cells through changing several signaling pathways (Behrouzfar et al., 2017; Gholinejad et al., 2017). Conversely, suppression of NAMPT remarkably declines metabolism and invigorates apoptosis in BC cell lines (Alaee et al., 2017; Hesari et al., 2018; Bolandghamat Pour et al., 2019a). Therefore, it can be considered as a promising approach to BC management.
miRNAs are highly conserved, single-stranded, endogenous, noncoding small RNAs consisting of 21–23 nucleotides. Mature miRNAs act as posttranscriptional regulators of gene expression by base pairing with mRNA molecules in their 3′-untranslated region (3′-UTR), controlling the translation process of the target mRNA (Nazarizadeh et al., 2020). As a recently discovered miRNA, miR-613 has been confirmed to be significantly involved in tumorigenesis of a wide variety of cancers. More importantly, the majority of available studies indicate that miR-613 remarkably suppresses the expression of oncogenes and malignant features of tumors meaning that it plays a tumor-suppressive role in several neoplasms (Mei et al., 2020). Notably, a pile of recently published articles suggest that downregulation of miR-613 is tightly associated with BC progression, and its forced overexpression can restrain BC development (Liu et al., 2020; Zang et al., 2020).
Our previous studies revealed that the NAMPT-mediated NAD salvage pathway significantly contributes to BC's promotion, and its suppression using miR-206, miR-381, and miR-154 could remarkably decrease the malicious nature of BC in Michigan Cancer Foundation-7 (MCF-7) and MD Anderson-Metastatic Breast-231 (MDA-MB-231) cells (Hesari et al., 2018; Bolandghamat Pour et al., 2019a, 2019b). Despite the tumor suppressive nature of miR-613, its effectiveness in NAMPT regulation has not been previously established. Therefore, this study was undertaken to investigate the value of miR-613 in BC patients and its effect on NAMPT expression and cell death in the BC cells.
Materials and Methods
Clinical samples
The tumor tissue and paired noncancerous margin specimens were taken from 30 patients who were referred to Rasoul-e-Akram Hospital (Tehran-Iran) during 2018–2020 for surgical resection. The freshly prepared samples were immediately snap frozen in liquid nitrogen and then stored at −80°C until analysis. All of the samples were confirmed by a pathologist who was blind to the research. The patients had not received chemo/radiotherapy before the sampling. All of the participants signed informed consent to be part of the study. Moreover, ethics approval was granted by the Institution Review Board (IRB) and Clinical Research Ethics Committee, Iran University of Medical Sciences, Tehran, Iran (Ethics ID No: IR.IUMS.FMD.REC.1400.120).
Cell lines and cell culture
Four cell lines, including MCF-7, MCF-10A, MDA-MB-231, and Human Embryonic Kidney 293T (HEK-293T), were procured from the Cell Bank of the Iranian Biological Resource Center (Tehran, Iran). The handling procedure was the same as our previous study (Bolandghamat Pour et al., 2019a). Briefly, STR profiling analysis was used to authenticate the cell lines. Bacterial and viral contaminations were also ruled out. MDA-MB-231 and MCF-7 cells were cultured in DMEM (Biosera, France). Moreover, MCF-10A and HEK-293T cells were cultured in mammary epithelial cell growth medium (Lonza/Clonetics, Switzerland) and DMEM/F12 (Biosera), respectively. All the culture media were supplemented with penicillin/streptomycin (1%) and FBS (10%). Additional supplements, including insulin (10 μg/mL), hydrocortisone (0.5 μg/mL), epithelial growth factor (20 ng/mL) and cholera toxin (100 ng/mL) were added to MCF-10A culture medium. The cells were kept in a humidified incubator at 37°C containing 5% carbon dioxide.
Target prediction
The following websites were used to check which miRNAs can potentially bind to NAMPT 3′-UTR: miRDB database (
Transfection of miR-613 mimic and inhibitor
The miR-613 level was manipulated following transfection with its sense and anti-sense oligonucleotides (mimic and inhibitor) (GenePharma, China). Further details regarding the miR-613 mimic, inhibitor, and negative control are shown in Table 1. The BC cells were transfected with 200 nmol/L of miRNA using polyethylenimine (PEI). Briefly, the cells were seeded into 6- or 96-well plates. One day later (24 h), after washing the cells, FBS- and antibiotic-free culture medium were added to the wells, and incubation continued for 1 h. Oligonucleotides and PEI were mixed in double-distilled water and allowed to form a complex at room temperature for 30 min. Finally, the cells were incubated for another 6 h after adding the mixture. Next, supernatants were removed, and fresh culture medium (containing 10% FBS) was added to each well, and maintained in a standard environment for 48 h. The quality and efficiency of the above procedure was evaluated by transfecting the cells with FAM-labeled (fluorescein isothiocyanate [FITC]) miRNA and examination of cells using flow cytometer after 24 h. The process was confirmed to be performed with high efficiency.
Sequences of miR-613 Mimic, miR-613 Inhibitor, and Their Respective Negative Controls
Luciferase assay
The 3′-UTR of human NAMPT, containing the putative target site of the miR-613 was synthesized and placed at the XbaI site in the pGL3-control vector (Promega, USA). Twenty-four hours before transfection, cells were seeded into 24-well plates (1.5 × 105 cells/well). Then, miR-613 mimic or inhibitor or its negative control, together with one of the following constructs, including NAMPT-3′-UTR-psiCHECK2, NAMPT-miRNA response element (MRE)-tandem-mut-psiCHECK2 (the vector containing the mutant MRE), or untransfected psiCHECK2 plasmid as the negative control were cotransfected into the HEK-293T cells using PEI. After 42 h, luciferase activity was measured using a Dual Luciferase Assay Kit (Promega), according to the manufacturer's guideline.
Real-time PCR for miR-613 and NAMPT measurement
Total RNA was extracted from the cells and tissue samples using Hybrid-R™ miRNA kit (GeneALL, South Korea), and was quantified by NanoDrop spectrophotometer. One microgram RNA was utilized to synthesize cDNA by EasyScript® One-Step gDNA Removal and cDNA synthesis SuperMix (TransGen, China). Before the synthesis of miR-613 cDNA, a poly (A) tail was added to the 3′-end of the miRNA transcripts by Escherichia coli Poly (A) Polymerase (PAP) (New England Biolabs, UK). Additionally, a primer containing a complementary sequence for the poly (A) tail and an adapter sequence was used in reverse transcription reaction (Table 2). Real-time PCR was conducted by RealQ Plus 2 × Master Mix Green High ROX™ (Amplicon, Denmark), employing U6 small nuclear RNA and GAPDH as internal controls to normalize the expression of miR-613 and NAMPT, respectively. Gene expression levels relative to the controls were calculated using the 2–ΔΔCT formula.
Sequences of Primers (Forward and Reverse) Used for Real-Time PCR
F, forward; NAMPT, nicotinamide phosphoribosyltransferase; R, reverse; RT, reverse transcription.
Cell survival assay
According to the manufacturer's guideline, the effects of miR-613 mimic and inhibitor on cell viability were assessed by tetrazolium-based WST-1 reagent (Roche Applied Science, Germany). Briefly, a cell suspension containing 5 × 103 cells/100 μL was seeded into a 96-well plate. Following overnight incubation at a standard environment, the cells were transfected with either miR-613 mimic or inhibitor and negative control. Next, WST-1 reagent (10 μL/well) was added to the wells, and after 4 h, optical density was read by a plate reader (BioTek Instruments Inc., Winooski, VT) at 450/650 nm.
Apoptosis assay
The effects of miR-613 on cell death were investigated by the BD FACSCalibur™ flow cytometer (BD Biosciences) using the Annexin V/Propidium Iodide (PI) Staining Kit (Roche Applied Science), according to the manufacturer's instructions at 488 nm excitation, a bandpass filter at 515 nm for FITC and 600 nm for PI detection. Briefly, 5 × 105 cells were seeded into a six-well plate and transfected with miR-613 mimic, inhibitor, and the negative control as described above. Cells were then harvested, washed twice with PBS, resuspended in Annexin V binding buffer, and stained with Annexin V and PI at room temperature in the dark for 30 min. Annexin V was used to detect both the early and late stages of apoptotic cells, whereas PI was used to detect late apoptotic and necrotic cells. Quantitative analysis of the FACS data was performed using the FlowJo software.
Western blotting
After lysis of the cells by RIPA buffer, the lysate was centrifuged at 16,000 × g for 10 min and then the supernatant was separated. Next, total protein level of the supernatant was estimated according to bicinchoninic acid assay (BCA) using a standard kit (Bio Basic, Canada). Sodium dodecyl sulfate/polyacrylamide gel electrophoresis 12% was employed to separate the samples. After the separation, the specimens were transferred onto polyvinylidene difluoride membranes (Thermo Fisher Scientific, USA). Then, 5% skimmed milk in Tris-Buffered Saline (TBS) 0.1% Tween-20 was used to block the membranes at 4°C, overnight. After incubation of membranes with anti-NAMPT antibody (PBEF/NAMPT Rabbit mAb; MyBioSource, USA) at a dilution of 1:1000 and anti-Rabbit HRP-conjugated secondary antibody (Abcam, USA) at a dilution of 1:5000, the specimens were visualized by Clarity Western ECL Substrate (Bio-Rad). The obtained bands were normalized against GAPDH bands (internal control) and analyzed by densitometry using ImageJ software (v1.52, NIH).
NAD level assay
NAD level was measured using the NAD/NADH assay kit (Abcam, UK) as described previously (Alaee et al., 2017). Briefly, the cell lysate was prepared using lysis buffer and total protein levels were measured using the BCA protein assay kit. Then, protein was removed from the samples by perchloric acid, and the NAD level was assessed by an enzymatic method. Absorbance was read at 450 nm, and the NAD levels were calculated by normalizing against the protein content.
Statistical analysis
All of the experiments were performed at least three times independently (except for the western blot, which was done two times). Statistical analysis was carried out using a software (GraphPad Prism version 8.0.2, San Diego, CA), and numerical data were presented as mean ± standard deviation. Multiple comparisons were conducted by analysis of variance, followed by the post hoc Tukey's test. The tissue expression of miR-613 was compared between the cancerous and marginal samples using the Student's t-test. p-Values <0.05 were considered statistically significant.
Results
MiR-613 was downregulated in BC cells and tissues
MiR-613 expression was first evaluated in cancerous and noncancerous marginal tissues by quantitative real-time PCR, and the results are depicted in Figure 1. As indicated, miR-613 was significantly downregulated in the tumor samples compared with the margins (Fig. 1A). Following clinical data, miR-613 expression was measured in cancer cell lines (MCF-7 and MDA-MB-231) and compared with that in MCF-10A normal breast cells and found to be remarkably lower in the MCF-7 (p < 0.0001) and MDA-MB-231 (p < 0.0001) cell lines compared with MCF-10A cells. As presented in Figure 1B, miR-613 expression levels were noticed to be around 2.5-fold in MCF-7 and 3-fold in MDA-MB-231 lower than MCF-10.

Basal expression levels of miR-613 in the clinical tissue samples
MiR-613 was upregulated by miR-613 mimic
Flow cytometry analysis revealed that more than 77% and 93% of MCF-7 and MDA-MB-231, respectively, were successfully transfected with labeled miR-613 mimic (Fig. 2). The level of miR-613 was manipulated in MCF-7 and MDA-MB-231 cells by transfecting with miR-613 mimic, inhibitor, and negative control. Following transfection with the miRNA mimic, the expression level of miR-613 was significantly elevated in both cell lines, whereas transfection with the miR-inhibitor had no significant effects on miR-613 levels (Fig. 3).

Flow cytometry analysis shows successful transfection of MDA-MB-231 [

The relative expression levels of miR-613 after transfection with its mimic and inhibitor, in MCF-7
NAMPT 3′-UTR was the target of miR-613
Bioinformatics analysis revealed that miR-613 could potentially target the NAMPT gene. MiR-613 was found to align with the position 2653–2659 of NAMPT 3′-UTR with 7mer-m8 exact match to the mature miRNA (Fig. 4A). To confirm this finding, HEK-293T cells were transfected with miR-613 mimic or inhibitor, negative control, and luciferase reporter vectors containing the putative NAMPT 3′-UTR target site. The obtained results demonstrated that the luciferase activity of NAMPT 3′-UTR in miR-613 mimic-transfected cells was significantly lower than the control (Fig. 4B). Meanwhile, the miR-613 inhibitor could significantly increase the luciferase activity in these cells compared with the control. Neither mimic nor inhibitor could significantly influence the luciferase activity of the mutated form of the miR-613 target in the 3′-UTR of NAMPT or psiCHECK2 plasmid alone.
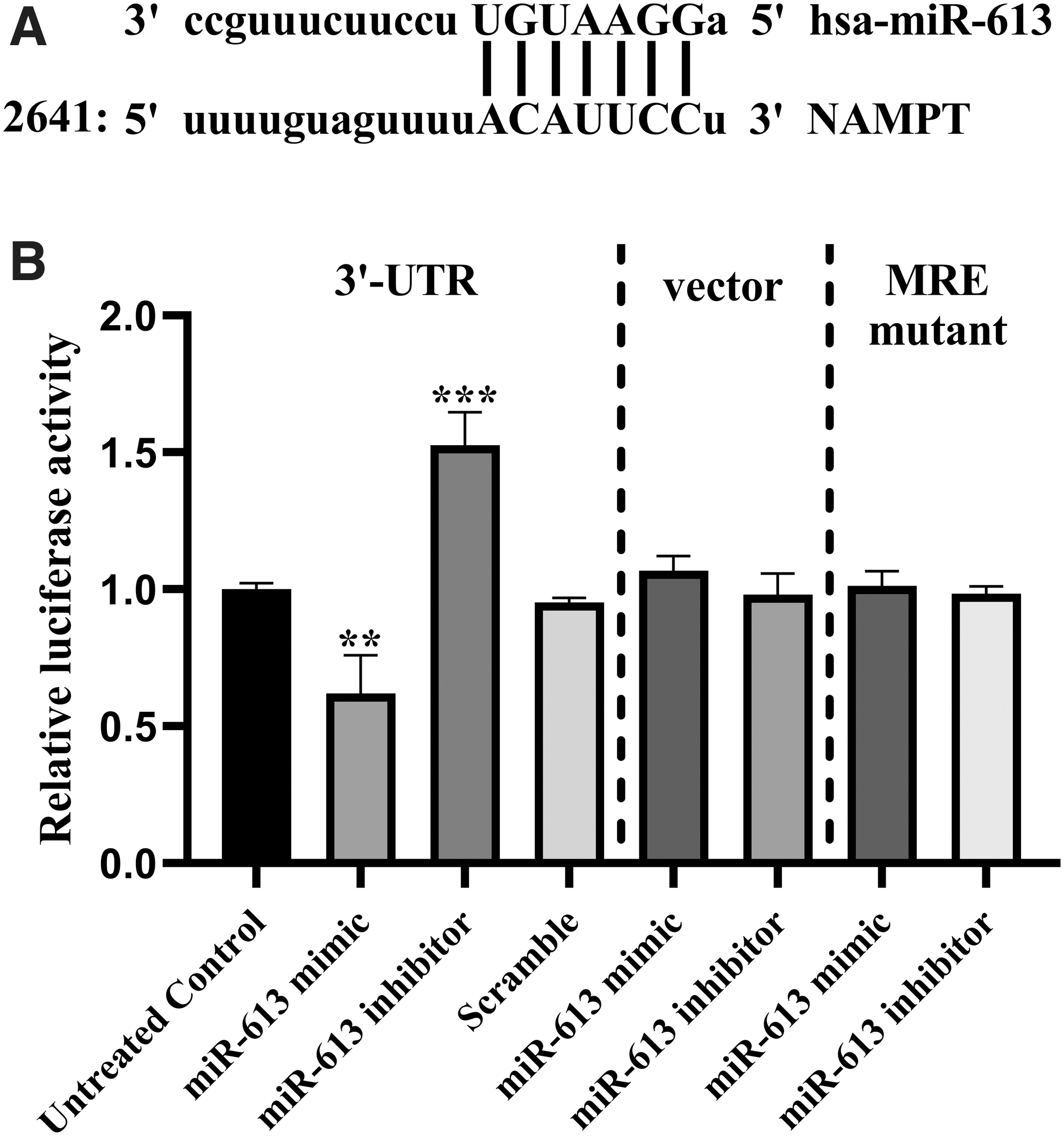
NAMPT is the target gene of miR-613. A. predicted alignment of miR-613 with the 3′-UTR sequence of NAMPT mRNA. B. reduction of luciferase activity of the construct containing 3′-UTR of NAMPT mRNA by miR-613 mimic and its augmentation by miR-613 inhibitor as determined by dual luciferase assay after cotransfection of wild-type NAMPT 3′-UTR with miR-613 oligonucleotides into HEK-293T cells. The unchanged vector and the vector containing the mutant MRE were used as negative controls. A scramble oligonucleotide was also used as negative control. The results were compared with untreated control group. **p < 0.01, ***p < 0.001. MRE, miRNA response element; NAMPT, nicotinamide phosphoribosyltransferase; UTR, untranslated region.
Upregulation of miR-613 suppressed NAMPT expression
The effect of miR-613 upregulation on NAMPT was evaluated both at the gene and protein levels. Forced overexpression of miR-613, using its mimic, could significantly suppress NAMPT gene expression in MCF-7 and MDA-MB-231 cells compared with the control. However, the miR-613 inhibitor had no significant effect on NAMPT gene expression in both cancer cell lines (Fig. 5A, B).
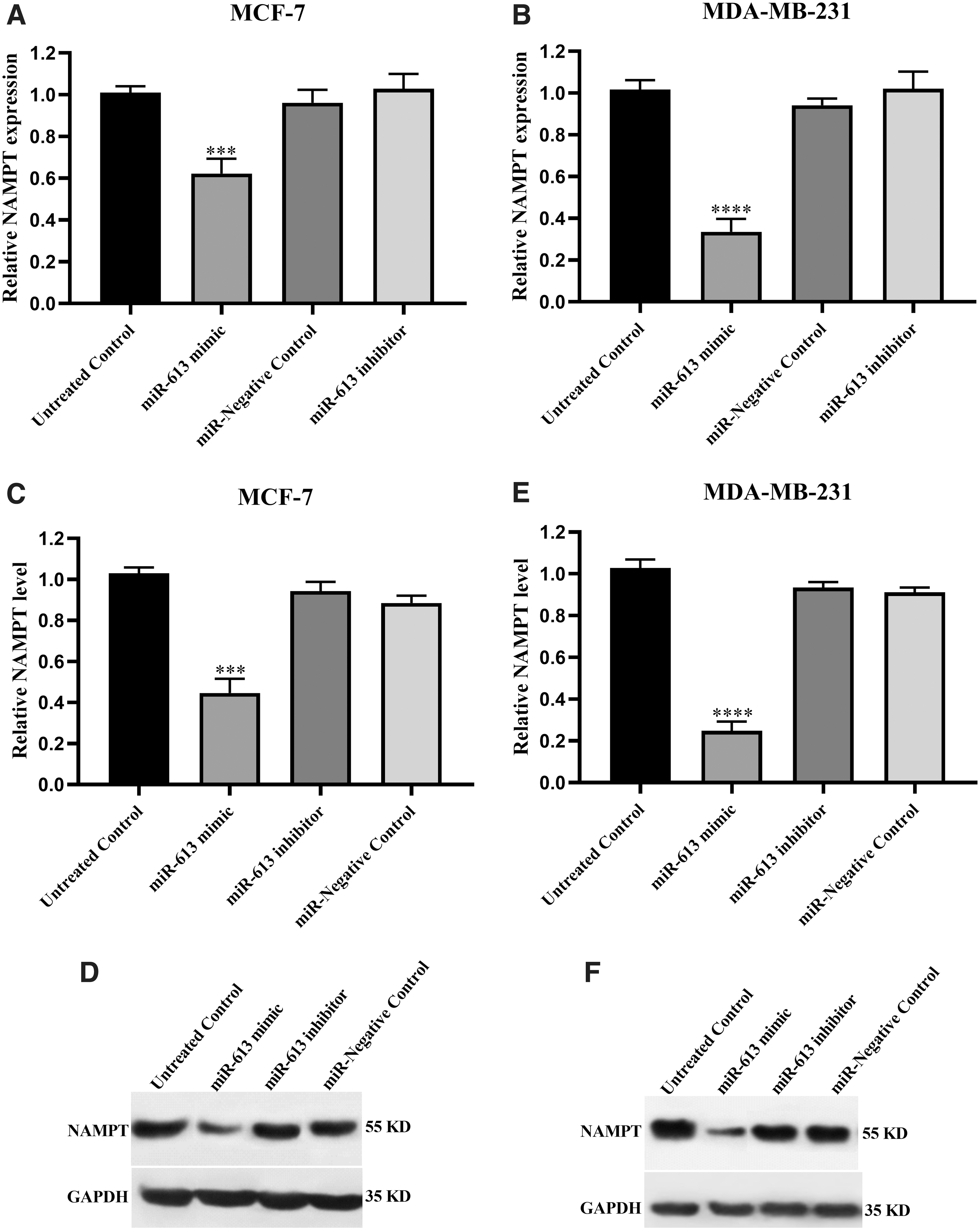
The effect of miR-613 transfection on NAMPT expression. The upper panels show mRNA expression of NAMPT in MCF-7
Parallel to these findings, western blotting revealed that miR-613 mimic could significantly repress NAMPT protein expression in MCF-7 and MDA-MB-231 cell lines compared with the untreated cells. More specifically, about 1.5-fold and 3-fold decreases were recorded in MCF-7 and MDA-MB-231, respectively. The protein levels of NAMPT did not change significantly in response to miR-613 inhibitor in both cell lines (Fig. 5C–F) (Supplementary Data).
miR-613 overexpression decreased NAD levels
The effects of miR-613 on NAD levels as the main product of NAMPT enzyme was investigated. Forced overexpression of miR-613 using its mimic oligonucleotide resulted in a significant reduction of NAD in MCF-7 and MDA-MB-231 cells compared with the control. The reduction rate was calculated to be ∼1.25-folds for MCF-7 and 2.5-folds for MDA-MB-231 (Fig. 6). The inhibitor of miR-613 did not exert any significant effect on NAD levels in either of cancer cells.
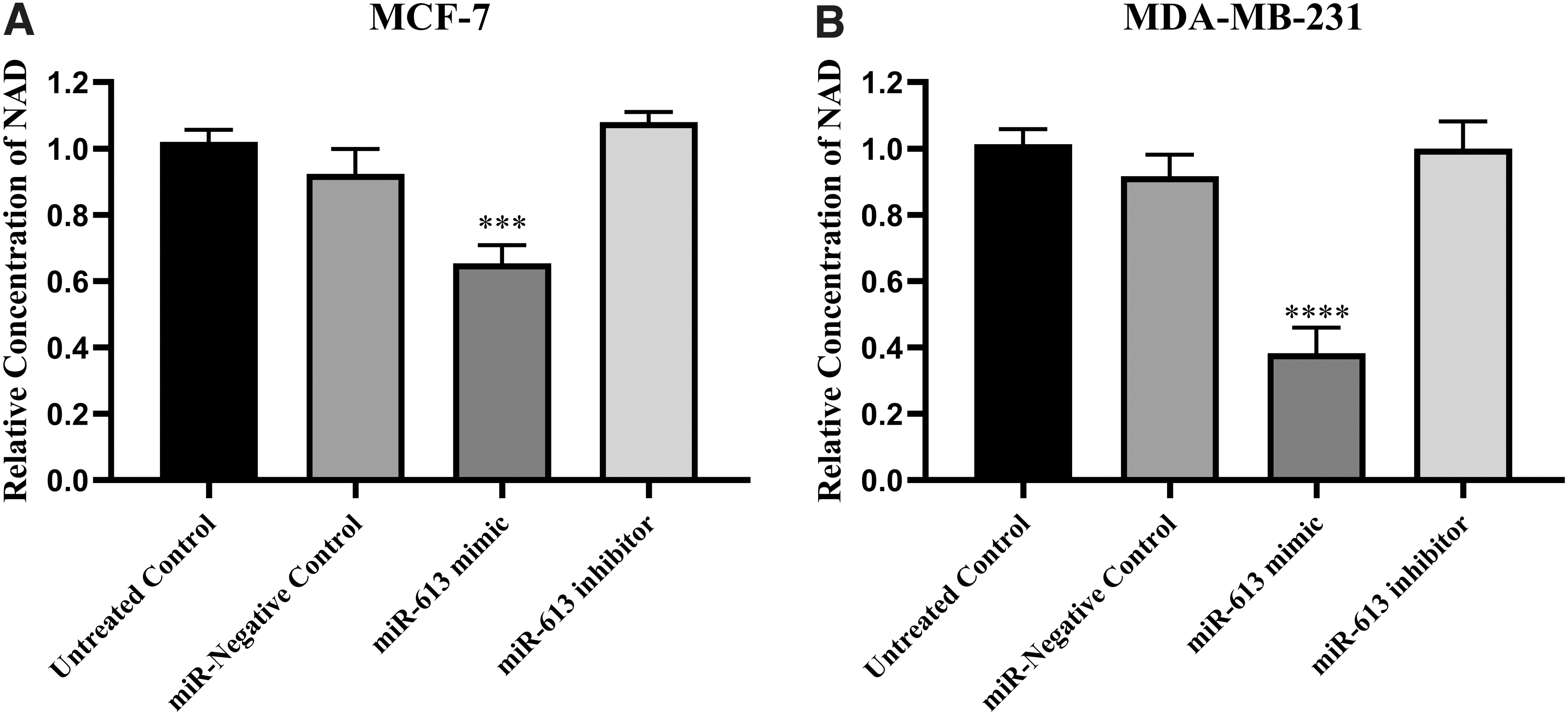
Cellular NAD levels in breast cancer cell lines MCF-7
miR-613 overexpression promoted cell death
Transfection with miR-613 mimic significantly accelerated cell death of the studied cell lines. As can be seen from Figure 6, the cell viability, assessed by WST-1 assay, was determined to be roughly 75% in MCF-7 and 40% in MDA-MB-231, which was significantly lower than the control cells (Fig. 7). Increased NAMPT expression level in cancerous cells might invigorate the progression of BC through elevating NAD levels, whereas its suppression can lead to reduced cell survival through boosting NAD production (Ghorbanhosseini et al., 2019). To test this hypothesis, transfected cells were treated with NAD (1 mM) after the end of the 6-h incubation period of transfection, and their viability was evaluated. Addition of NAD was able to revert the survival of both cancer cells back to their normal levels, confirming that the reduction of NAD by miR-613 was mainly responsible for its effect on cell viability.
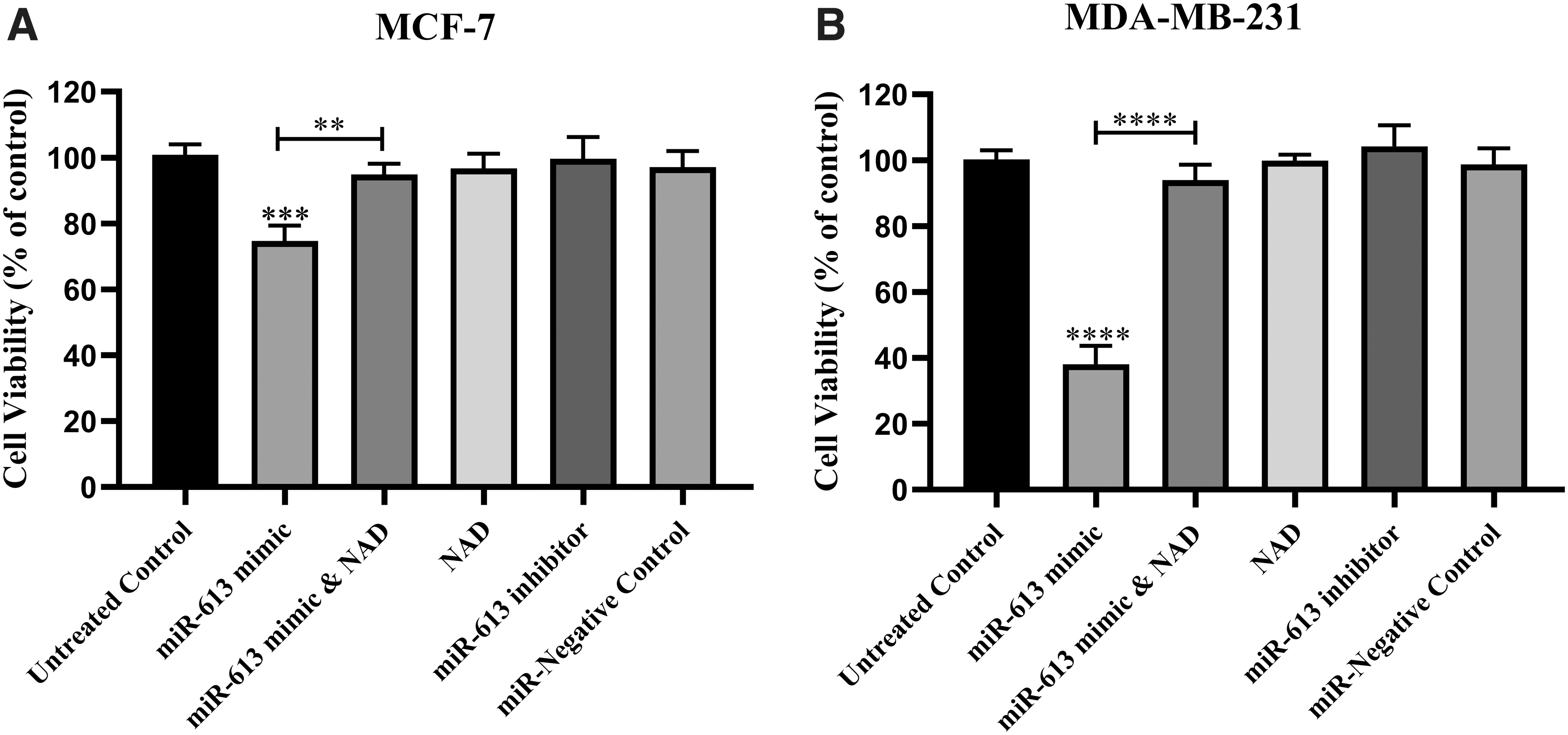
The effect of overexpression of miR-613 on the cell survival of MCF-7
In line with the above findings, miR-613 mimic could induce apoptosis in cancer cell lines. As indicated in Figure 8, the population of apoptotic cells was found to be 25% in MCF-7 and 65% in MDA-MB-231, which was significantly higher than the control cells. Combining NAD with miR-613 mimic led to a significant decrease in apoptosis, compared with the miR-mimic alone. Meanwhile, miR-613 inhibitor had no significant effects on cell death compared with the untreated control.
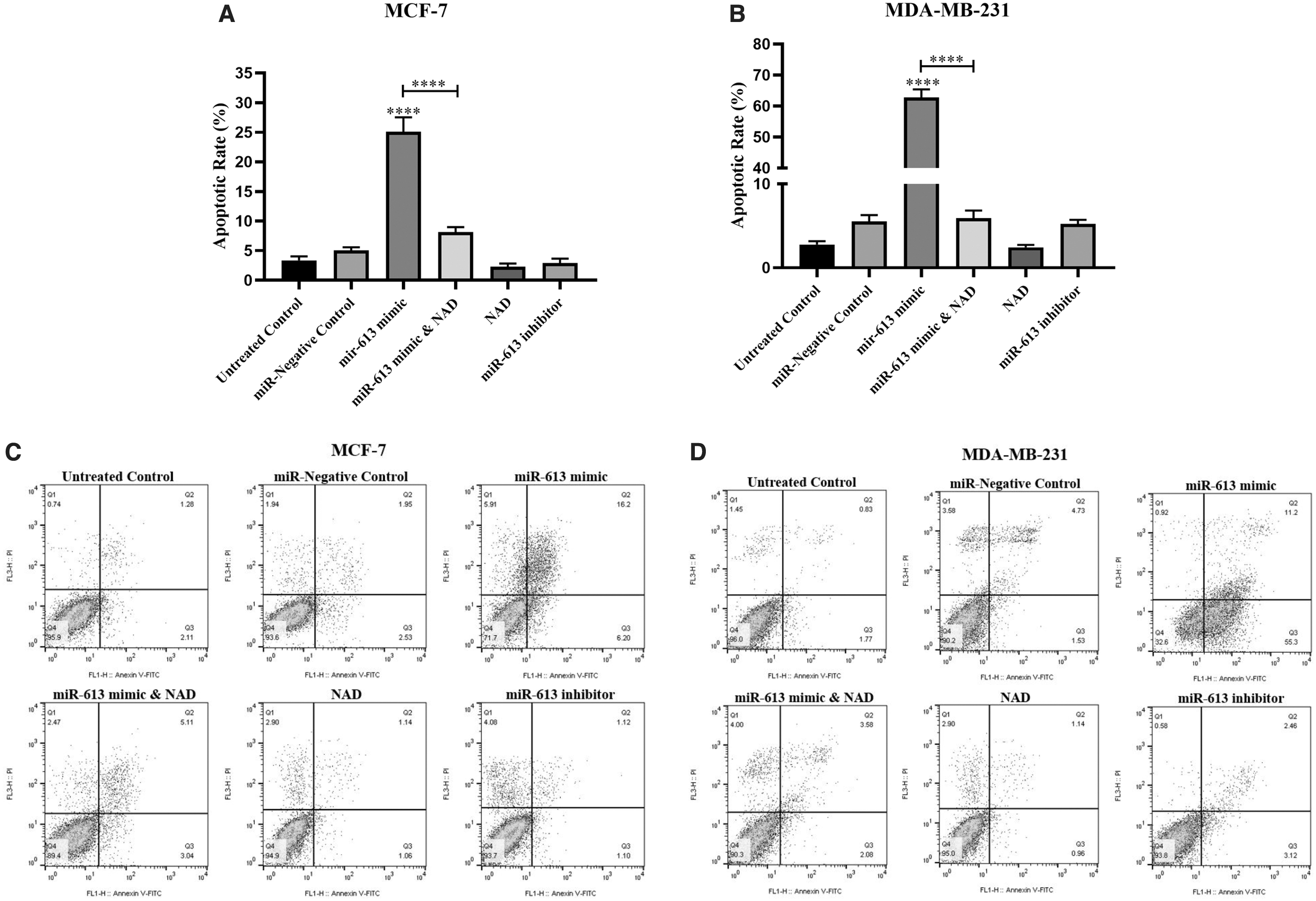
The average population of apoptotic cells in MCF-7
Discussion
BC is still responsible for remarkable death and economic losses worldwide, and understanding the molecular mechanisms beyond BC development might lead to improvement of the treatment strategies and reduce BC mortality (Zhou et al., 2013). NAMPT has been introduced as a key regulator of BC growth and metastasis (Sheikhpour, 2017). MiRs have recently been proposed to be involved in tumorigenesis and progression of BC; therefore, they could be considered as an innovative tool for diagnosis and treatment of BC (Hesari et al., 2018; Liu et al., 2020; Peng et al., 2020; Shibuya et al., 2020; Li et al., 2021). We previously presented miRNAs as effective tools to promote apoptosis and reduce cell viability in BC cells by inhibiting NAMPT activity and subsequently decreasing NAD levels (Alaee et al., 2017; Hesari et al., 2018; Bolandghamat Pour et al., 2019a, 2019b). In this study, for the first time, using the bioinformatics analysis and luciferase reporter assay, we discovered that miR-613 targets NAMPT and vastly diminishes both mRNA and protein levels of NAMPT, leading to the reduction in NAD levels. We also noticed that miR-613 significantly promoted cell death. This study revealed that miR-613 has a tumor-suppressive role in BC, which agrees with previous studies (Li et al., 2016; Sang et al., 2018; Ji and Hu, 2020).
In the present research, we illustrated that miR-613 was significantly downregulated in the BC tissue samples compared with the adjacent tissues, suggesting miR-613 as a tumor suppressor of BC. The downregulation was also observed in the BC cell lines compared with MCF-10A (epithelial cells), and interestingly the levels of miR-613 in the triple negative breast cancer (TNBC) more invasive cell line (MDA-MB-231) were significantly lower than the non-TNBC counterpart (MCF-7). All of these findings are consistent with the previous studies (Xiong et al., 2018; Liu et al., 2020; Li et al., 2021).
Our team's previous research disclosed that NAMPT mRNA was significantly higher in MCF-7 and MDA-MB-231 cells than MCF-10A cells (Hesari et al., 2018). In the present study, we confirmed that the transfection of BC cell lines with miR-613 mimic significantly diminished cellular NAMPT at the mRNA and the protein levels. This finding proved that NAMPT expression regulation through miR-613 occurs both at the transcriptional and translational level, possibly through degradation of NAMPT mRNA (Wang et al., 2016). The effect of numerous miRNAs in modulating cellular NAMPT levels was determined in different studies. Raised level of miR-154 and miR-381 in BC cell lines repressed NAMPT mRNA expression and protein levels (Bolandghamat Pour et al., 2019a, 2019b). Parallel with our findings, Zhang et al. (2013) and Adyshev et al. (2014) documented remarkably reduced NAMPT expression at both protein and mRNA levels following overexpression of miR-26b in colorectal cancer cell lines and miR-374a and miR-568 in human lung endothelial cells. Moreover, in support of our study, it has been proven that NAMPT 3′-UTR is a potential target for multiple miRNAs in various cancers (Zhang et al., 2013; Adyshev et al., 2014; Bolandghamat Pour et al., 2019a, 2019b) and other disorders, including HIV-1-contaminated cells (Chen et al., 2013), neonatal sepsis (Li et al., 2018), and obesity (Choi et al., 2013).
NAD recycling is critical to cancer cells' longevity, and blocking the NAD salvage pathway leads to metabolic collapse and cell death (Ju et al., 2016). NAMPT is significantly involved in the biosynthesis of NAD, and therefore its inhibition can be considered a suitable approach to reduce intracellular NAD (Bolandghamat Pour et al., 2019a). Cell survival and apoptosis are tightly correlated with NAMPT activity and this enzyme also provides NAD as the substrate for SIRT1, which is required for cell longevity. Conversely, decreased SIRT1 activity due to reduced NAD level strongly correlates with promoted apoptosis (Bolandghamat Pour et al., 2019b). In this study, we demonstrated that the inhibition of NAMPT and the subsequent reduction in NAD levels could suppress cell viability and promote apoptosis. We had previously shown that inhibition of NAMPT by its specific inhibitor could effectively reduce NAD levels and induce apoptosis in BC cells (Alaee et al., 2017). In support of our findings, researches have demonstrated that inhibition of NAMPT by miRNAs is an efficient way of modulating NAD content and cell viability (Choi et al., 2013; Zhang et al., 2013; Ju et al., 2016).
In the current study, treatment of the cells with NAD could neutralize the effects of miR-613 mimic on cell viability and apoptosis verifying that the apoptosis-inducing effect of miR-613 is mainly exerted through inhibition of NAMPT and the subsequent decline in cellular NAD content.
Conclusion
In summary, our experiments demonstrated that miR-613, which is diminished in patients with BC, serves as a tumor suppressor by directly targeting NAMPT such that its overexpression could decrease viability and induce apoptosis of BC cells through reducing intracellular NAD. Therefore, miR-613 can be considered as a potential target for the invention of novel treatments against BC. However, more clinically oriented studies are still required to clarify its usability as an anticancer agent.
Footnotes
Disclosure Statement
The authors declare that there is no conflict of interest.
Funding Information
This research was financially supported by grants from the Iran University of Medical Sciences (Grant No. 97-02-30-3298) and Iran National Science Foundation (Grant No. 97013570).
Supplementary Material
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.