
Abstract
The sperm-associated antigen 5 (SPAG5) is an important protein in mitosis and cell cycle checkpoint regulation, with more attention as a novel oncogene in various cancers. High level of SPAG5 expression has been detected in our clinical gastric cancer (GC) samples and The Cancer Genome Atlas GC data. However, the bio-function and potential mechanism of SPAG5 in GC remain unclear. In this study, we investigated the role of SPAG5 in GC development and the correlation between SPAG5 and 5-fluorouracil (5-FU) treatment. SPAG5 expression was increased in GC samples compared with that in normal tissues (80.8% vs. 22.0%), which was apparently associated with a worse outcome. Biological experiments showed that knockdown of SPAG5 induced apoptosis and suppressed proliferation in cells and animal models. Downregulation of SPAG5 enhanced the sensitivity of 5-FU in GC cells. Gene microarray chip identified 856 upregulated and 787 downregulated genes in SPAG5 silencing cells. Furthermore, 12 significant genes, including CDKN1A, CDKN1B, EIF4E, MAPK1, and HSP90B1, belonged to the PI3K/AKT signaling pathway using ingenuity pathway analysis. Meanwhile, real-time PCR and Western blotting results showed that knockdown of SPAG5 inhibited PI3K/AKT signaling pathway. Collectively, SPAG5 promotes the growth of GC cells by regulating PI3K/AKT signaling pathway, which could be the promising target gene in GC therapy.
Introduction
Gastric cancer (GC) is one of the most malignancies in China (Chen et al, 2016; Yang, 2006). Even though the current treatment of GC has been improved, it is still challenging based on the complex etiology with more interactions between environmental factors and genetic variations, as well as the difficulty in early detection of GC. Therefore, it is of great significance to investigate the mechanisms underlying GC progression.
Sperm-associated antigen 5 (SPAG5/Astrin) is a component of mitotic spindle localized to ch17q11.2 (Chang et al, 2001), which is mainly expressed in sperm cells, but hardly expressed in other normal tissues. Recent studies have shown that SPAG5 functions as the essential regulator of mitosis (He et al, 2020). Dysregulation of SPAG5 promotes the progression of different types of cancers, including, hepatocellular carcinoma (Liu et al, 2018; Yang et al, 2018), breast cancer (Zhou et al, 2019), bladder urothelial carcinoma (Wang et al, 2018), cervical cancer (Yuan et al, 2014), and GC (Liu et al, 2019) and the overexpression of SPAG5 was analyzed in gastric cancer (n = 408) and normal (n = 211) samples from TCGA database. However, the clinical significance and function of SPAG5 are poorly understood in GC. We will further illustrate the role of SPAG5 in GC development.
Hence, we detected the abundance of SPAG5 in GC and normal tissues, and explored the potential molecular mechanism of SPAG5 using gene microarray chip, real-time PCR, Western blot analyses, colony formation, chemo-resistance, and animal model analyses by knocking down of SPAG5 in GC cells.
Materials and Methods
Clinical samples
This study was approved by the Affiliated Hospital of Qinghai University and the 7th medical center of PLA General Hospital. One hundred twenty-two patients with GC were enrolled from January 2012 to December 2017 at 7th medical center of PLA General Hospital. None of the patients received any therapy intervention before surgery. The study was approved by the Affiliated Hospital of Qinghai University (2020.1-2023.12) and the 7th Medical Center of PLA General Hospital Institutional Review Board (2021-90). Each patient of this study obtained the informed consent for specimen preservation. Informed consent was written by each patient.
Cell culture and RNA interference
MKN45 and HGC27 cells were grown in Dulbecco's modified Eagle's medium (Corning), which contained 10% FBS. Using the commercial shSPAG5 or shCtrl empty vectors (GV 112; Genechem, Shanghai, China), we stably transfected MKN45 cell lines with Lipofectamine 2000 (Invitrogen, Carlsbad, CA). Forty-eight hours after transfection, stable cell lines were constructed by incubating with 400 μg/mL of G418 (Genview) for 3 weeks. Furthermore, SPAG5 was also interfered by siRNAs, which were synthesized from GenePharma. Targeted sequence was as follows: siSPAG5#1 sense, 5′-CAGAAUCUGCUUCACCUCUUUdTdT-3′ and antisense, 5′-AAAGAGGUGAAGCAGAUUCUGdTdT-3′; siSPAG5#2 sense, 5′-GCAGCAGAUUUCCGUGUCAAUdTdT-3′ and antisense, 5′-AUUGACACGGAAAUCUGCUGCdTdT-3′; and siCtrl sense, 5′-UUCUCCGAACGUGUCACGUdTdT-3′ and antisense, 5′-ACGUGACACGUUCGGAGAAdTdT-3′.
Microarray analysis
To dissect gene expression regulated by SPAG5, total RNA was extracted from shCtrl and shSPAG5 cells by using Trizol, following the indicated protocols. After detecting the quantity and quality of total RNA, three parallels of knockdown group and control group were subjected to mRNA microarray analysis on 100 format Affymetrix Gene Chip. Bioinformatics analysis of the results was analyzed from CapitalBio, according to the protocols.
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay
Cell growth was examined by using 3-(4,5-dimethylthiazol-2-Yl)-2,5-diphenyltetrazolium bromide (MTT) assay (Genview JT343). shCtrl and shSPAG5 cells were plated in 96-well plates for the concentration of 2000 cells per well (3–5 repetitions per group). At the indicated time, 20 μL (5 mg/mL) of MTT were added into each well and the plates were maintained at 37°C for 4 h. Then OD value at 490/570 nm was measured on the Microplate reader (Tecan infinite M2009PR; Switzerland).
Colony formation
Cell growth was also determined by colony formation assay. In brief, 400–1000 stable transfected and control MKN45 and HGC27 cells were plated in 6-well plates. After growing for 2 weeks, cell colonies were formed and were viewed by staining with GIEMSA (AR-0752; DingGuo, Shanghai, China) and photographed.
mRNA detection
RNA was extracted from indicated cells (80% cell density in 6-well plates) using Trizol (Invitrogen), following the indicated instructions. Stem-loop reverse transcription (RT) primers were synthesized by (Ribio, Gunagzhou, China), (SPAG5-F:5′-TTGAGGCCCGTTTAGATACCA-3′, SPAG5-R:5′-GCTTTCCTTGG AGCAATGTAGT T-3′). cDNA was prepared using cDNA RT Kit (M-MLV; Promega). cDNA was quantified by using SYBR master mix on the qPCR system. The primer sequences for qPCR were as follows: SPAG5 forward, 5′-CTGAGCAGTAGAACTGAGGCT-3′ and reverse, 5′-TCCACATGATTGACACGGAAAT-3′. β-actin forward, 5′-GAGCTGCGTGTGGCTCCC-3′ and reverse, 5′-CCAGAGGCGTACAGGGATAGCA-3′.
Tissue microarray construction and immunohistochemistry
To assess the expression of SPAG5, tissue microarray containing 174 primary GC specimens and 50 normal tissues was collected from the tumor bank of Affiliated Hospital of Qinghai University and the 7th medical center of PLA General Hospital. The sections were subjected to immunohistochemistry (IHC) staining of SPAG5 by using EnVision+ assay kit following the introductions. SPAG5 antibody was purchased from Abcam (ab184142, 1:50 dilution). Staining intensity was scored by two independent pathologists. The percentage of positive cells was calculated as follows: 0, negative; 1, 10% ≤ positive cells; 2, 10% < positive cells ≤50%; 3, 50% < positive cells ≤75%; and 4, 75% < positive cells. After multiplying the two scores, the result of 0 was scored as negative, 1–4 was scored as weak, 5–8 was scored as moderate, and 9–12 was scored as strong. Median cutoff point of SPAG5 was 4. Patients were subdivided into SPAG5 high and low expression group.
Western blotting
Cells were lysed in RIPA buffer (Beyotime, China). Protein concentration was measured by BCA Protein Assay Kit (P0010S; Beyotime). Total proteins were loaded on SDS-PAGE gels and immunoblotted onto PVDF membranes. Immunodetection was performed using a rabbit anti-SPAG5 antibody (1:1000 dilution, #60940; CST), anti-pAKT antibody (1:1000 dilution, #4060; CST), anti-AKT antibody (1:1000 dilution, #4685; CST), anti-PTEN antibody (1:1000 dilution, #9188; CST), anti-EIF4E antibody (1:1000 dilution, #2067; CST), anti-CDKN1A antibody (1:1000 dilution, #2947; CST), and mouse anti-GAPDH (1:2000 dilution, SC-32233; Santa Cruz, CA).
Xenografted tumorigenesis
A total of 20 male mice (BALB/c-nu, 5-week old, 18–20 g) were obtained from Beijing Weitonglihua Experimental Animals Company (China). Animal experiments were conducted according to the Guidelines of the Care and Use of Laboratory Animals (National Institutes of Health [NIH] publications nos. 80-23, revised 1996) and the institutional guidelines. Animal study was approved by the Ethics Committee of Qinghai University (2020-04). All mice were divided into two groups (10 mice/group) randomly. In brief, 5 × 106 of shCtrl and shSPAG5 MKN45 cells were subcutaneously transplanted into Balb/c nude mice (female, 4-week old).
The length (L) and width (W) of the tumors were monitored every 5 days after tumor formation. Tumor volume was calculated following the formula, V = L × W 2 × 0.5. All the animals were anesthetized by intraperitoneal injection with 3% pentobarbital sodium (0.04 mg/g per mice) and were euthanized by peritoneal injection of chloral hydrate (0.4 mg/g per mice). The xenografted tumorigenesis was performed one time.
Statistical analysis
All the statistical data were analyzed with GraphPad prism 6.0 software. Chi-Square test was used to analyze clinical data. Kaplan–Meier survival plots were applied to analyze patients' survival. Student's t-test was applied to compare the difference between two groups. Statistical significance was considered when p < 0.05.
Results
High level of SPAG5 expression in GC
To explore the clinical relevance of SPAG5 in GC, we performed IHC staining to assess the expression of SPAG5 in GC tissues. Around 80.8% (139/172) of GC samples showed higher level of SPAG5 compared with normal tissues (22.0%, 11/50) (Fig. 1A and Table 1). There was no association between SPAG5 expression and gender (p = 0.076), TNM stage (p = 0.340), tumor invasive depth (p = 0.359), lymph node metastasis (p = 0.510), and distant metastasis (p = 0.613), whereas it was correlated with the Lauren's classification (p = 0.001) and age groups (p = 0.048) (Table 2). In multivariate analysis, SPAG5 expression (p = 0.002), tumor invasive depth (p = 0.036), lymph node metastasis (p = 0.018), and the Lauren's classification (p = 0.045) were independent risk factors for the overall survival of GC patients (Fig. 1B and Table 3).
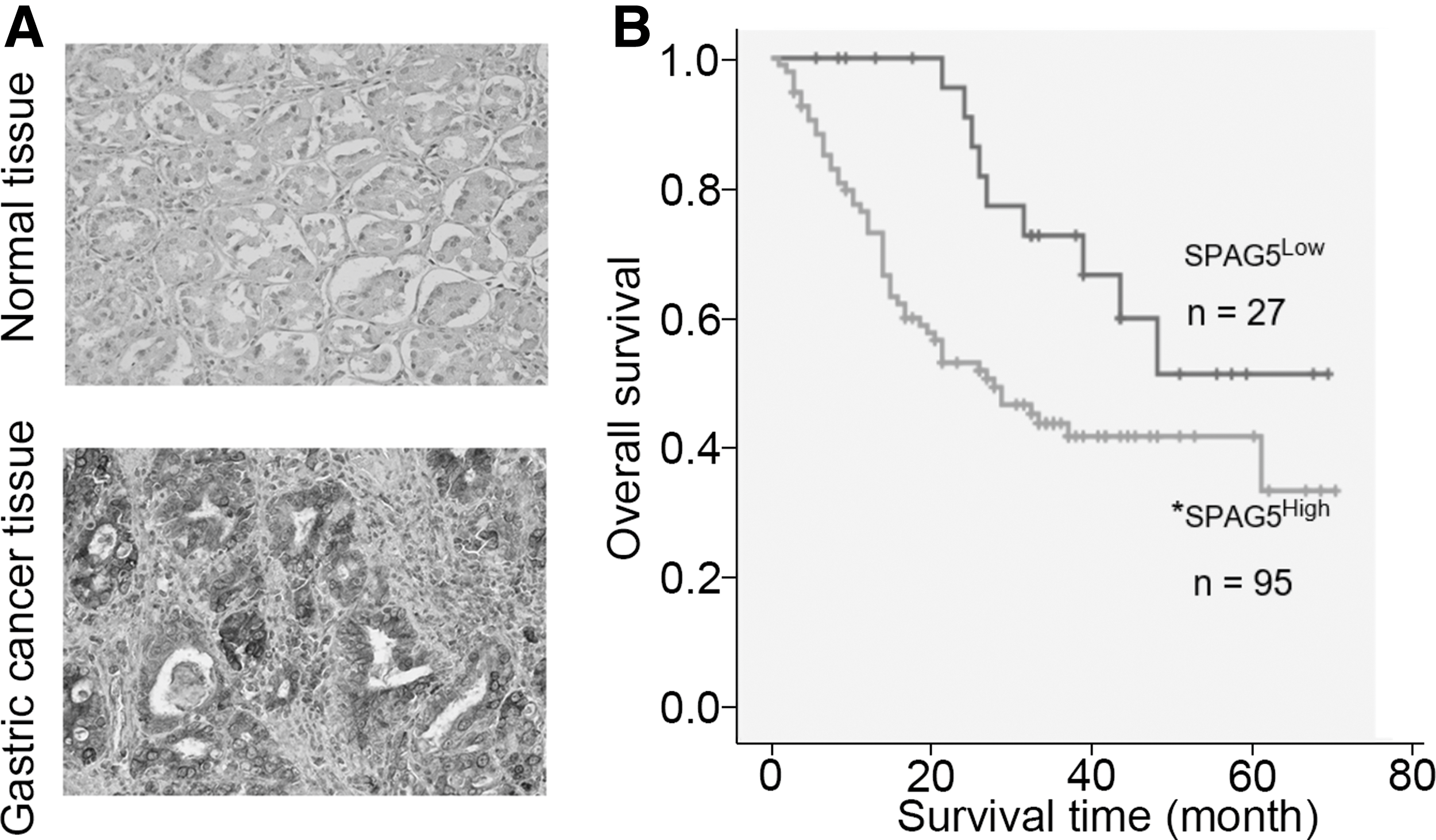
IHC staining of SPAG5 expression in clinical samples.
The Different Expression of Sperm-Associated Antigen 5 in Gastric Cancer and Normal Stomach Tissues
GC, gastric cancer; SPAG5, sperm-associated antigen 5.
The Correlation Between Sperm-Associated Antigen 5 Expression and Other Clinical Features in Gastric Cancer Patients
The Multivariate COX Regression of Sperm-Associated Antigen 5 and Other Clinical Features in Gastric Cancer
CIs, confidence intervals.
Downregulation of SPAG5 suppresses cell proliferation in vitro
Next, we used siRNA to knock down SPAG5 in HGC27 and MKN45 cells. Two siRNA sequences were used to avoid off-target effect. qRT-PCR and immunoblotting results confirmed knockdown efficiency (Fig. 2A, B). MTT results demonstrated that SPAG5 downregulation obviously reduced the proliferation ability of HGC27 and MKN45 cells (Fig. 2C, D). Likewise, SPAG5 silencing reduced colony formation in HGC27 and MKN45 cells (Fig. 2E, F). Apoptosis of both cells was enhanced after SPAG5 downregulation (Fig. 2G, H). Cell cycle was arrested at G0/G1 phase by SPAG5 reduction in HGC27 and MKN45 cells (Fig. 2I, J). Furthermore, SPAG5 knockdown also suppressed the migration of HGC27 cells (Fig. 2K, L). Taken together, SPAG5 contributes to GC cell proliferation, growth, and migration in vitro.
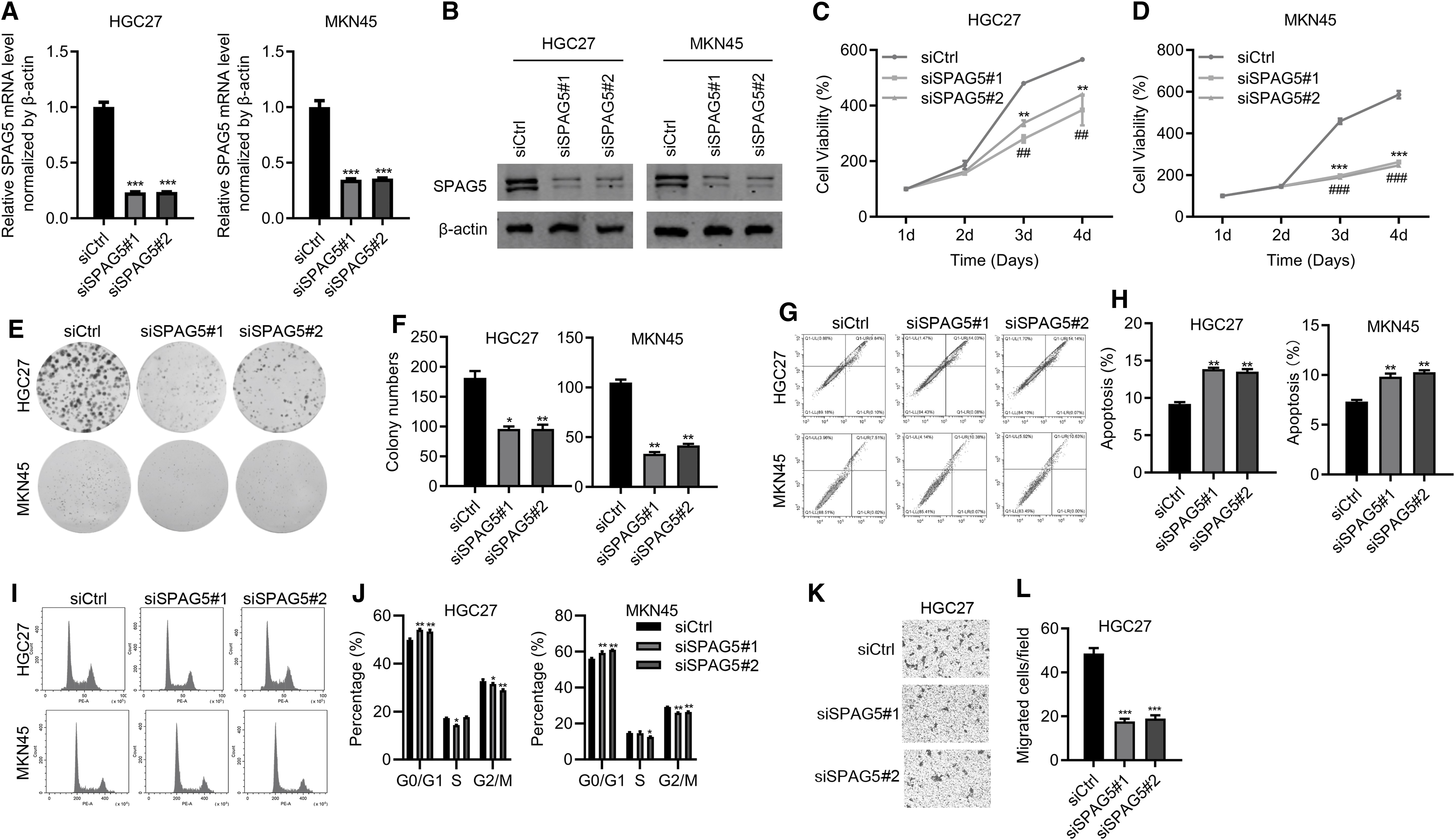
Downregulation of SPAG5 suppresses cell proliferation in vitro.
Downregulation of SPAG5 suppresses xenografted tumorigenesis
To investigate the in vivo function of SPAG5 in tumor-bearing models, MKN45 cells carrying lentivirus expressing shCtrl and shSPAG5 were subcutaneously implanted into female nude mice (4 weeks, 10/group). After tumors initiating 22 days using tumor subcutaneously implanted, tumor volume was measured every 5 days. All the mice were euthanatized later. The results showed that when shCtrl cells developed visible and large tumors, shSPAG5 cells formed small ones (Fig. 3), which indicated that SPAG5 knockdown suppressed tumorigenesis of MKN45 cells in nude mice.
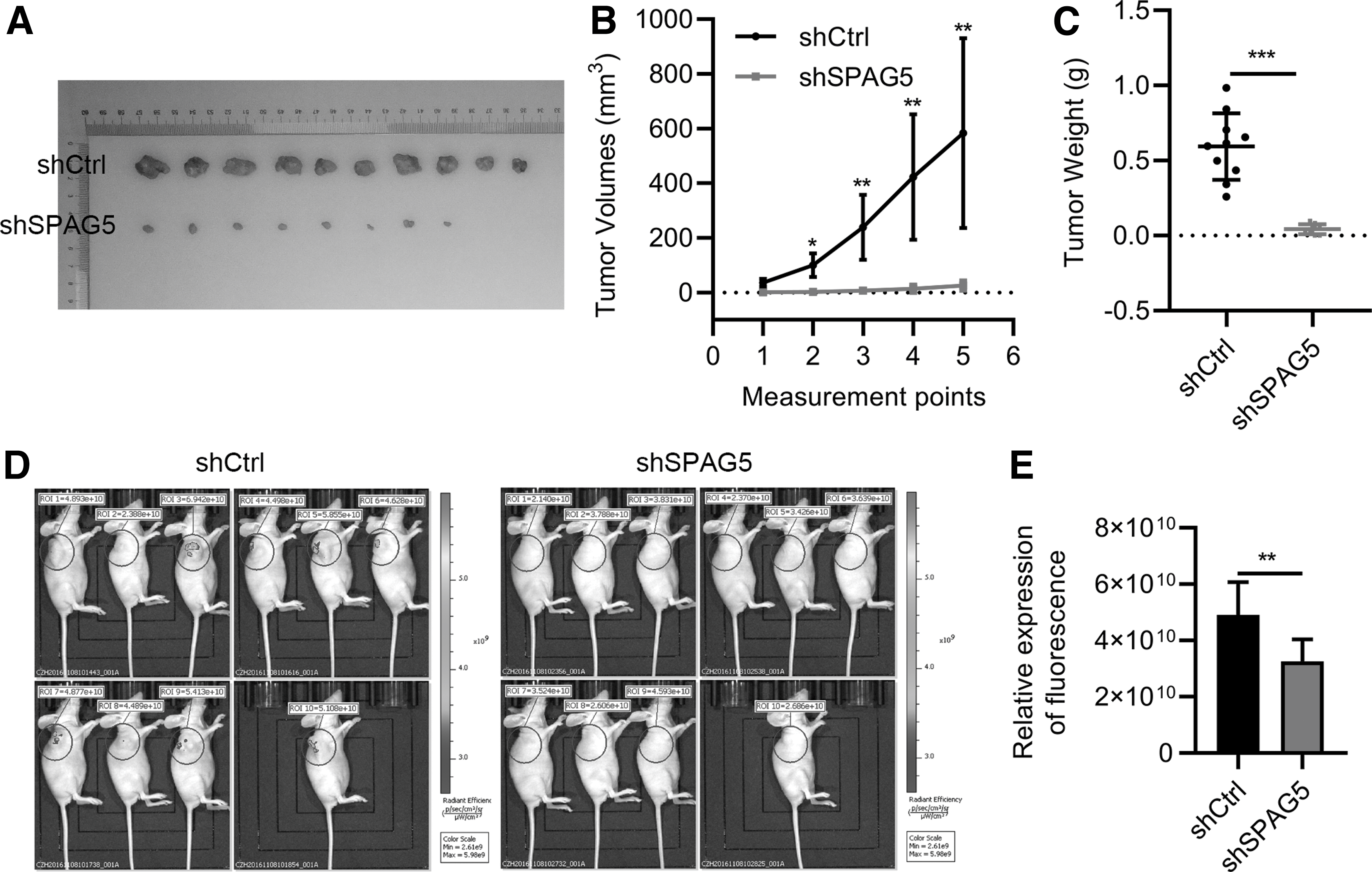
Downregulation of SPAG5 suppresses xenografted tumorigenesis. shCtrl and shSPAG5 MKN45 cells were subcutaneously transplanted into nude mice (female, 4-weekold). The mice were euthanatized 42 days later.
Expressing profiling of dysregulated genes after knockdown of SPAG5
To study the downstream molecular mechanisms of SPAG5, we used the gene microarray chip to measure the expression pattern of genes between shCtrl- and shSPAG5-transfected MKN45 cells. Comparing with shCtrl cells, 856 genes were upregulated and 787 genes were downregulated in shSPAG5 cells (Fig. 4A). We then utilized ingenuity pathway analysis, signaling pathway analysis, and the result showed that most of the significant genes regulated by SPAG5 were enriched in ERK/MAPK signaling and PI3K/AKT signaling (Fig. 4B). PI3K/AKT signaling pathway is a well-known oncogenic signaling pathway and we performed qRT-PCR and immunoblotting to validate these changes. qRT-PCR results showed that CDKN1A, CDKN1B, and PTEN were upregulated, while EIF4E was downregulated after SPAG5 knockdown (Fig. 4C). Immunoblotting results found that AKT phosphorylation was reduced (Fig. 4D). Collectively, SPAG5 knockdown suppressed PI3K/AKT signaling in MKN45 cells.
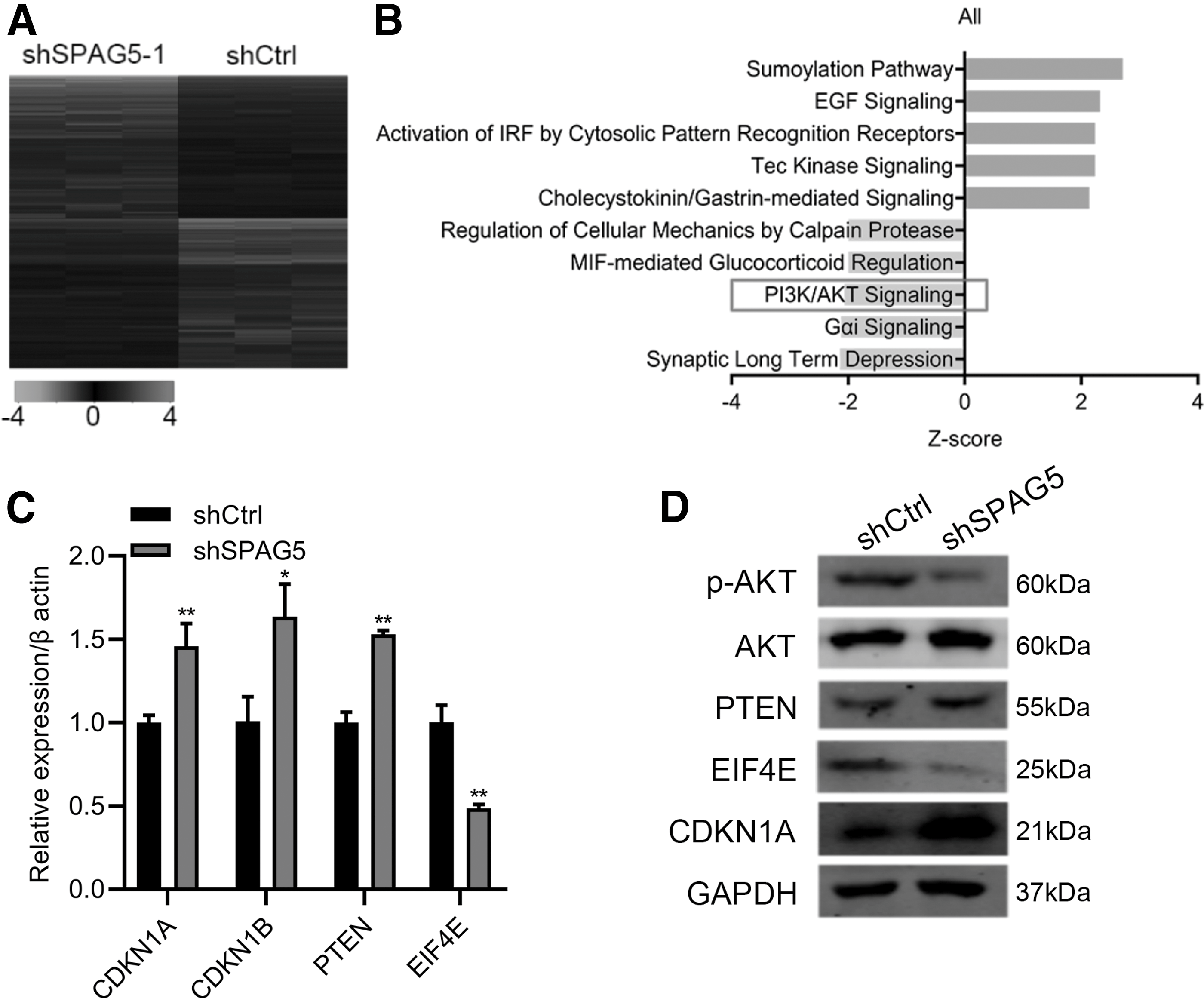
The expression pattern of genes in MKN45 cells transferred with shSPAG5 and shCtrl.
Knockdown of SPAG5 increases the sensitivity of 5-fluorouracil in GC cells
5-fluorouracil (5-FU) is a regular chemotherapeutic drug for GC patients. To explore whether SPAG5 expression is associated with the sensitivity of 5-FU in GC, siCtrl, and siSPAG5, GC cells were treated with different dosage of 5-FU. Cell viability was examined 48 h after treatment. The results showed that SPAG5 downregulation decreased IC50 of 5-FU in HGC27 and MKN45 cells (Fig. 5A, B). Then siCtrl and siSPAG5 cells were treated with dimethyl sulfoxide (DMSO) and 5-FU. The cell viability was measured at different times. The results showed that, comparing with siCtrl cells, siSPAG5 significantly increased the inhibitory effect of 5-FU on the proliferation ability of HGC27 and MKN45 cells (Fig. 5C, D).
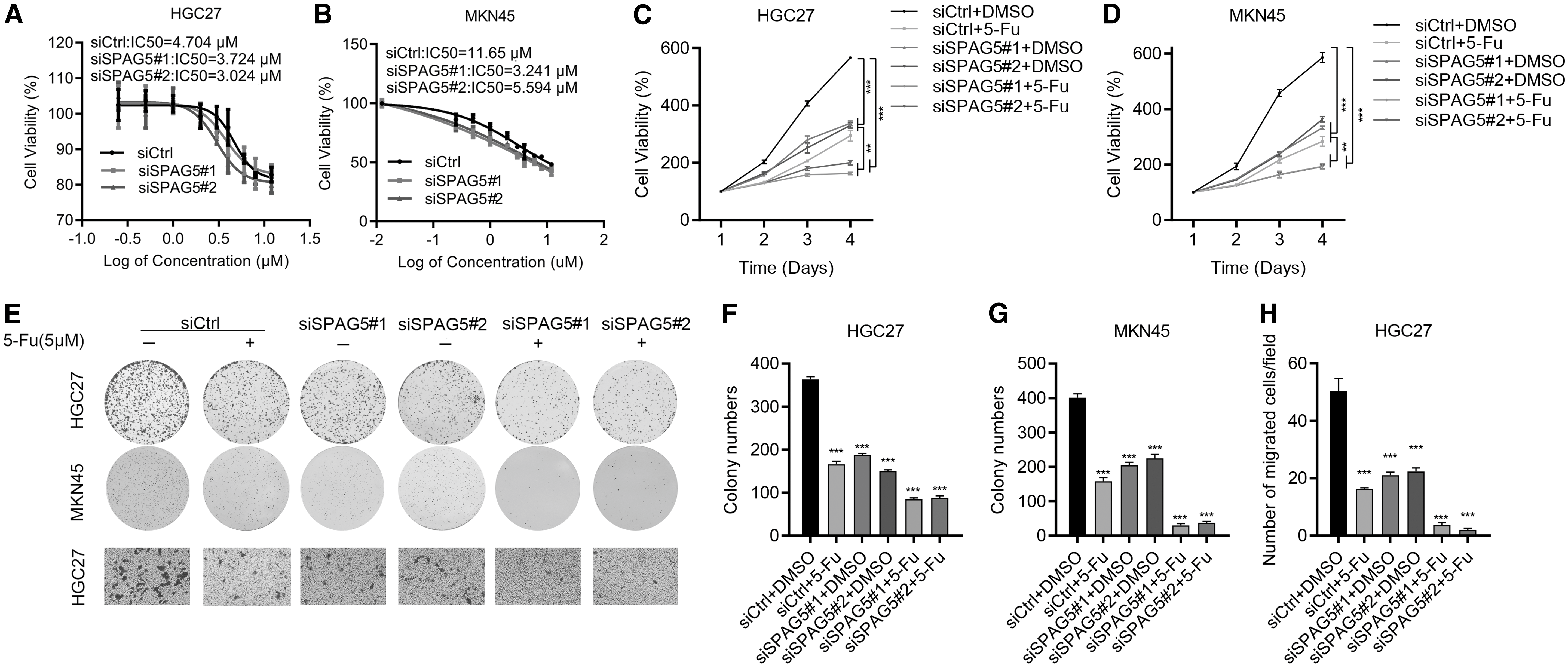
SPAG5 knockdown enhances 5-Fu effectiveness in GC cells.
To validate the results, we treated siCtrl and siSPAG5 cells with DMSO and 5-FU, and subjected the cells to colony formation and transwell assays. Consistent with the MTT results, SPAG5 knockdown dramatically enhanced the toxicity of 5-FU on the growth and migration of GC cells (Fig. 5E–H). These data suggest that siSPAG5 enhances the sensitivity of 5-FU in GC treatment. Thus, SPAG5 can be used as a potential biomarker for indicating the efficacy of 5-FU in GC patients.
Discussion
In this study, we identified that SPAG5 was an oncogene in GC and was correlated with the treatment effectiveness of 5-FU in GC cell. First, SPAG5 was highly expressed in GC tissues and was inversely correlated with the prognosis of GC patients. Then, loss-of-function experiments demonstrated that SPAG5 was critical for GC cell proliferation and migration. Molecular profiling identified that SPAG5 positively regulated PI3K/AKT signaling pathway. Finally, downregulation of SPAG5 enhanced the sensitivity of 5-FU for treatment of GC cells on cell growth and migration. Our results showed that SPAG5 could be an oncogene and drug target for GC.
SPAG5 is a microtubule-related protein that contributes to spindle formation during mitosis. As a centriolar satellite protein, SPAG5 promotes the translocation of CDK5RAP2 to centrosome by interacting with CEP72 and CDK5RAP2 (Kodani et al, 2015). In addition, SPAG5 can also interact with MCPH-related proteins to promote their translocation to centrosome, resulting in centriole duplication (Kodani et al, 2015). Since SPAG5 is closely associated with mitosis, dysregulation of SPAG5 is correlated with cancer development. For example, SPAG5 high expression is positively correlated with the poor prognosis of hepatocellular carcinoma and cervical cancer (Liu et al 2018; Yuan et al, 2014). SPAG5 promotes lung cancer development by suppressing P53 (Wang et al, 2019). SPAG5 activation of FOXM1/MMP2 contributes to the metastasis of osteosarcoma (Li et al, 2020a).
In breast cancer patients who received systematic therapy, the expression of SPAG5 is a promising biomarker to predict the prognosis (Zhu et al, 2019). SPAG5 also contributes to breast cancer progression by activating Wnt/β-catenin signaling pathway (Jiang et al, 2019). Moreover, another study demonstrated that SPAG5 was transcriptionally regulated by TEAD/YAP/TAZ network and post-transcriptionally regulated by miR-10b-3p in breast cancer (Canu et al, 2021). Transcriptional and post-transcriptional overexpression of SPAG5 supported the malignant growth of breast cancer. These results suggest that SPAG5 functions as an emerging oncogene in cancers (He et al, 2020).
In this study, we showed that SPAG5 inhibition triggered cell apoptosis and decreased cell growth in vitro and in vivo. Eight hundred fifty-six upregulated and 787 downregulated genes were found after SPAG5 knockdown. In addition, PI3K/AKT signaling pathway was positively regulated by SPAG5 in GC cells. Since PI3K/AKT is frequently activated and contributes to the development of various cancers, including GC (Hennessy et al, 2005; LoRusso, 2016; Wadhwa et al, 2013; Worby and Dixon, 2014), we propose that SPAG5 promotes GC cell proliferation and migration, at least partly, by potentiating PI3K/AKT signaling pathway.
We also showed that SPAG5 knockdown promoted the expression of PTEN, a well-known tumor suppressor gene and an inactivator of PI3K/AKT (Lee et al, 2018). In addition, SPAG5 knockdown suppressed the expression of EIF4F, overexpression of which has been shown to contribute to the activation of PI3K/AKT pathway (Culjkovic et al, 2008). These results indicate that SPAG5 could activate PI3K/AKT through negative regulation of PTEN and positive regulation of EIF4F. However, whether SPAG5 had a direct effect on the activity of PI3K/AKT signaling pathway needs further experiments to address.
GC represents one of the most lethal malignancies in both females and males worldwide. Besides surgery, chemotherapies are the preferential treatment strategies for GC. The 5-FU is a promising chemotherapy drug in GC (Ilson, 2018). Recently, numerous studies have shown that molecular changes, such as oncogene amplification or activation, or tumor suppressor inactivation, is associated with the treatment effectiveness of 5-FU in GC. For example, Helicobacter pylori CagA protein reduces 5-FU efficacy in GC cells by activating glucose metabolism (Gao et al, 2020). Suppression of β-glucosidase decreases 5-FU resistance by inducing lysosomal dysfunction (Li et al, 2020b).
Moreover, cell cycle is also related to 5-FU resistance in GC cells (Kim et al, 2020). Nevertheless, the significance of SPAG5 in 5-FU sensitivity in GC is to be determined. In this study, we showed that SPAG5 downregulation overcomes the resistance of 5-FU in GC cells. SPAG5 silencing enhanced the efficacy of 5-FU in cell proliferation and migration. Thus, SPAG5 could be targeted to increase the sensitivity of 5-FU for GC patients.
Conclusion
Collectively, SPAG5 potentiates GC malignancy through activation of PI3K/AKT signaling. Silencing of SPAG5 enhances the effect of 5-FU on GC cell treatment. Thus, SPAG5 is a potential antitumor target in GC.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants from the Key R&D and transformation program of Qinghai Province-Special project of science and technology assistance (no. 2021-QY-213), the Chinese and Tibetan Medicine Innovation Research Project NO: J2022015 the CAS (Light of the West China) Program (no. 2019-33), the National Natural Science Foundation of China (no. 81460429), the Open Project of State Key Laboratory of Plateau Ecology and Agriculture, Qinghai University (no. 2019-ZZ-07), and the Scientific Research Project of Research Team of Medical College of Qinghai University (no. 2020-KYT-2).
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.