
Abstract
Scaffold materials used for bone defect repair are often limited by osteogenic efficacy. Moreover, microRNAs (miRNAs) are involved in regulating the expression of osteogenic-related genes. In previous studies, we verified the enhancement of osteogenesis using a grooved porous hydroxyapatite scaffold (HAG). In the present study, we analyzed the contribution of HAG to the osteogenic differentiation of human placenta-derived mesenchymal stem cells (hPMSCs) from the perspective of miRNA differential expression. Furthermore, results showed that miRNAs were differentially expressed in the osteogenic differentiation of hPMSCs cocultured with HAG. In detail, 16 miRNAs were significantly upregulated and 29 miRNAs were downregulated with HAG. In addition, bioinformatics analyses showed that the differentially expressed miRNAs were enriched in a variety of biological processes, including signal transduction, cell metabolism, cell junctions, cell development and differentiation, and that they were associated with osteogenic differentiation through axon guidance, mitogen-activated protein kinase, and the transforming growth factor beta signaling pathway.
Furthermore, multiple potential target genes of these miRNAs were closely related to osteogenic differentiation. Importantly, overexpression of miR-146a-5p (an upregulated miRNA) promoted the osteogenic differentiation of hPMSCs, and miR-145-5p overexpression (a downregulated miRNA) inhibited the osteogenic differentiation of hPMSCs.
Background
Bone is a mineralized mesenchymal tissue that resizes mechanical forces and regulates mineral homeostasis and energy metabolism (Lian et al., 2012). However, bone defects due to trauma, disease, and other causes impair people's quality of life and cause a significant social and economic burden. Although bone has a certain regeneration ability, serious bone injuries, such as large bone defects, still need to be treated by bone grafting (Yang et al., 2013; Montazerolghaem et al., 2016). Autografts are the most ideal method to avoid the occurrence of graft immune response, but they are limited by multiple surgeries and donor-site injury.
Therefore, bone tissue engineering, a scaffold material loaded with seed cells and growth factors, has become a more promising method for clinical therapy. As a key part of bone tissue engineering, the development of biomaterials with different properties is particularly important. Among them, hydroxyapatite (HA) is the most widely used scaffold material.
However, when HA is used in bone repair, the osteogenic response is not ideal because the high density of micropores on the surface allows the newly formed artificial bone to move freely (Mondal et al., 2016; Perez and Mestres, 2016). To address this problem, we previously developed porous hydroxyapatite scaffolds with a grooved structure (HAG scaffolds) and demonstrated that HAG scaffolds promote bone formation and neovascularization better than HA scaffolds both in vitro and in vivo (heterotopic and orthotopic implantation) (Ren et al., 2018).
Tissue engineering strategies are still limited by the inaccessibility of adequate osteogenic cells (Kolambkar et al., 2010). Mesenchymal stem cells (MSCs) are a group of cells with multidirectional differentiation potential that can differentiate into a variety of cell types, including osteoblasts and adipocytes, in vitro (Ulrich et al., 2015). MSCs are used in hematopoietic recovery, regenerative medicine, and tissue engineering (Chen et al., 2006; Brooke et al., 2007). Bone marrow-derived mesenchymal stem cells (BMSCs) are the most commonly used cell type in tissue engineering. However, bone marrow collection remains an invasive process; it is difficult to obtain large quantities of bone marrow, and bone marrow decreases significantly with donor age (Owen and Friedenstein, 1988; Fehrer and Lepperdinger, 2005).
In contrast, the placenta, a transient organ that maintains fetal tolerance, constitutes a rich MSC library (Parolini et al., 2010; Maxson et al., 2012). More importantly, MSCs are easily isolated from the placenta without invasive procedures, thereby avoiding ethical concerns (Brooke et al., 2009; Patel et al., 2014). Human placenta-derived mesenchymal stem cells (hPMSCs) express a variety of cell surface antigens, including CD29, CD90, CD140b, CD44, CD146, and CD166, but they lack the expression of hematopoietic- or endothelial-specific antigens, such as CD14, CD34, CD31, CD45, or CD133 (Pilz et al., 2011; Tormin et al., 2011; Komaki et al., 2017).
MicroRNAs (miRNAs) are endogenous small noncoding ∼22-nt RNAs that regulate a series of physiological and pathological processes, including bone regeneration and metabolism, by targeting mRNA sequences of related genes (Liu and Paroo, 2010; Ha and Kim, 2014; Oliviero et al., 2019; Giordano et al., 2020; Gargano et al., 2021). Proliferation, matrix maturation, and mineralization are three successive stages in osteogenesis, which are achieved through temporally and spatially expressed genes and fine regulation by a host of miRNAs (Lian et al., 2004).
miRNA array profiling has been widely used to identify differential miRNA expression during osteogenic differentiation. Inose et al. (2009) revealed that 36% of miRNA expression is downregulated and that 3% of miRNA expression is upregulated during bone morphogenetic protein 2 (BMP2)-induced osteogenic differentiation of mouse C2C12 mesenchymal progenitor cells. Moreover, 33 miRNAs are differentially expressed in differentiated and undifferentiated hBMSCs (Eskildsen et al., 2011). Additionally, during the mineralization stage of murine calvaria-derived preosteoblasts (MC3T3 cells), 58 miRNAs are upregulated, and 10 miRNAs are downregulated (Li et al., 2009).
In detail, overexpression of miR-124 inhibits osteogenic differentiation of BMSCs by targeting SP7 (Tang et al., 2019), and miR-23a overexpression inhibits osteogenic differentiation of BMSCs (Zhuang et al., 2019). Moreover, Tian et al. (2015) revealed that miR-23a inhibits osteogenic differentiation of BMSCs by targeting RUNX family transcription factor 2 (Runx2). Moreover, miR-21 has been identified to promote the migration and osteogenic differentiation of BMSCs in vitro (Yang et al., 2019). Yan et al. (2018) found that miR-92a binds to the 3′-untranslated region (UTR) of SMAD family member 6 (Smad6) to inhibit the degradation of Runx2 degradation and promote osteogenic differentiation of BMSCs. As mentioned above, different miRNAs are involved in the complex process of osteogenic differentiation. However, the miRNA profile related to hPMSC osteogenic differentiation is still unclear.
In the present study, we cocultured hPMSCs with HAG scaffolds and evaluated the differential expression of miRNAs in the process of cell osteogenic differentiation induced by HAG scaffolds in hPMSCs using microarray assays. We further explored the biological functions that these differential miRNAs may be associated with osteogenesis through bioinformatics analysis and revealed that they were closely related to osteogenic differentiation-related signaling pathways, including axon guidance, mitogen-activated protein kinase (MAPK), and transforming growth factor beta (TGF-β) signaling pathways. Our study provides a basis for further studies regarding biomaterial osteogenic regulatory functions, especially the involvement of miRNAs.
Materials and Methods
Cell culture and transfection
The immortalized hPMSCs used in this study originated from the State Key Laboratory of Biological Therapy at Sichuan University (provided by the National Experimental Cell Resource Sharing Platform,
For osteogenic differentiation, hPMSCs were cultured with conditioned medium (DMEM with 10% FBS, 20 mM β-glycerophosphate, 50 g/mL vitamin C, and 10 M dexamethasone; Sigma-Aldrich). After 7 days of induction culture, alkaline phosphatase (ALP) activity was detected according to the manufacturer's instructions (A059-2; Nanjing Jiancheng Bioengineering Institute, China). Alizarin Red staining (ARS) was performed to detect calcium deposits after 21 days of induction culture (G3280; Solarbio Life Sciences, China).
For cell transfection, when cells reached 80% confluency, GFP-NC (100 pM) and miRNA mimics (100 pM; miR146a-5p and miR145-5p; synthesized by GenePharma) were transfected into cells using the Lipofectamine™ 2000 (Thermo Fisher Scientific) transfection reagent according to the manufacturer's protocol. Nuclei were then stained with 4′,6-diamidino-2-phenylindole (DAPI; Sigma-Aldrich) 48 h after transfection. Fluorescent images of GFP and DAPI were captured by a ZEISS AXIO Observer A1. Twenty-four hours after transfection, the medium was replaced with osteogenic induction medium. After 7 days of osteogenic induction culture, ALP staining was performed using the ALP Staining Kit (Wako, Japan) according to the manufacturer's protocol, and the mRNA expression of osteogenic markers was detected. ARS was performed after 14 days of induction culture.
Preparation of scaffolds and hPMSCs cocultured with HAG
The HAG scaffold was produced and cleaned according to previously reported methods (Ren et al., 2018). The surface micromorphology of HAG was confirmed by scanning electron microscopy (ZEISS Gemini 300). The scaffolds were incubated overnight in DMEM before the cell experiment at 37°C. For coculture with hPMSCs, 20 μL of cell suspension (4 × 106/mL) was directly added dropwise onto the surface of the HAG scaffold and incubated for 1 h, and culture medium was then added. The osteogenic induction medium was replaced the next day.
Quantitative real-time PCR
Total RNA was extracted by TRIzol™ reagent and reverse transcribed to cDNA using M-MLV (Thermo Fisher Scientific). Quantitative fluorescence detection was performed using 2X ChamQ Universal SYBR Master Mix (Vazyme, China) and an ABI 7500. The primer sequences are shown in Tables 1 and 2 (designed and synthesized by Tsingke, China).
Primers for microRNA Reverse Transcription
Primers for Quantitative Polymerase Chain Reaction Analysis
ALP, alkaline phosphatase; BMP2, bone morphogenetic protein 2; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; OSX, osterix; RUNX2, RUNX family transcription factor 2.
Western blotting
Cells were collected, lysed in cell lysis buffer, and vortexed every 5 min for 25 min on ice. The lysates were then centrifuged at 12,000 g and 4°C for 10 min. Proteins were isolated by 10% SDS–PAGE (sodium dodecyl sulfate–polyacrylamide gel electrophoresis) and transferred to polyvinylidene fluoride membranes (Millipore). After incubation with glyceraldehyde-3-phosphate dehydrogenase, ALP and osterix (OSX) (1:1000; Huabio, China) primary antibodies overnight, the membranes were washed five times with TBST (2‰ Tween). The membranes were then incubated with horseradish peroxidase-conjugated secondary antibody for 1 h. Finally, a Super ECL Kit (Biosharp, China) was used to detect the protein bands using a Tanon-5200.
Microarray assays
Total RNA was extracted using TRIzol reagent and used to construct a miRNA library by NEBNext® Multiplex Small RNA Library Prep Set for Illumina (NEB). The library quality was determined by an Agilent 2100 Bioanalyzer (KangChen Biotech, Shanghai, China), and sequencing was performed using an Illumina NextSeq 500 Sequencer. Bowtie was used to map small RNA tags to the reference sequences (Langmead et al., 2009), and miRDeep2 software was used to search for potential known miRNAs (Friedländer et al., 2012).
Bioinformatics analysis
Edge R was used to analyze the differential expression of miRNAs between groups. Based on the TargetScan7.1 and mirdbV6 databases, target genes were screened for the top 10 miRNAs that were significantly upregulated and downregulated. Functional enrichment analysis of target genes was then performed by Gene Ontology, including molecular function (MF), cellular component (CC), and biological process (BP) (Mao et al., 2005). The miRNA–mRNA networks were constructed using Cytoscape.
Statistical analysis
After sequencing, miRNAs with expression differences of two times or more were selected for subsequent bioinformatics analysis. Independent repeated experiments were used to perform and analyze related results (n = 6, mean ± standard deviation). Student's t-test was conducted to compare means between two groups. p-Values <0.05 were considered significant.
Results
Osteogenic differentiation of hPMSCs is promoted by HAG
HA is widely used to promote bone formation. In a previous study based on porous HA scaffolds, we prepared a new scaffold material, HAG, by integrating microchannel structures, and we found that HAG promotes the osteogenic response of hPMSCs compared with HA (Ren et al., 2018). Consistently, in the present study, we confirmed the osteogenic effect of HAG in hPMSCs (Fig. 1). The properties of hPMSCs were detected by flow cytometry. The CD44- and CD90-positive surface markers were highly expressed (97.08%) in the cells, but CD45 (0.10%) was barely expressed in the cells (Fig. 1A).

HAG scaffolds increase the osteogenic differentiation of hPMSCs.
The porous structure (arrow shown, left) and 25–30 μm groove structure (arrow shown, right) on the HAG scaffold surface are shown in Figure 1B. After osteogenesis was induced for 7 and 14 days, the mRNA expression of ALP, BMP2, and OCN was quantified by quantitative real-time PCR (qRT-PCR), and the protein levels of ALP and OSX were detected at 7 days (Fig. 1C, D). ARS was used to detect mineralized nodules 3 weeks after induction (Fig. 1E). The results verified the positive effect of HAG scaffold on osteogenic differentiation in hPMSCs.
The miRNA expression in HAG-hPMSCs and control hPMSCs varies
The involvement of miRNAs in the regulation of bone formation has been reported in several studies (Li et al., 2009; Tian et al., 2015; Tang et al., 2019; Zhuang et al., 2019). However, the roles of miRNAs in bone formation mediated by HAG scaffolds remain unclear, especially in hPMSCs.
Thus, to elucidate how HAG scaffolds promote osteogenic differentiation of hPMSCs, a microarray assay was used to analyze the miRNA expression profiles between HAG-hPMSCs and hPMSCs. We observed 45 mature miRNAs that were significantly differentially expressed in the HAG-hPMSC group (FC ≥2, Fig. 2A). Red represents highly expressed genes, and green represents downregulated genes. Darker colors indicate more distinct expression differences. Compared with differentiated hPMSCs, the expression of 16 miRNAs in HAG-hPMSCs was upregulated, and the expression of 29 miRNAs was downregulated as shown in the volcano plot (FC ≥2, Fig. 2B).
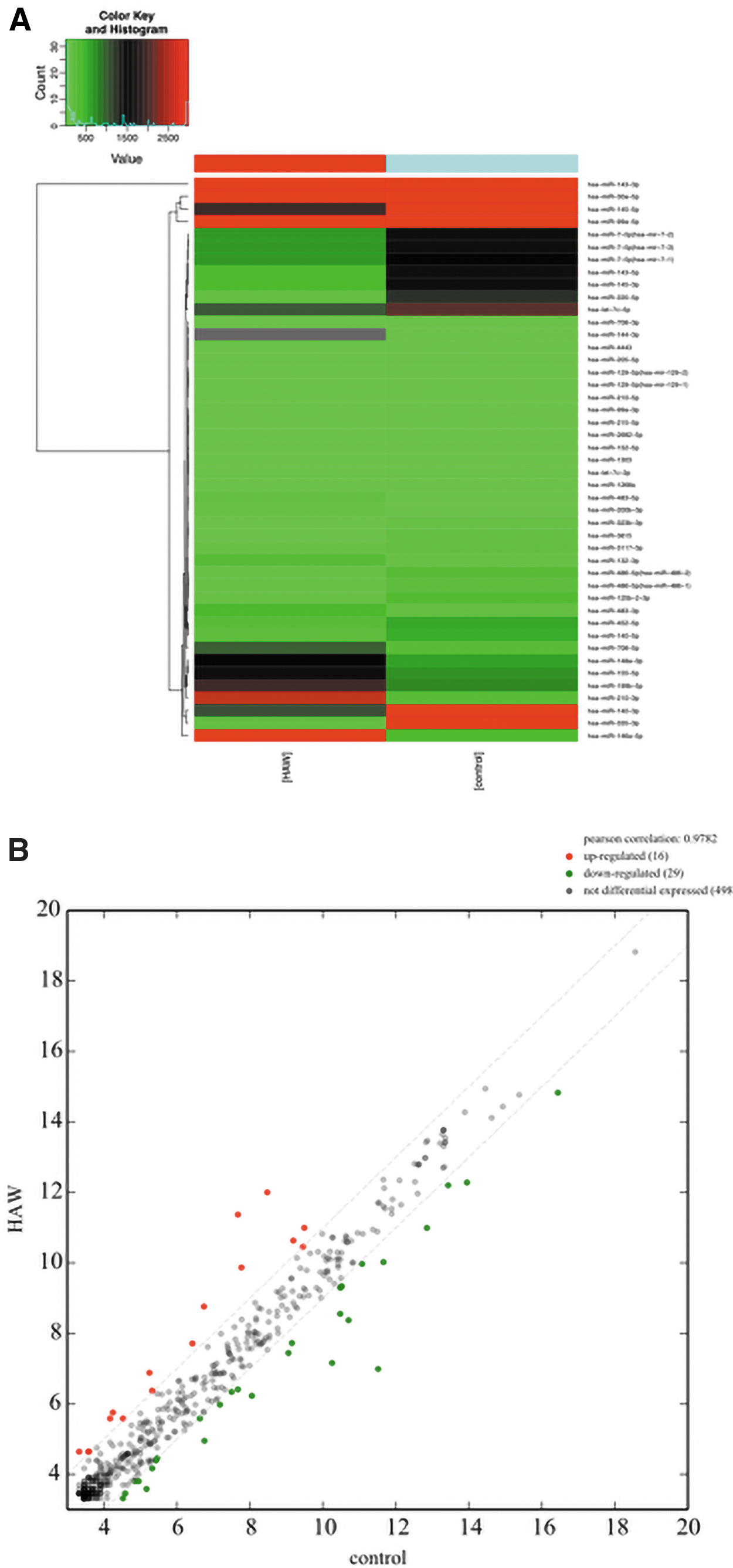
miRNA differential expression profiles in HAG-hPMSCs and hPMSCs.
Pathway and functional analysis of differential miRNAs
In the present study, Gene Ontology enrichment analysis was performed based on differentially expressed miRNAs, including BP, CC, and MF, by cluster profiler in R software to explore the biological functions that may be regulated by differential miRNAs. The statistically significant results (p < 0.05) showed that these miRNAs participated in the regulation of cell metabolism (p = 2.58E-12), cell junctions (p = 6.45E-08), cell development (p = 1.82E-11), cell differentiation (p = 4.04E-07), and signal transduction (p = 8.12E-12) (Fig. 3A). Importantly, the process of osteogenic differentiation is closely related to these functions.
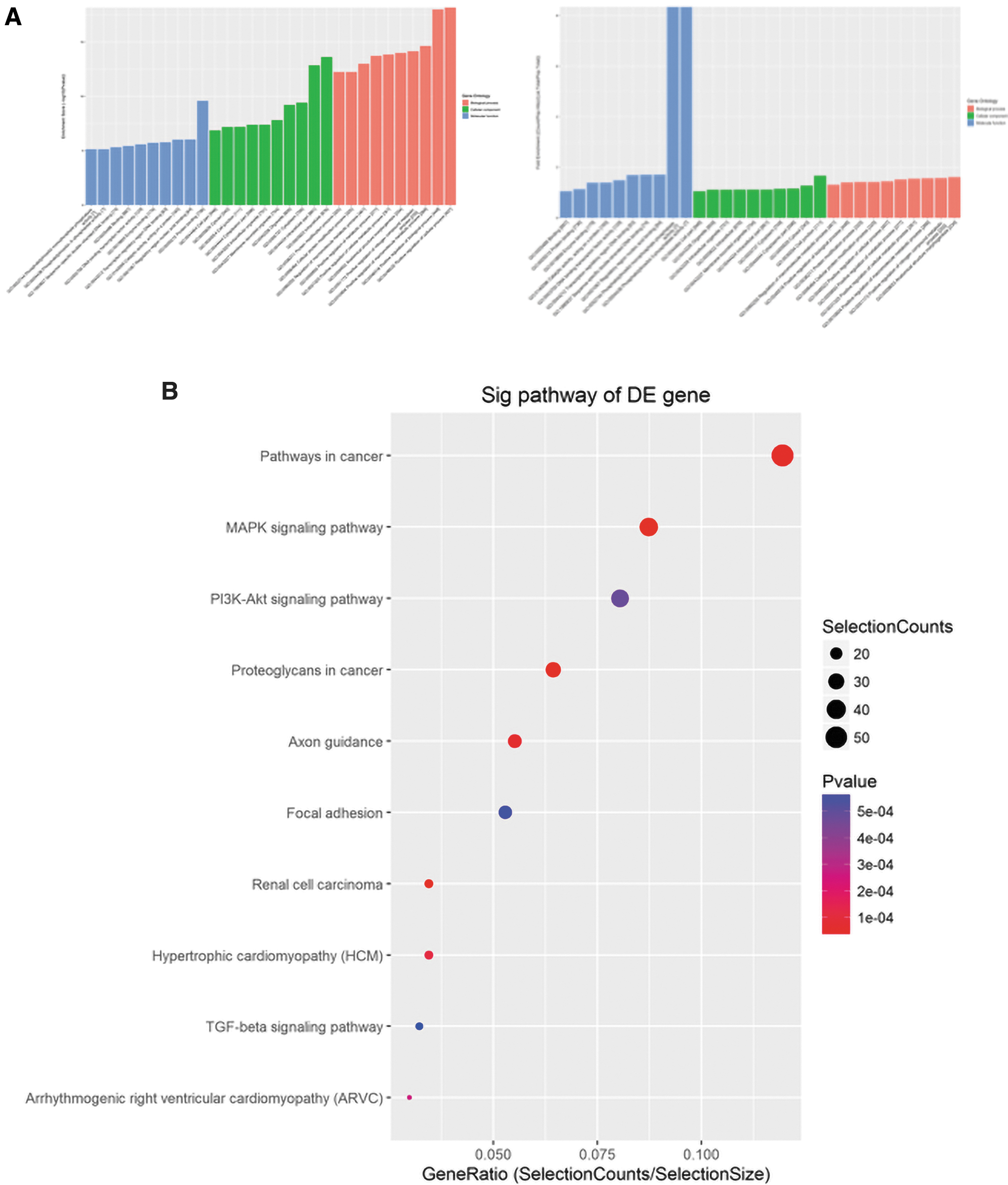
Pathway and GO analysis.
Furthermore, Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis was used to understand the signaling pathways, in which these differentially expressed miRNAs are involved in regulation. The results indicated that genes in multiple signaling pathways, including the axon guidance (p = 5.02E-05), MAPK (p = 6.65E-07), and TGF-β signaling pathways (p = 5.7E-04), were highly associated with osteogenic differentiation and were potential target genes for these differentially expressed miRNAs (Fig. 3B). These results suggested that HAG may achieve its osteogenic effect by promoting the differential expression of these miRNAs.
qRT-PCR validation of miRNA expression and miRNA–mRNA network analysis
According to the microarray analyses, the six miRNAs (miR-210-3p, miR-146a-5p, miR-483-5p, miR-3615, miR-125b-2-3p, and miR-145-5p) with the most significant expression differences were verified by qPCR. Consistently, the expression of miR-210-3p, miR-146a-5p, and miR-483-5p was significantly increased, whereas the expression of miR-3615, miR-125b-2-3p, and miR-145-5p was significantly decreased (FC >2, p < 0.05) (Fig. 4A). miRNAs act as post-transcriptional regulators and regulate the expression of target genes, mainly through binding with the 3′-UTRs of target genes (Liu and Paroo, 2010; Ha and Kim, 2014). Thus, TargetScan7.1 and mirdbV6 were used to predict the target genes of the abovementioned six miRNAs, and miRNA–mRNA networks were constructed by Cytoscape (Fig. 4B).

Verification of the expression of selected miRNAs and construction of the miRNA–mRNA interaction network.
As shown in Figure 4B, the miRNAs targeted hundreds of mRNAs, but the mRNA of BRCC3 was recognized by different miRNAs. More importantly, many target mRNAs are involved in osteogenic differentiation-related pathways, such as Kruppel-like factor 7 (KLF7), SRY-box transcription factor 9 (SOX9), and brain-derived neurotrophic factor (Veldman et al., 2007; Iezaki et al., 2018).
Overexpression of miR-146a-5p promotes the osteogenic differentiation of hPMSCs
To verify the effect of differentially expressed miRNAs on osteoblastic differentiation, we transfected miR-146a-5p mimics into hPMSCs. As shown in Figure 5A and B, the mimics were transfected into cells successfully. Furthermore, we observed that the expression of osteogenic genes was increased when miR-146a-5p was overexpressed (Fig. 5C). Following osteogenic differentiation, the hPMSCs transfected with miR-146a-5p mimics showed a higher intensity of ALP staining and formed a larger number of calcified nodules compared with the control hPMSCs (Fig. 5D, E).
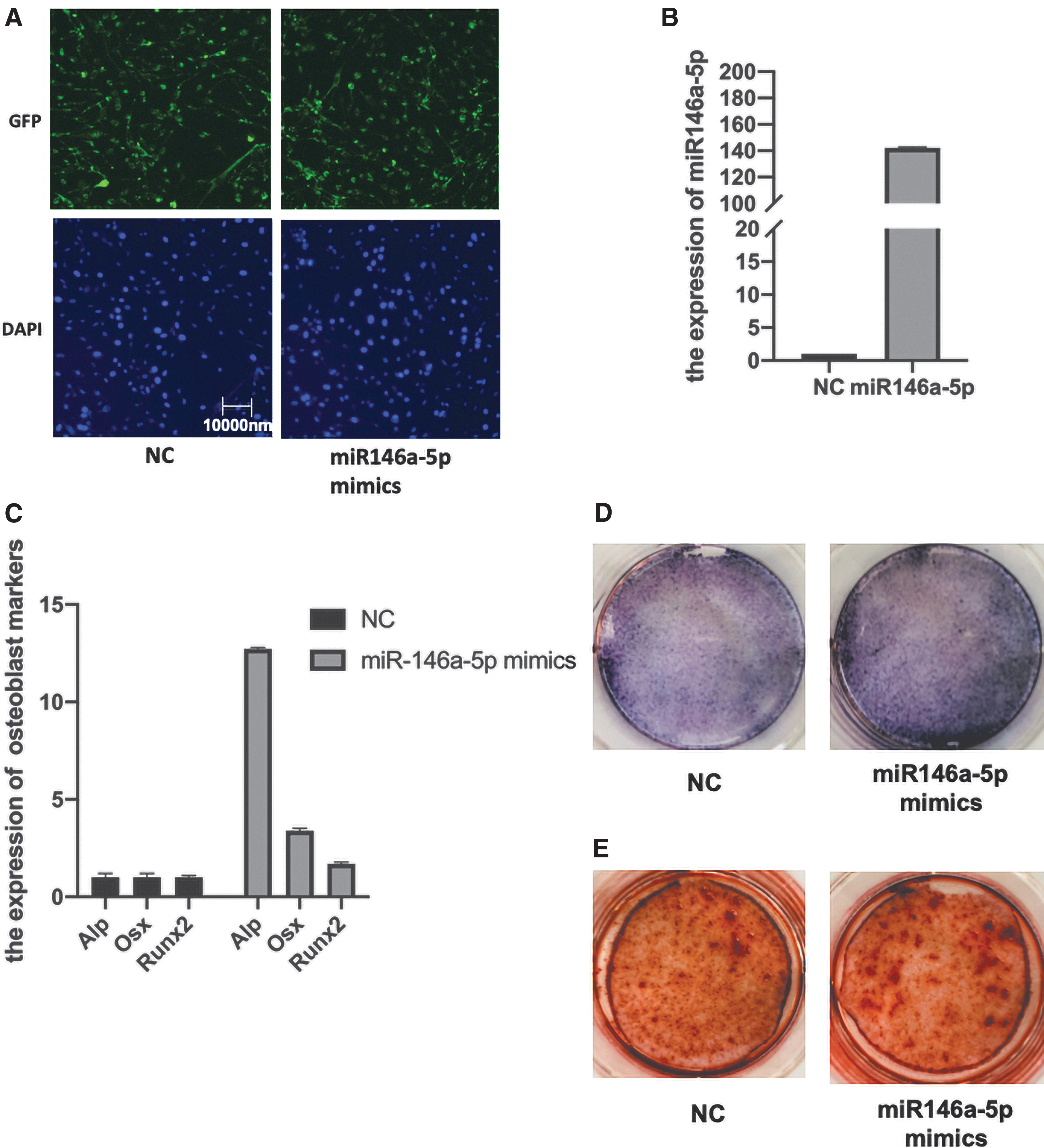
Overexpression of miR-146a-5p promotes the osteogenic differentiation of hPMSCs.
Discussion and Conclusion
As the key component of bone tissue engineering, scaffolds serve as a critical function in supporting cell adhesion, growth, pro-osteogenesis, and so on. Among them, HA is widely used in bone repair due to its similar chemical composition to natural bone and superior osteogenic capacity in vivo (Lee et al., 2000; Cao and Dai, 2002; Ghomi et al., 2011).
More importantly, the osteogenic capacity of scaffolds is correlated to its surface structure (Davison et al., 2015; Zhu et al., 2017). Murphy et al. (2010) reported that scaffolds with well-connected macroporous structures (300–500 μm) were more favorable for nutrient and metabolic waste transport, vascular growth, and direct osteogenesis. Habibovic et al. (2006) found that biphasic calcium phosphate composite ceramics with 17% microporosity promoted bone formation but 3% did not. In our previous study, porous HA scaffolds with a grooved structure (HAG scaffolds, width: 25–30 μm) on the surface were prepared and showed good protein adsorption enhancing direct growth and osteogenic gene expression (Ren et al., 2018). However, the mechanism by which HAG promotes osteogenic differentiation remains unclear.
An increasing number of studies have revealed that miRNAs are involved in regulating cell attachment and proliferation (Liu and Paroo, 2010; Ha and Kim, 2014). Consistently, several studies have revealed that miRNAs are involved in the osteogenic process of biomaterials. Pizzicannella et al. (2018) reported that miRNA-210 mediates vascular endothelial growth factor A (VEGF-A) upregulation in human periodontal ligament stem cells cultured on 3D HA ceramic scaffold. Izadpanahi et al. (2018) found that nanotopographical cues influence the osteogenic differentiation process of hMSCs through the modulation of lncRNAs and miR-125b as negative regulators of osteogenesis. In addition, miR-23a is significantly downregulated in the osteogenic differentiation process of BMSCs on nanostructured titanium surfaces, and Zhuang et al. (2019) revealed that p53 mediates the miR-23a/CXCL12 pathway in the osteogenic differentiation of BMSCs on nanostructured titanium surfaces. These studies provide evidence for the potential mechanism of HAG scaffold promoting the differential expression of miRNAs.
Therefore, we explored the osteogenic mechanisms of HAG scaffolds, particularly the miRNAs involved. In the present study, we detected differential expression of miRNAs in the HAG-hPMSC group compared with cells without HAG. Among them, the expression of miR-210-3p, miR-146a-5p, miR-483-5p, miR-3615, miR-125b-2-3p, and miR-145-5p was the most significant.
There are several studies on the role of these miRNAs in osteogenesis. Cheung et al. (2014) reported that overexpression of miRNA146a increases the expression of RUNX2 by downregulating SMAD2 and SMAD4. In addition, two material associations suggest that the osteogenic effect is closely related to miRNA146 expression. Portland cement, a bioaggregate, has been shown to enhance osteogenesis through miR-146a upregulation. Moreover, exogenous miR-146a increases the expression of osteogenic genes, including ALP and bone morphogenic protein (Wang et al., 2018). Zhou et al. (2016) revealed that the simplest form of silicate, Si(OH)4, stimulates human MSC osteoblastic differentiation by upregulating the expression of miR146a.
Further analysis has revealed that Si(OH)4 upregulates miR-146a to antagonize the activation of nuclear factor kappa B (NF-κB). Consistently, we found that miR-146a-5p was highly expressed in hPMSCs cocultured with the HAG scaffold and that miR-146a-5p mimics promoted osteogenic differentiation (Figs. 4A and 5). VEGF has been shown to facilitate cell migration into engineered bone, thereby triggering tissue regeneration (Kempen et al., 2009). Pizzicannella et al. (2018) reported that miRNA-210 mediates VEGF upregulation in human periodontal ligament stem cells cultured on 3D HA ceramic scaffold (Izadpanahi et al., 2018). Asgharzadeh et al. (2018) found that the osteogenic differentiation of MSCs is promoted by miR210 upregulation.
A previous study on the osteogenic differentiation of bone marrow MSCs induced by the Chinese medicine active ingredient CN has shown that miR-483 is one of the key miRNAs involved in the regulation (Hou et al., 2018). Moreover, miR-125b has been found to attenuate the osteogenic differentiation of periodontal ligament cells by targeting NKIRAS2 (NFKB inhibitor interacting Ras-like 2) and enhancing NF-κB signaling (Xue et al., 2018).
Additionally, Wang et al. (2017) reported that Cbfβ (core-binding factor subunit beta) and BMPR1b (bone morphogenetic protein receptor type 1B) are the downstream targets of miR-125b in inhibiting osteoblastic differentiation of MSCs (Huang et al., 2014a). Furthermore, in the present study, miR-125b-2-3p was downregulated in hPMSCs cocultured with the HAG scaffold, and miR-125b-2-3p mimics decreased osteogenic differentiation (Fig. 4A). For miRNA-145, Jin et al. (2013, 2020) reported that Sp7 (also named osterix), semaphorin 3A, and FoxO1 (forkhead box O1) are targets that mediate the inhibition of osteogenic differentiation. However, no studies related to osteogenesis of miR-3615 have been reported at present.
The integration of pathway analysis, functional analysis, and miRNA–mRNA cross-network analysis showed that the miRNAs differentially expressed in the HAG scaffold group were highly correlated with the axon guidance, MAPK, and TGF-β signaling pathways, which have been reported to be involved in the regulation of osteogenic differentiation (Lee et al., 2002; Koh et al., 2006; Tamagnone and Giordano, 2006; Kanno et al., 2007; Ge et al., 2009; Huang et al., 2014b; Hao et al., 2018).
For example, KLF7 was predicted to be a highly likely target of miR-146a-5p, and miR-146a-5p was upregulated in the hPMSC-HAG group (Fig. 4A, B). KLF7 has been described as a regulator of axon outgrowth, possibly by stimulating nerve growth factor and tyrosine kinases A and B as well as the growth cone plasticity protein, GAP43 (Lei et al., 2001; Veldman et al., 2007; Caiazzo et al., 2010; Blackmore et al., 2012; Li et al., 2017).
Furthermore, several studies have shown that many “axon guidance” molecules play important roles in the development of bone, such as neuropilin 1 and semaphorin 7A (Asgharzadeh et al., 2018; Hou et al., 2018; Xue et al., 2018). Consistently, Caiazzo et al. (2010) reported that Klf7 silencing is detrimental to altering the osteogenic potential of mouse embryonic fibroblasts. In addition, Sox9 was predicted to be a target of miR-145-5p (a downregulated miRNA) (Fig. 4B). Iezaki et al. (2018) revealed that Sox9 is a prominent mediator of Erk5 (MAPK7)-dependent skeletogenesis. Thus, miR-145-5p may inhibit osteogenic differentiation by targeting SOX9.
Consequently, our results indicated that HAG scaffolds may promote bone formation by regulating the differential expression of miRNAs. In detail, we successfully uncovered the statistically significant miRNAs in HAG-hPMSCs and constructed a miRNA–mRNA network through different bioinformatics analyses. However, the differentially expressed miRNAs are only candidate miRNAs, and the miRNAs associated with osteoblast differentiation analyzed by combining biological databases or literature are worthy of attention.
Thus, the present work only provides necessary information about the possible role of miRNAs in promoting bone regeneration of HAG scaffolds. The present study had several limitations. We only compared miRNAs from osteogenically differentiated hPMSCs with and without HAG scaffolds after induction for 7 days, which may have excluded information about the dynamics of miRNAs during osteogenic differentiation. In addition, we did not elucidate the specific mechanism of action of differentially expressed miRNAs in the promotion of osteogenic differentiation by HAG. Therefore, these questions will be explored in future research.
Ethics Approval and Consent to Participate
All experimental protocols were approved by the Medical Ethics Committee of Sichuan Provincial People's Hospital, Affiliated Hospital of UESTC.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
The National Natural Science Foundation of China (82071168), and the Science and Technology Department of Sichuan Province Foundation (2021YFS0009) supported the present study.