
Abstract
Twenty-two functional α-disintegrin and metalloproteinases (ADAMs) have been identified in humans, 12 of which have proteolytic activity. The role of ADAMs in cancer has attracted increasing attention. However, the expression and significance of ADAMs in lung adenocarcinoma (LUAD) remain unclear. Most recently, we investigated the transcriptional data of ADAMs and related overall survival in patients with LUAD based on several databases, including TCGA, cBioPortal, Kaplan-Meier Plotter, LinkedOmics, KEGG, TIMER, and TISIDB. Knockdown of ADAM12 was performed in vitro to verify its biological function. According to our findings, 10 ADAMs exhibited significant differential expression in LUAD compared with cancer-adjacent normal tissues. ADAM12 expression was significantly higher in LUAD tissues than in paracancerous tissues, and lower ADAM12 expression was associated with better survival. Genetic alterations of ADAM12 mainly included missense mutations, amplifications, and deep deletions. ADAM12 and positively correlated genes were mainly enriched in protein digestion and absorption, extracellular matrix-receptor interaction, and adhesion plaques. ADAM12 had a moderate correlation with immune cell markers EBIP1, CCNB1, EXO1, KNTC1, PRC1, and FAM198B. Prognostic model was established based on ADAM12 and immune-related genes. In vitro experiments revealed that knocking down ADAM12 inhibited cell proliferation, migration, and invasion. ADAM12 potentially plays an important role in the occurrence of LUAD and may be utilized as an immunotherapy target and a valuable prognostic biomarker for LUAD.
Introduction
Lung cancer is the most common cause of cancer-related death worldwide, with 1.8 million new cases and 1.6 million deaths out of all cancer types. Approximately 85% of cases suffer from non-small cell lung cancer (NSCLC) (Bray et al., 2018). NSCLC mainly includes lung adenocarcinoma (LUAD) and lung squamous cell carcinoma (LUSC), with the former being most frequent (Liu et al., 2015; Osmani et al., 2018). LUAD is common among nonsmokers, women, and East Asian patients.
Over the past 20 years, the introductions of targeted therapies and immunotherapy have benefited certain patients with advanced NSCLC. However, LUAD treatment remains challenging, necessitating the identification of new driver genes to expand the patient population benefiting from targeted therapy. Furthermore, new predictors of immunotherapy response are also required.
Twenty-two functional α-disintegrin and metalloproteinases (ADAMs) have been identified in humans, but only 12 of these have proteolytic enzyme activity (ADAM8, 9, 10, 12, 15, 17, 19, 20, 21, 28, 30, and 33) (Jones et al., 2016). ADAMs are involved in a variety of biological processes, including cell adhesion, intercellular signal transduction, extracellular matrix (ECM) dynamics, and cellular transport (Blobel, 2005; Li et al., 2019).
The role of ADAMs in cancer has attracted increasing attention. Studies have shown that ADAM8 upregulates miR-720 expression, which in turn promotes triple-negative breast cancer (TNBC) metastasis by regulating the β1-integrin/ERK signaling pathway, and is thus expected to serve as a biomarker for the early detection of ADAM8-positive TNBC or a predictor of treatment response (Das et al., 2016). ADAM17 expression is generally high in gastric cancer, colon cancer, liver cancer, breast cancer, NSCLC, and other cancer types, promoting tumor metastasis through Notch and Wnt signaling (Shen et al., 2016; Li et al., 2019; Lu et al., 2019; Pelullo et al., 2019; Yang et al., 2019).
In lung cancer, ADAM9, ADAM19, and ADAM23 are highly expressed, while ADAM28 expression is low (Ota et al., 2016; Lin et al., 2017; Mochizuki et al., 2018; Wang et al., 2019). ADAM12 is highly expressed in a variety of tumors, promoting invasion and metastasis (Huang et al., 2018; Luo et al., 2018; Wang et al., 2018). Bioinformatics analysis of LUAD previously revealed that the high expression of ADAM12, BTK, and ERG is mainly related to adhesion plaques and the activation of immunemodulatory pathways (Yue et al., 2019). However, no report has focused on ADAM12 in LUAD.
In recent years, a great number of studies have demonstrated that the tumor immune microenvironment (TIM) plays a vital role in cancer progression and therapeutic efficacy. The tumor microenvironment (TME) generally consists of tumor cells, immune cells, and nonimmune cell subsets (e.g., fibroblasts, pericytes, and endothelial cells), as well as ECM proteins, cytokines, and growth factors (Rossi et al., 2020). ADAMs belong to the superfamily of zinc-dependent metalloproteinases, which process and degrade ECM proteins, growth factor-binding proteins, and cell adhesion molecules, as well as cell surface receptors (Sternlicht and Werb, 2001; Shiomi et al., 2010). Therefore, ADAMs have major implications within the TME.
In a variety of cancers, matrix metalloproteinases (MMPs) and ADAMs cleave the NKG2D ligand from the surface of tumor cells, preventing its interaction with the NKG2D receptor on the surface of natural killer (NK) cells. As a result, the receptor undergoes endocytosis and degradation, while tumor cells escape NK cell-mediated death (Groh et al., 2002). Wu et al. (2019) found that, within the TME, ADAM17 specifically cleaves activated FcγR CD16A on NK cells, causing them to degranulate, which in turn compromises their tumor-killing effect. This observation suggests that ADAM17 may be a potential target for cancer immunotherapy. Taken together, it is essential to further our understanding of the role of ADAMs within NSCLC. In particular, the function of ADAM12 within the TIM has attracted our attention.
In this study, we obtained The Cancer Genome Atlas (TCGA) data to conduct an in-depth analysis of 12 ADAMs with proteolytic enzyme activity in LUAD tissues and adjacent normal tissues. More specifically, we sought to explore differentially expressed ADAM-encoding genes in LUAD, subsequently focusing on ADAM12. We then assessed its association with clinical pathological characteristics, patient survival, and the TIM, thus providing guidance for LUAD treatment.
Materials and Methods
Data acquisition
Identification of differentially expressed ADAM-encoding genes in TCGA data
In this study, TCGA (
cBioPortal database analysis
The cBioPortal Cancer Genomics Portal (v3.7.21) (
LinkedOmics database analysis
LinkedOmics (
Immune infiltration database (TIMER and TISIDB)
We used the TIMER (
Construction of a prognostic risk score model
Univariate Cox proportional hazard regressions were performed on 42 immune-related genes extracted from the TISIDB database to identify factors related to prognoses (p < 0 .05). Then, the Least Absolute Contraction and Selection Operator (LASSO) Cox regression method was applied to those identified genes (Zhang et al., 2020; #890). A multivariable model with the immune-related genes was constructed. Those genes with nonzero coefficients were screened out to calculate the risk score. A prognostic risk score was generated for each patient with the following formula:
where the value is the relative expression of each selected gene's z-score conversion. The median risk score was considered the cutoff value to divide TCGA LUAD patients into a high-risk group and low-risk group. Univariate and multivariate Cox proportional hazards regression analyses were performed to test whether the immune-related gene-based prognostic model was an independent prognostic factor. A Kaplan-Meier survival curve was constructed. In addition, a receiver operating characteristic (ROC) curve analysis was implemented by using the Survival ROC package to evaluate the prognostic capability of the above model (Heagerty, 2000; #889).
Cell culture and cell transfections
Human bronchial epithelial cell BEAS-2B and the human LUAD cell line A549 were donated by Professor Huang Yi from Fujian Provincial Hospital. Human LUAD cell lines PC-9, H827, and H1299 were purchased from Central South University. All cells were cultured in RPMI 1640 medium (Gibco) supplemented with 10% fetal bovine serum (10% FBS) under a humidified atmosphere of 37°C and 5% CO2. Human ADAM12-targeted small interfering RNAs (siRNAs) were designed and purchased from hippobiotec (Huzhou, China) and Lipofectamine 2000 reagent (Thermo Fisher Scientific) used for small RNA transfection.
The siRNA sequence used in this study was as follows: ADAM12si#1:5′-CCUCAAGGCAACUAAGUAUdTdT-3′; ADAM12si#2:5′-GCGAGAUGAGAGAUGCUAAdTdT-3′; ADAM12si#3:5′-GCAGAUAACCAAGGUUUAAdTdT-3′; and sicontrol:5′-UUCUCCGAACGUGUCACGUdTdT-3′.
Quantitative reverse transcription-polymerase chain reaction
TRIzol reagent (Invitrogen) was used to isolate total cellular RNA. Reverse transcription of ADAM12 was performed using the PrimeScript™ RT reagent Kit (Takara, Japan). Quantitative reverse transcription-polymerase chain reaction (qRT-PCR) was performed using the GoTaq® qPCR Master Mix kit (Promega). ADAM12 expression in NSCLC cell lines was determined by the 2−ΔΔCT method. GAPDH expression was used as an internal control. The primer sequences used in this study were as follows: ADAM12, 5′-ATTCAGCAGTCAGTCTCAGCAC-3′ (forward) and 5′-CTTTTCAGCTTCTTCGCTGG 3′ (reverse); and GAPDH, 5′-GGTGTGAACCATGAGAAGTATGA-3′ (forward) and 5′-GAGTCCTTCCACGATACCAAAG 3′ (reverse).
Western blotting
Cells were lysed with RIPA buffer (Beyotime, Shanghai, China) containing a protease inhibitor cocktail (Thermo Scientific, Waltham, MA) and phenylmethanesulfonyl fluoride. The protein concentration of samples was determined by the BCA method. Thirty to fifty micrograms cell lysate were loaded and separated by sodium dodecyl-sulfate polyacrylamide gel electrophoresis (SDS-PAGE). Protein was then transferred onto polyvinylidene fluoride membranes (Sigma, St. Louis, MO), which were incubated with a specific primary antibody. They were then incubated with specific secondary antibodies. Autoradiography was employed for band density quantification (Quantity One software; Bio-Rad). β-tubulin served as a loading control. The antibodies used were as follows: ADAM12 (sc-293225, Santa Cruz, UA) and β-tubulin (YM3139; Immunoway, China).
Cell Counting Kit 8 Assay
PC-9 cells were digested and resuspended after transfection and were then seeded in 96-well plates at 1 × 105 cells/mL (100 μL/well) and cultured overnight. Subsequently, at 0, 24, 48, 72, and 96 h, each well was added with 10 μL CCK-8 solution (Beyotime) and cells were incubated for an additional 4 h. At last, the absorbance of each well at 450 nm was detected by using a microplate reader.
Wound-healing assay
For the wound-healing experiment, cells were seeded into a 6-well plate (1 × 105 cells/well), and a single layer of cells was scratched with a 10 μL plastic pipette tip to form a uniform wound. The distance between the two edges of migrating cell sheets was photographed under a microscope and quantified. All experiments were repeated at least thrice.
Transwell migration and invasion assays
For the migration assay, 2 × 104 cells in 200 μL serum-free medium were placed in the upper chamber of the Transwell system (8 μm pore size; BD Biosciences, San Jose, CA). For the invasion assay, the chamber was coated with Matrigel (BD Biosciences, Franklin Lakes, NJ), and incubated at 37°C for 4h to solidify Matrigel. Around 5 × 104 cells in 200 μL serum-free medium were transferred to the upper chamber covered with Matrigel, and 600 μL RPMI 1640 medium containing 10% FBS were added to the lower chamber as a chemoattractant. After 24 h of incubation, the remaining cells in the upper chamber were removed, and those in the lower chamber were fixed with formaldehyde and stained with crystal violet (Sigma).
The cells were then imaged under an IX71 inverted microscope (Olympus, Tokyo, Japan) and counted. Then the images were taken at 200 times magnification in five random fields of view, and the number of cells was measured by ImageJ software. All experiments were repeated at least thrice.
Statistical analysis
TCGA data were analyzed using R software (version 4.0.2), and all other experimental results were analyzed using GraphPad Prism 8 statistical software. The unpaired t-test was used for comparison between the two groups, analysis of variance (ANOVA) was used for analyzing differences between multiple groups, and the Kaplan-Meier test was used to compare survival between groups. For experiments performed in triplicate, the data are expressed as the mean ± standard deviation. p < 0.05 indicated that the difference is statistically significant (*p < 0.05, **p < 0.01, ***p < 0.001).
Results
Abnormal expression of ADAMs in LUAD patients
As some ADAMs play significant roles in proteolysis, we explored the expression levels of ADAMs (ADAM8, 9, 10, 12, 15, 17, 19, 20, 21, 28, 30, and 33) with proteolytic enzyme activity in LUAD. Most of these exhibited significant differential expression between LUAD and adjacent normal tissues (Fig. 1A and Table 1). In particular, ADAM12, ADAM28, ADAM8, ADAM15, and ADAM20 expressions were significantly increased in LUAD tissues (p < 0.01 and |log2-fold change (FC)| >1).
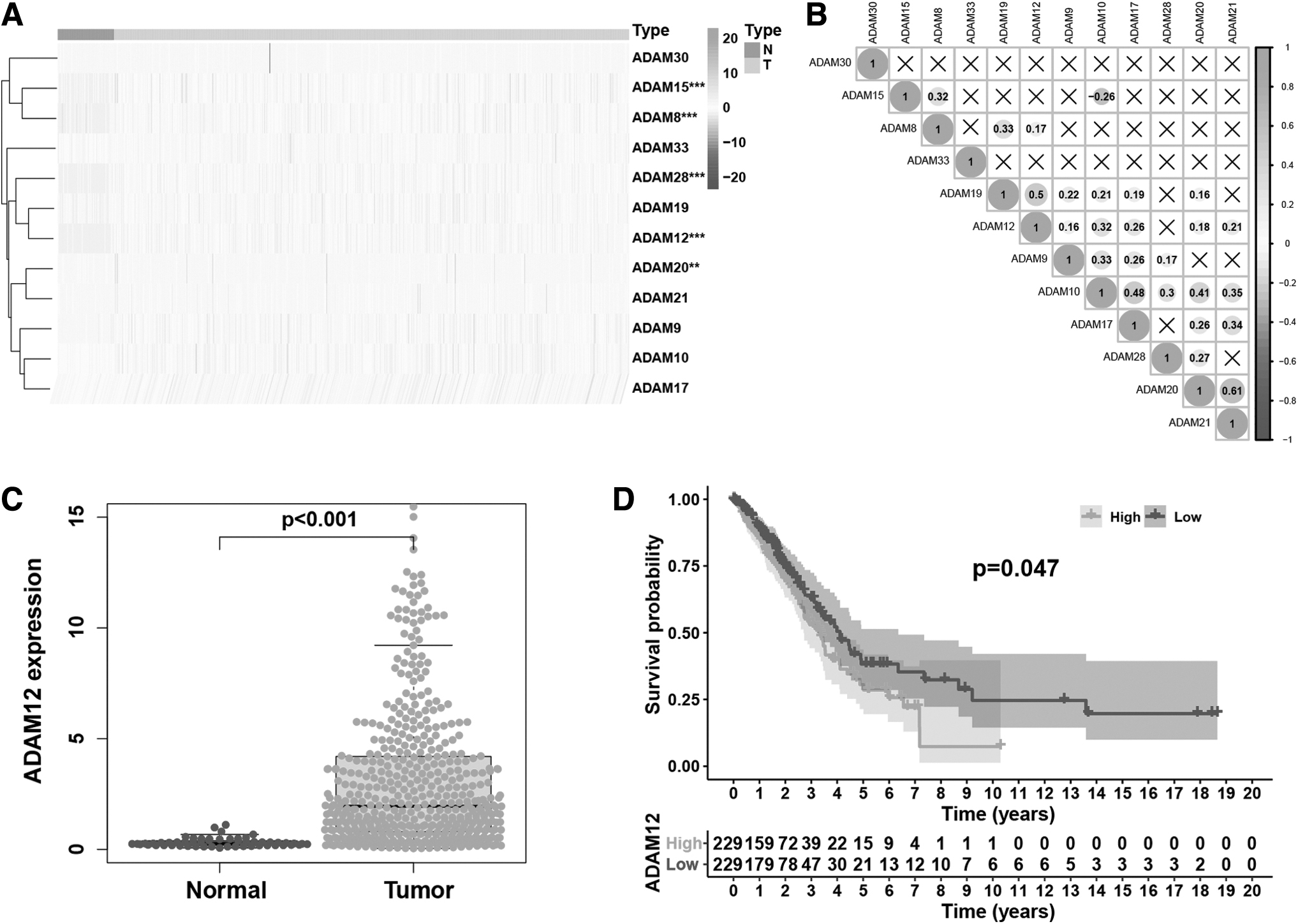
Differences in the expression of ADAMs between LUAD and adjacent normal tissues in TCGA database.
Difference of α-Disintegrin and Metalloproteinase Expression Between Lung Adenocarcinoma Tissues And Adjacent Normal Tissues
FC, fold change; LUAD, lung adenocarcinoma.
We then sought to clarify the correlation among ADAM expressions. Correlation analysis revealed that ADAM19, ADAM12, ADAM9, and ADAM10 exhibited weak to strong correlations with other genes, among which ADAM21 was most strongly correlated with ADAM 20 (R = 0.61), followed by ADAM12 and ADAM19 (R = 0.5) (Fig. 1B). Taken together, correlation analysis indicated a certain association between the expression of different ADAMs.
In the previous literature, studies have found the prognostic biomarkers related to ADAM12 and LUAD TME through bioinformatics (Yue et al., 2019; #703). We further analyzed the expression differences of ADAM12 between LUAD and adjacent normal tissues. ADAM12 expression was significantly increased in LUAD tissues. Furthermore, low ADAM12 expression was significantly associated with better survival outcomes (Fig. 1C, D).
Genetic alteration, expression, and coexpression analyses of ADAM12 in LUAD patients
We analyzed the genetic alterations of ADAM12 in LUAD patients by using the cBioPortal online tool. Patients’ information were obtained from four databases, OncoSG,2020, MSKCC, TCGA PUB, and TSP. ADAM12 mutations, amplifications, and deep deletions were detected, and mutations were more common in LUAD (Fig. 2A, B and Supplementary Fig. S1). To further understand the biological functions of ADAM12, we used the LinkedOmics database to analyze genes co-expressed with ADAM12 in LUAD.

ADAM12 copy number variation and co-expressed genes.
As shown in Figure 2C, 10,626 genes (red dots) exhibited a strong positive correlation with ADAM12, while 9361 genes (green dots) had a strong negative correlation with ADAM12 (false discovery rate [FDR] <0.01, |Spearman's correlation| >0.5). The top 50 positively correlated genes are shown in Figure 2D. Furthermore, we conducted KEGG analysis of these 50 genes, which indicated that they were mainly enriched in protein digestion and absorption, ECM-receptor interactions, and focal adhesions (Fig. 2E), suggesting ADAM12-related genes were closely related to the TME.
The influence of ADAM12 on the TIM
As our findings suggested that ADAM12 is closely related to the TME, we sought to explore whether ADAM12 is associated with tumor immune cell infiltration. We used gene set enrichment analysis (GSEA) to analyze abnormally activated immune-related signaling pathways related to ADAM12 differential expression (FDR <0.05, nonoperative management p-value <0.05) (Fig. 3A). High expression of ADAM12 was closely related to ECM-receptor interactions, B cell receptor signaling pathways, NK cell-mediated cytotoxicity, T cell receptor signaling, and Toll-like receptor signaling.

Influence of ADAM12 on the TIM.
Taken together, ADAM12 was closely related to tumor immune infiltration. TIMER data indicated somatic CNV of ADAM12 were related to the degree of immune cell infiltration. In particular, the chromosomal arm-level gain of ADAM12 was related to a decrease in B cell, CD4+ cell, and dendritic cell infiltration (p < 0.01).
The TIMER database was used to analyze the relationship between four types of somatic CNV (deep deletion, arm-level deletion, arm-level gain, and high amplication, diploid/normal as controls) and the infiltration level of six types of immune cells. Studies showed that compared with the normal group, the infiltration level of B cells, CD4+ T cells, and dendritic cells in the arm-level gain group decreased (p < 0.05). Moreover, the infiltration level of CD8+ T cells and macrophage decreased in the high amplification group (p < 0.001) (Fig. 3B). Furthermore, the expression level of ADAM12 was positively correlated with the degree of infiltration of most immune cell types (p < 0.05) (Fig. 3C).
Based on available literature (Lu et al., 2020; Ma et al., 2020), we selected CD4+ cell, macrophage, neutrophil, and dendritic cell markers to further explore the association of ADAM12 expression with the degree of immune cell infiltration. After tumor purification, cell markers with |r| > 0.4, p < 0.001 were screened. BRIP1 (|r| = 0.446), CCNB1 (|r| = 0.403), EXO1 (|r| = 0.45), KNTC1 (|r| = 0.414), PRC1 (|r| = 0.418), and FAM198B (|r| = 0.423) had moderate to strong correlations with ADAM12 (Fig. 3D and Supplementary Table S1). Five of the six immune cell markers were CD4+ T cell markers, suggestive of an important role for CD4+ T cells in the influence of ADAM12 on immune infiltration.
The influence of ADAM12 and related immune modulatory molecules on prognosis
A variety of immune genes were found to be related to ADAM12 expression. Thus, we used the TISIDB database to further explore the correlation between the expression of immunomodulatory molecules and ADAM12. As shown in Figure 4A and B, most immunostimulatory and immunoinhibitory proteins were positively correlated with the expression of ADAM12 in LUAD and other cancer types. Particularly, in LUAD, TNSF4 (|r | = 0.68, p < 0.001) and PDCD1LG2 (|r| = 0.402, p < 0.001) exhibited the strongest correlation with ADAM12.
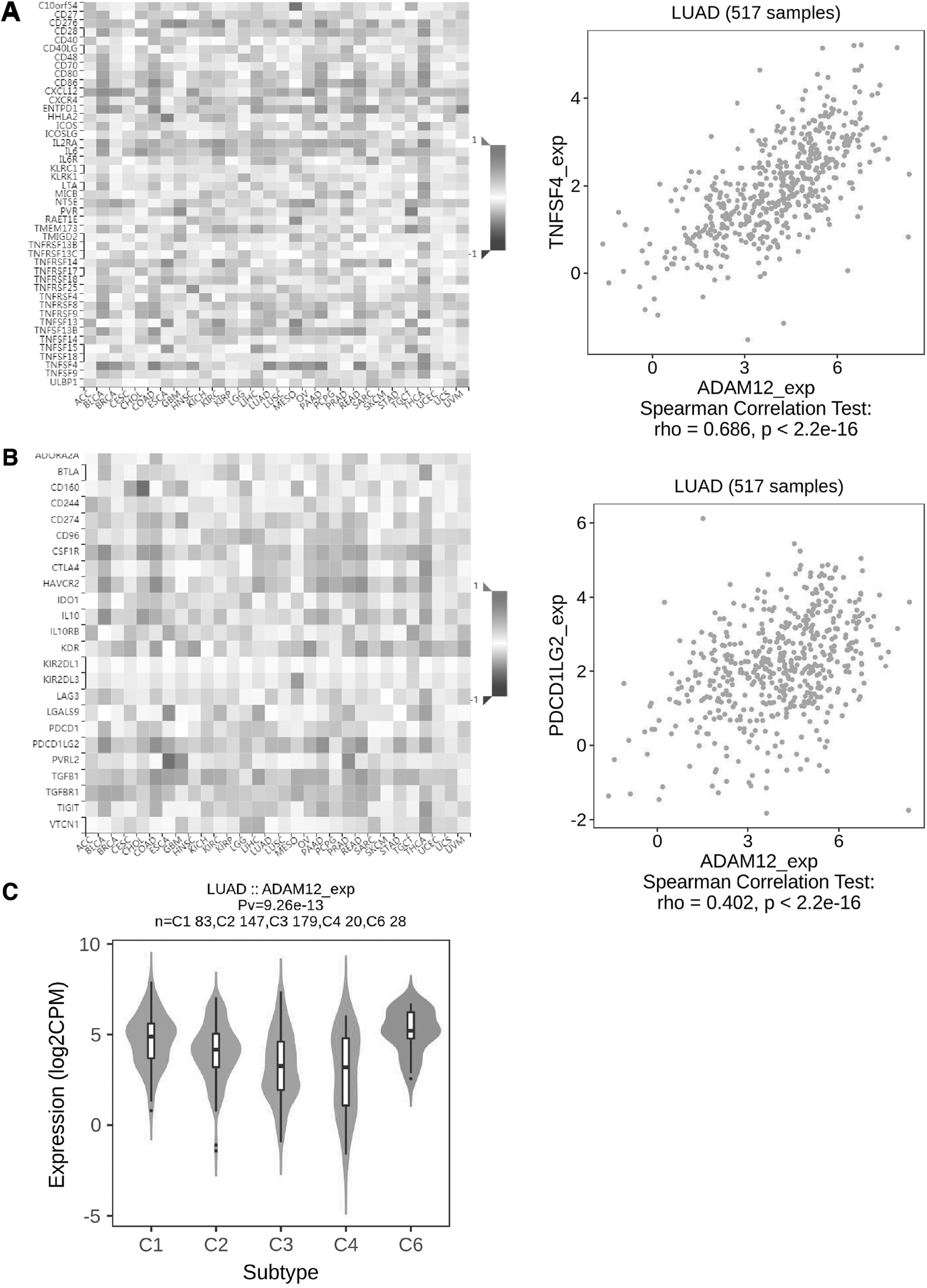
ADAM12-related immunomodulatory molecules.
LUAD was divided into five immune-based categories, including C1 (wound healing); C2 (interferon-gamma dominant); C3 (inflammatory); C4 (lymphocyte depleted); and C6 (transforming growth factor [TGF]-β dominant). ADAM12 expression was highest in the C6 group (TGF-β-dominant), while the C4 group (lymphocyte depleted) exhibited the lowest ADAM12 expression level (Fig. 4C).
To investigate the prognostic significance of ADAM12 and related immunomodulators, we performed a univariate Cox regression analysis and obtained eight prognostic-associated candidate hub genes (Supplementary Table S2). Subsequently, these eight prognostic-associated candidate hub genes were analyzed by LASSO Cox regression to investigate their impact on patient survival time and clinical outcomes; six hub genes were found to be independent predictors in LUAD patients (Fig. 5A, B and Supplementary Table S3).

A prognostic model based on ADAM12 and related immunomodulatory molecules.
Patients were then divided into high-risk and low-risk groups based on the median risk score. The prognosis of patients was indeed more favorable in the low-risk group than in the high-risk group (p < 0.001) (Fig. 5C). To evaluate the accuracy of prognostic risk values, we performed ROC analysis. The AUC for 1-, 3-, and 5-year survival in TCGA data was 0.726, 0.661, and 0.639, respectively (Fig. 5D).
We further analyzed the relationship of high- and low-risk groups with immunoregulatory molecule expression as well as clinicopathological characteristics. Immunostimulatory CD40LG and CD80 expression as well as immunoinhibitory IL-10 expression were lower in the high-risk group. Furthermore, the expression of ADAM12 as well as the immunostimulatory RAET1E and PVR was higher in the high-risk group. The N stage (p < 0.001), T stage (p < 0.001), pathological stage (p < 0.001), and fustat (p < 0.001) were significantly different between the high- and low-risk groups (Fig. 5E). We then employed univariate and multivariate Cox regression to analyze whether the risk score could be used as an independent prognostic factor.
Univariate regression analysis indicated that stage (hazard ratio [HR] = 1.646, 95% confidence interval [CI] = 1.394–1.944, p < 0.001), T (HR = 1.597, 95% CI = 1.283–1.986, p < 0.001), N (HR = 1.769, 95% CI = 1.440–2.174, p < 0.001), and risk score (HR = 5.563, 95% CI = 3.563–8.685), p < 0.001) were significantly correlated with overall survival, while age, gender, and M stage were not (Fig. 5F). Multivariate Cox regression analysis suggested that risk score (HR = 4.440, 95% CI = 2.657–7.418, p < 0.001) could be used as an independent prognostic factor (Fig. 5G). Taken together, these results indicated that ADAM12 and related immunomodulators have prognostic potential in LUAD patients.
ADAM12 knockdown inhibited the proliferation, migration, and invasion of LUAD cells in vitro
We employed Western blotting to detect ADAM12 protein expression in BEAS-2B and LUAD cells. ADAM12 expression was generally high in LUAD cells (Fig. 6A). The sicontrol, siADAM12#1, siADAM12#2, and siADAM12#3 plasmids were, respectively, transfected into PC-9 cells. The results of qRT-PCR and Western blot experiments consistently showed that siADAM12#3 had the highest knockdown efficiency.

Knockdown of ADAM12 inhibited the growth and metastasis of LUAD cells in vitro.
Therefore, siADAM12#3 was chosen as siADAM12 group for subsequent experiments (Fig. 6B and Supplementary Fig. S2). Cell proliferation was assessed by CCK-8 assay, the results of which indicated that compared with the sicontrol group, cell proliferation in the siADAM12 group was noticeably decreased (Fig. 6C). Transwell migration and invasion assays, in addition to a wound-healing assay, indicated that knockdown of ADAM12 inhibited PC-9 cell migration and invasion (Fig. 6D, E). Taken together, the above-presented results suggested that ADAM12 expression was implicated in the metastasis of LUAD.
Discussion
Although currently available multimodal therapy has greatly improved the survival of LUAD patients, treatment outcomes remain unsatisfactory. Thus, the identification of novel therapeutic targets is of great value for LUAD diagnosis and treatment. ADAMs are members of a family of multidomain transmembrane and secretory proteins. Specific ADAMs are upregulated in human cancers and are associated with tumor progression and poor prognosis (Jiao et al., 2018). However, the role of ADAMs in LUAD has rarely been studied.
In this work, we analyzed TCGA data for the expression of 12 ADAMs with proteolytic enzyme activity in LUAD. ADAM12 was highly expressed in LUAD and significantly associated with poor prognosis. We then used the cBioPortal database to explore gene CNV underlying the abnormal expression of ADAM12. Furthermore, we utilized the LinkedOmic database and GSEA to explore the potential biological functions of ADAM12, finding that ADAM12 was closely related to the TME.
The TIMER and TISIDB resources were used to explore the impact of ADAM12 within the TIM. We constructed a prognostic model with ADAM12 and related immunomodulatory molecules. Finally, we verified the biological function of ADAM12 through a number of in vitro assays. To the best of our knowledge, this is the first comprehensive analysis of ADAM12 in LUAD, and the first work to highlight the relationship of ADAM12 with the TIM.
An increasing body of evidence has established the abnormal expression of ADAMs during tumorigenesis (Duffy et al., 2009; Aydin et al., 2015). ADAMs promoted tumor growth and metastasis by regulating the activation of growth factors (such as EGF), cytokines (tumor necrosis factor-α), and integrins (Mochizuki and Okada, 2007).
Among the 12 ADAMs with proteolytic enzyme activity, ADAM9, 10, and 17 have been most extensively investigated within cancer. ADAM9 expression is typically high in cervical, ovarian, prostate, lung, pancreatic, and other cancer types, wherein it is associated with poor prognosis (Lin et al., 2017; Hua et al., 2018; Mohd Isa et al., 2019; Xing et al., 2019; Guo et al., 2020). ADAM10 and ADAM17 are important α-secretases, which can cleave amyloid precursor protein into active fragments and were previously reported as related to poor breast cancer prognosis (Tsang et al., 2018).
Studies have indicated that, compared with that in normal tissue, inflamed tissue, and benign tumor controls, ADAM10 expression was significantly increased in NSCLC cell lines and patient tumors. Furthermore, it was associated with a poor prognosis (Yoneyama et al., 2018). Li et al. (2019) found that ADAM17 expression was higher in gastric cancer patients with lymph node metastasis than in those without. Also, ADAM17 was suggested to promote lymph node metastasis through Notch and Wnt signaling.
In this study, we analyzed ADAMs in 497 cases of LUAD and 54 normal lung tissues from TCGA database. We observed that the expression of most ADAMs was dysregulated in LUAD, with that of ADAM12 being significantly increased.
ADAM12 expression was previously reported to be high in breast, esophageal, pituitary tumor, and choriocarcinoma (Huang et al., 2018; Luo et al., 2018; Wang et al., 2020). In choriocarcinoma, ADAM12 silencing significantly increased the expression of autophagy-related proteins LC3B and ATG5, thereby promoting chorionic cell apoptosis, which indicated that ADAM12 may be a new therapeutic target for choriocarcinoma (Wang et al., 2020).
Besides, in the hypoxia environment of breast cancer cells, HIF-mediated ADAM12 expression, cleaved membrane-bound HB-EGF, forms free HB-EGF, upregulates EGFR-to-FAK signal transduction, and promotes distant metastasis of cells (Wang, 2021; #957). However, there have been limited studies on ADAM12 in NSCLC. Earlier studies had found that the membrane type of ADAM12 (ADAM12-L) could be a prognostic factor in resected pathological (p-) stage I LUAD (Mino, 2009; #958).
Xu et al. explored single-nucleotide polymorphisms in 132 Notch Pathway genes with overall survival (OS) of 1185 NSCLC patients. ADAM12 rs10794069 was prognostic factor for OS in NSCLC patients in a risk-genotype dose–response manner. These results showed that ADAM12 may have biological significance and therapeutic application in NSCLC (Xu, 2016; #959). Through bioinformatics analyses, Yue et al. (2019) previously reported ADAM12, BTK, and ERG were mainly related to the activation of pathways involving focal adhesion and immune regulation within the TME. However, this was the only report on the biological function of ADAM12 in LUAD. We carried out in vitro experiments and observed, for the first time, that ADAM12 knockdown reduced the proliferation, migration, and invasion of LUAD cells.
To further understand the biological role of ADAM12 in LUAD, 19,987 ADAM12 co-expressed genes were identified using the LinkedOmics database. KEGG enrichment analysis revealed that genes positively correlated with ADAM12 were enriched in ECM-receptor interactions and adhesion-related pathways, further confirming the association of ADAM12 with the TME. Qi et al. (2020) previously suggested that the ADAM family proteases could be used as potential biomarkers of immune infiltration in pancreatic cancer. In this study, GSEA revealed that ADAM12 was related to B cell receptor, ECM receptor, T cell receptor, and TOLL-like receptor signaling pathways, as well as NK cell-mediated cytotoxicity, thus confirming the implication of ADAM12 within the TIM.
CNV describes the phenomenon of a DNA fragment with a difference in gene copy number, potentially leading changes in gene expression, which may in turn promote tumorigenesis. In this study, we found that ADAM12 mainly exhibited missense mutations, followed by deep deletions and amplifications. Interestingly, ADAM12 mRNA expression was highest in the cases of deep deletion. Furthermore, arm-level gain was correlated with the infiltration of B cells, CD4+ T cells, and dendritic cells. In addition, high amplification was associated with the tumor infiltration of CD8+ T cells and macrophages. In contrast, the degree of neutrophil infiltration was not affected by ADAM12 copy number alterations.
Immune markers are relevant for tumor prognosis and immunotherapy response prediction (Kim et al., 2016; Goodman et al., 2017). Zhang et al. (2020) reported that deleterious NOTCH mutations may lead to the activation of immune-related genes and were associated with an improved tumor immunotherapy response, suggestive of their potential as predictive markers in NSCLC. Furthermore, ZFHX3 mutations were also associated with enhanced immunogenicity and activation of antitumor immunity (Zhang et al., 2021), supporting their utility as immunotherapy predictive markers.
To provide a theoretical basis for the relationship between ADAM12 and tumor immune infiltration, we determined the association between ADAM12 expression and various classic immune biomarkers. ADAM12 was strongly correlated with CD4+ cell markers BRIP1, CCNB1, EXO1, KNTC1, and PRC1.
Therefore, ADAM12 may be utilized as a biomarker of immune infiltration based on its putative relationship with these immune markers, thus predicting LUAD prognosis. Through the TISIDB database, we found that the immunostimulatory TNFSF4 and ADAM12 had a significant positive correlation, suggestive of ADAM12's implication in LUAD immunotherapy. We constructed a prognostic model based on ADAM12 and five related immunomodulatory molecule-encoding genes. The model accurately predicted patient prognosis. Based on the above results, we speculated that ADAM12 expression may affect the prognosis of LUAD patients through its effects within the TIM.
Nevertheless, our study had certain limitations. First, we screened immune cell markers from published literatures and analyzed the correlation between ADAM12 and immune cell markers using TIMER database. However, in fact, these genes are not unique to immune cells. Second, no in vitro or in vivo experiments were conducted to confirm the association of ADAM12 and immune infiltrations. Thus, further investigation is warranted to verify this relationship, allowing for a better understanding of ADAM12 in the context of tumor treatment.
Conclusion
In summary, we found that ADAM12 may act as an oncogene, affecting LUAD prognosis. Furthermore, bioinformatics analysis suggested that ADAM12 and its co-expressed genes regulated ECM-receptor interactions and focal adhesions in LUAD. ADAM12 CNV was related to the level of immune cell infiltration, suggesting that it may play a role in regulating the TIM, which provides a new direction for future studies focused on LUAD biomarkers.
Footnotes
Authors’ Contributions
Y.Q.X. and J.F.P. conceived of and designed the study strategy. J.F.P. and Z.D.H. collected and analyzed the data. J.F.P. and Y.Q.X. wrote the article. All authors read and approved the final article.
Acknowledgment
The authors thank the TCGA, LinkedOmics, TIMER, TISIDB, and cBioportal for their open access databases.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by Youth Program of National Natural Science Foundation of China (Grant no.: 82102991) and Natural Science Foundation of Fujian Province (Grant no.: 2021J05084).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.