
Abstract
The striped knifejaw (Oplegnathus fasciatus) and spotted knifejaw (Oplegnathus punctatus) are prominent members of the Oplegnathidae family and are rocky reef-loving fishes with high ecological and economic value. However, the frequent occurrence of diseases in these fishes has severely restricted the development of their breeding industry. Toll-like receptors (TLRs) play an important role in resistance to pathogens as part of innate immunity. Genome-wide scans and cross-species comparative analysis revealed 10 TLRs in O. fasciatus (OfTLRs) and only 5 in O. punctatus (OpTLRs). In contrast to those of mammals and other fishes, the TLR family of Oplegnathidae underwent significant contraction events, especially in O. punctatus (only TLR1, TLR2, TLR14, TLR5, and TLR21 were retained). A phylogenetic tree divided the 10 OfTLRs into 5 subfamilies: TLR1, TLR3, TLR5, TLR7, and TLR11. The five OpTLR genes were divided into three different subfamilies: TLR1, TLR5, and TLR11. Quantitative real-time PCR revealed that all OpTLRs were expressed in the examined tissues, especially the immune system-related tissues, such as the spleen, gill, head kidney, and middle kidney. The expression of OpTLRs was high at the early stage of development (5 days posthatching [dph]) and decreased gradually until 30 dph. We speculated that maternal immunity or the developmental function of TLRs played an important protective role in the early stage. However, from 30 to 60 dph, TLR expression was low. At this time, juvenile fish are susceptible to viruses and begin to show TLR self-expression with weak immunity. Artificial immunity enhancement is needed to improve the environmental resistance of juvenile fish. In summary, our results not only provide valuable basic data for future studies of the TLR gene family in Oplegnathidae fish but also lay a solid foundation for Oplegnathidae fish research.
Introduction
The immune system of teleost fish consists of innate and adaptive immunity, each of which plays different roles in immune functions (Bai et al, 2017). In general, since most fish are thermophilic, innate immunity can be activated at most temperatures, and adaptive immunity, such as lymphocyte differentiation and antibody production, depends more on higher temperatures. Therefore, fish are more dependent on the innate immune system to resist disease invasion, and innate immunity is the first line of host defense (Rebl et al, 2010; Secombes and Wang, 2012). In the innate immune system, some innate pattern recognition receptors (PRRs), such as Toll-like receptors (TLRs), RIG-I-like receptors, NOD-like receptors, and C-type lectin receptors, play vital roles in responding to pathogens (Rao and Su, 2015).
These PRRs are mainly used to identify pathogen-associated microbial patterns (PAMPs) by conserved structures of invading pathogens. As one of the most important groups of PRRs, TLRs are conserved and ancient innate PRRs in the innate immune system that play a crucial role in recognizing highly conserved structural motifs of PAMPs and danger-associated molecular patterns (DAMPs) that are released by the body during stress or injury and subsequently induce the host immune response (Nie et al, 2018). TLRs consist of an extracellular leucine-rich repeat (LRR) domain for ligand recognition, a transmembrane domain, and an intracellular Toll/interleukin 1 receptor (TIR) signaling domain (Horng and Medzhitov, 2001; Takeda and Akira, 2004). The LRR and TIR domains are used to identify pathogens and for downstream signal transduction, respectively (Liao et al, 2017). To date, research on TLRs has mainly focused on their innate immunity pathway and provided information about their innate immunity mechanism.
TLR genes were first discovered in fruit flies (Drosophila melanogaster) (Anderson et al, 1985); since then, they have been found in vertebrates, including many kinds of teleosts. Among mammals, 10 different kinds of TLRs have been identified in humans, and 12 kinds of TLRs have been identified in mice (Ji et al, 2018). Although there are 10 TLR genes in birds, they are different from those in mammals. There are direct homologs of mammalian TLR1, TLR 2, TLR3, TLR4, TLR5, and TLR7 in birds, but TLR8 and TLR9 are missing (Boyd et al, 2007; St Paul et al, 2013; Smith et al, 2004; Temperley et al, 2008).
As many as 222 TLR genes have been found in echinoderms, such as sea urchin (Strongylocentrotus purpuratus) (Huang et al, 2008); however, only one TLR gene was found in Aenorabdrtis elegans (Wojtkowiak, 2007). In teleost fishes, 21 TLRs have been identified, including TLR1-5, 5S, TLR7-9, TLR13, TLR14, TLR18-23, and TLR25-28. Among them, TLR1-3, 5, and 7-9 are similar in structure and function to those of mammals. However, the TLR4 gene is missing from most fish genomes. In addition, special TLRs of teleost fish include TLR5s, 18-20 and 23, and TLR25-28. Furthermore, the number of TLRs conserved among all vertebrates (only 13 types) is much lower than that in teleosts (21 types) (Sahoo, 2020).
Related research has shown that TLRs have undergone more changes in teleost fish than in mammals during the process of evolution (Cao et al, 2020). Nevertheless, several related studies have suggested that TLRs have sustained conservative structures over the long period of biological evolution (Tafalla et al, 2013) based on the similarity of the key structures of TLRs between mammals and fish (Chen et al, 2020). According to some previous research, the number and kinds of TLRs vary among species, and different TLRs perform different functions. Therefore, studying the TLRs of different species and understanding their characteristics and main roles can help us understand pathogen recognition mechanisms and further develop disease prevention and control measures.
The striped knifejaw and spotted knifejaw fishes, Oplegnathus fasciatus and Oplegnathus punctatus, are among the most important maricultured species and economically important rocky reef marine fish distributed in the Yellow Sea, in the East China Sea, along the Taiwanese coast, near Hokkaido south of Japan, and south of the Korean coast. These species belong to Oplegnathidae family and are famous for their high commercial, ecological, ornamental, and economic value (Du et al, 2017; Xiao et al, 2019).
However, large-scale outbreaks of rainbow virus and neuro necrosis virus in O. fasciatus and O. punctatus during the breeding process caused a high mortality rate during production, which has largely restricted the healthy and rapid development of the breeding industry for Oplegnathidae fish (Jung and Jung, 2017; Jung and Oh, 2000). Furthermore, these virus outbreaks resulted in substantial economic losses for breeders. Therefore, carrying out immuity-related analysis of Oplegnathidae fish will facilitate further exploration of the immune regulatory mechanism in Oplegnathidae fish.
In this study, genome-wide characterization of TLRs in O. fasciatus and O. punctatus was performed, including analysis of their gene family composition, physicochemical properties, chromosomal locations, motif compositions, structural protein domains, phylogenetic relationships, and quantitative expression. In addition, the tissue-specific and developmental expression patterns of TLRs were quantitatively analyzed to understand their expression characteristics. Our systematic study of these TLRs represents a starting point for further exploration of TLRs and provide fundamental data for improving e disease resistance. In addition, the healthy development of the Oplegnathidae breeding industry should be promoted.
Materials and Methods
Genome-wide identification and characterization of TLRs in Oplegnathidae
To identify the TLRs in O. fasciatus and O. punctatus, the available TLR genes of other species, such as Homo sapiens, Mus musculus, and other teleost fishes, were downloaded from the NCBI genome database (
After combining the HMM search results and BLAST results, the candidate TLR genes in O. fasciatus and O. punctatus were identified. The sequence obtained was named OfTLR and OpTLR, respectively. The molecular weights (MWs) and isoelectric points (PIs) of the candidate TLR genes were evaluated by the online site EXPASy (
Phylogenetic analysis of TLR genes in Oplegnathidae
To explore the phylogenetic relationships of TLR genes between Oplegnathidae fish and mammals, birds, and other teleost fish, we downloaded homologous TLR genes of these species from the NCBI database to construct a local dataset. Then, these downloaded protein sequences were aligned by MUSCLE from TBtools (Edgar, 2004). Finally, MEGA 11 was used to construct a neighbor-joining phylogenetic tree. Bootstrapping with 1000 replicates was applied to quantify the confidence of each node in the tree (Quiniou et al, 2013). Furthermore, the downloaded protein sequences were used to build a map for visualizing TLR gene numbers in these species.
Experimental materials
This study was approved by the Animal Care Committee of Institute of Oceanology, Chinese Academy of Sciences (IOCAS), and carried out in accordance with guidelines for research and teaching animal care and use of the IOCAS (ethics code permit No. 2022C007). We used MS-222 (Hengfei, China) at a dose of 40 mg/L to anesthetize the fish before the experiment to minimize suffering and pain.
Three experimental O. punctatus fish were obtained from Yantai Mingbo Aquatic Products Co., Ltd. The adult fish had an average body weight of 206.1 ± 6.6 g and an average body length of 16.9 ± 0.6 cm. The fish were cultured in water at 24.0°C ± 0.9°C. Of the three fish, two were male and one was a female. To explore the expression patterns of TLR genes in healthy fish tissues, we collected tissue samples from the three adult fish. Eight healthy tissues were used for RNA extraction: gill, liver, spleen, head kidney, middle kidney, brain, heart, and skin.
In addition, individuals of O. punctatus were sampled at different developmental stages, namely 5 days posthatching (dph) (average body length 0.37 ± 0.02 cm), 16 dph (average body length 0.53 ± 0.04 cm), 30 dph (average body length 1.20 ± 0.12 cm), 50 dph (average body length 1.94 ± 0.22 cm), and 60 dph (average body length 3.67 ± 0.30 cm). O. punctatus fish in the above developmental stages were obtained from Haihe Turbot Nursery in Wenden, Weihai. Whole fish were used as samples for all developmental periods, with three replicates for all the sampled fish. The samples were placed into freezing tubes for rapid addition of liquid nitrogen. Finally, the tubes containing samples were placed in a −80°C freezer for preservation.
RNA extraction and cDNA synthesis
Total RNA of all samples mentioned above was extracted using the TRIzol (Invitrogen) extraction method. The integrity of the RNA was determined by 1% agarose gel electrophoresis (after dissolution with Tris Acetate-EDTA buffer). An Ultramicro spectrophotometer (NanoDrop2000) was used to detect the concentration and purity of the RNA (where OD260 nm OD280 nm ≥1.8 and OD260 nm OD230 nm ≥1 indicated good quality). An Evo M-MLV RT Kit (Accurate Biology, Changsha, China) was used to synthesize first-stand cDNA according to the manufacturer's instructions, and the cDNA was stored at −20°C.
Quantitative real-time PCR analysis of OpTLR genes
Quantitative real-time PCR (qRT-PCR) was used to detect the mRNA levels of TLR genes in different tissues and developmental periods with a CFX96™ real-time detection system (Bio-Rad). Specific primers of TLR genes were designed for qRT-PCR analysis by the NCBI (Table 1). The qRT-PCR mixture consisted of 10 μL SYBR Green I, 0.4 μL (10 μmol/L) forward and reverse primers, 7.2 μL sterilized dd H2O, and 2 μL cDNA for a total volume of 20 μL. The PCR amplification program was as follows: 95°C for 30 s, followed by 39 cycles of 94°C for 5 s and 59°C for 30 s. The melting curve was constructed with the following program: 94°C for 10 s, 65°C for 5 s, and 94°C for 5 s.
Primers Used for Expression Profiling
Pfaffl's (2001) method was adopted to detect the relative expression level of target TLRs. For tissue-specific expression analysis, the expression level in the gills of individuals was used as the baseline, and the expression levels in the other tissues were expressed relative to this baseline. For stage-specific analysis of expression, the expression level at 5 dph was used as the baseline, and the expression levels in other developmental stages were expressed relative to this baseline. In addition, the housekeeping gene β-actin was utilized as a reference gene for all cDNA normalization (Wang et al, 2020). All specific primers were synthesized by TsingKe Biotechnology Co., Ltd.
Statistical analysis
TLR gene expression levels (mean ± standard deviation) were analyzed by one-way ANOVA, in R software (version 4.1.1) with p < 0.05 indicating statistical significance.
Results
Sequence analysis of OfTLRs and OpTLRs
Combining the HMM search results with the BLAST results, a total of 10 available OfTLR genes and 5 OpTLR genes were identified in the genomes of O. fasciatus and O. punctatus, respectively. These 10 sequences were named OfTLR1, OfTLR2a, OfTLR2b, OfTLR3, OfTLR5, OfTLR7, OfTLR9, OfTLR23a, OfTLR23b, and OfTLR21, and belong to the TLR1, TLR3, TLR5, TLR7, and TLR11 subfamilies. There were only five TLRs in O. punctatus, named OpTLR1, OpTLR2, OpTLR5, OpTLR14, and OpTLR23, which belonged to the TLR1, TLR5, and TLR11 subfamilies, respectively.
Table 2 provides information about the chromosomal locations, mRNA and protein lengths, MWs, and PIs of these TLRs. The protein length of OfTLRs ranged from 787 (OfTLR2b) to 1128 (OfTLR21) amino acids. The maximum MW of OfTLR proteins was observed for OfTLR23a (127.51 kDa), while the minimum was observed for TLR2b (89.8 kDa). The theoretical PI varied from 5.52 (OfTLR23a) to 8.99 (OfTLR3). In O. punctatus, the longest protein was OpTLR14, and the MW varied from 79.78 kDa (OpTLR23) to 105.81 kDa (OpTLR14).
Characteristics of Toll-Like Receptor Genes in Oplegnathidae Fish
MW, molecular weights.
Chromosomal location and protein domain analysis of TLR genes
As shown in Figure 1, the results of gene localization analysis show that 10 OfTLRs and 5 OpTLRs were distributed on different chromosomes. Specifically, OfTLR2b and OfTLR3 were located on chromosome 6, OfTLR14 and OfTLR21 were located on chromosome 3, and OfTLR23a and OfTLR23b were located on chromosome 21.

Chromosomal locations of TLR genes in Oplegnathidae.
To obtain a better understanding of the physiological functions of these candidate TLR genes, their domains were analyzed. The TLRs were found to contain three main domains, including a TIR, a transmembrane region (TM), and between 7 and 16 LRR domains, as shown in Figure 2.

The conserved domain of the TLR protein in Oplegnathidae.
Motif composition and sequence alignment of TLR genes
To obtain further information about the structural diversity of the TLR family in Oplegnathidae fish, the MEME online software was applied to predict the conserved motifs of TLR family proteins. Fifteen different conserved motifs were identified. The phylogenetic relationships between Oplegnathidae fish and two other teleost species were combined with the conserved motifs (Fig. 3). The conserved motifs in distinctive TLR subfamilies remained highly consistent and revealed similar protein functions among members of the same subfamily.

Conserved motif analysis of different bony fish TLR proteins based on a phylogenetic tree. The abbreviations of species are as follows: Dr: Danio rerio, Aj: Argyrosomus japonicus, Of: Oplegnathus fasciatus, Op: Oplegnathus punctatus.
To better understand how the TLR genes of Oplegnathidae fish activate downstream regulatory pathways, DNAMAN was used to analyze the structural domain of TLR genes and identify conserved regions. The TLRs showed three conserved regions, named BOX1, BOX2, and BOX3. In BOX2, the amino acid sequence was very conserved, except in OfTLR1 and OpTLR1. In addition, an insertion event occurred in the OfTLR23b gene. However, a deletion event occurred in BOX3 (Liu et al, 2019) (Fig. 4).

The TIR domains of Oplegnathidae were compared by DNAMAN. The red boxes BOX1, BOX2, and BOX3 represent the conserved structural domains and the black boxes indicate the TIR structural domains. TIR, toll/interleukin 1 receptor.
Distribution and evolutionary analysis of TLR genes
To understand the distribution of TLR families in other species, such as some mammals, birds, and other teleost fishes, IBS was used to plot the distribution of TLRs in different species. TLR1, TLR2, and TLR5 were ubiquitous in most species, while TLR4 and TLR8 were lost in O. fasciatus compared to other teleost fishes. In contrast to other teleost fishes, O. fasciatus possessed only two copies of TLR2 and TLR13. In O. punctatus, however, no copy of TLR genes was present. Most of the TLRs were lost in O. punctatus, such as TLR3, TLR7, TLR8, TLR9, TLR13, and TLR21, in contrast to O. fasciatus (Fig. 5).
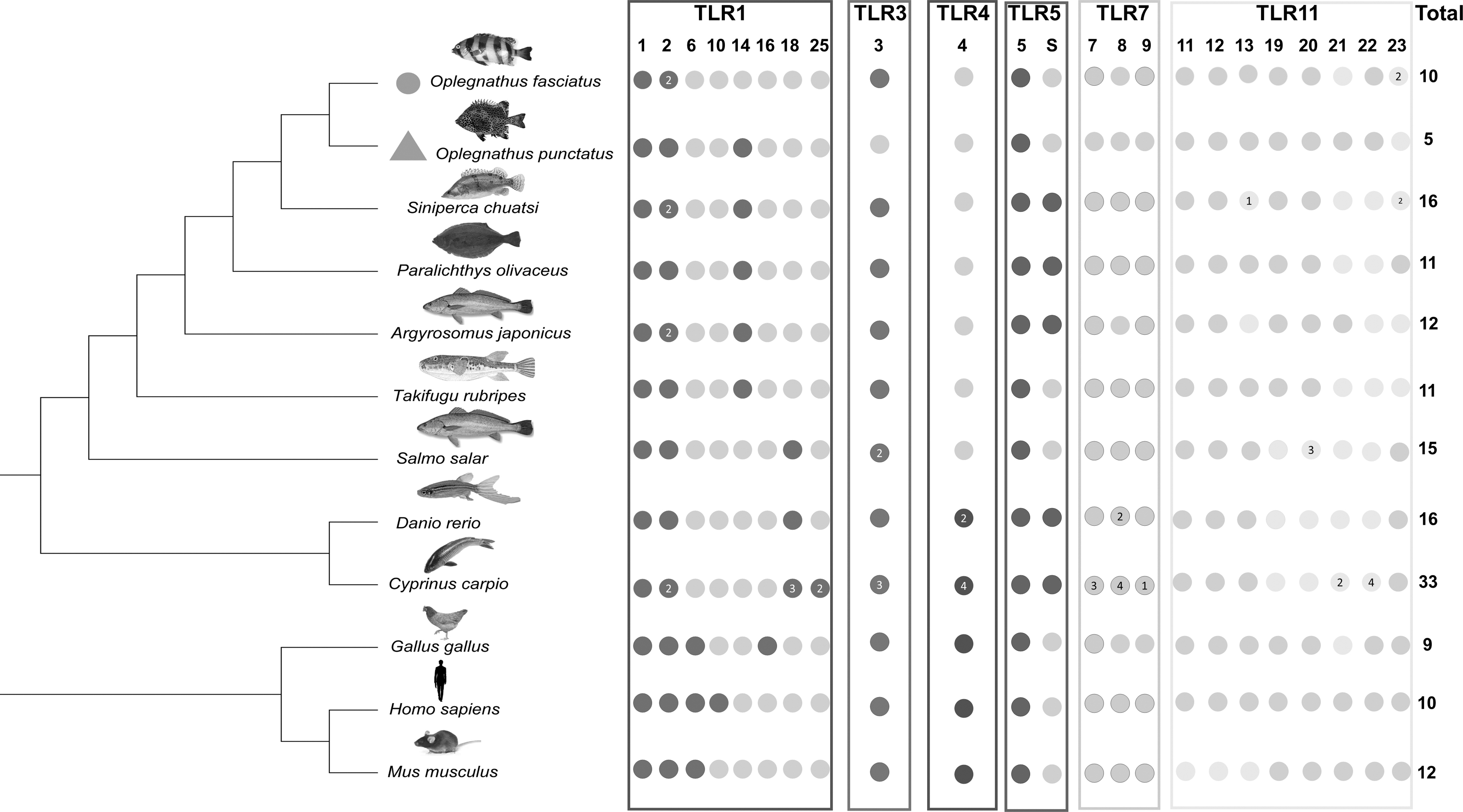
Distribution of TLRs in different species. The TLR genes identified in different species are indicated by colored circles. When there are multiple identical TLR genes identified, gene numbers are indicated within colored circles. Genes that were not identified are indicated by gray circles.
To establish the phylogenetic relationships within the TLR gene family, we constructed a tree based on the amino acid sequences of the other 10 species, as shown in Figure 6. The results of the evolutionary tree showed that the TLRs could be divided into eight groups, that is, TLR1, TLR3, TLR4 TLR5, TLR7, TLR8, TLR9, and TLR11, which belong to six main subfamilies: the TLR1, TLR3, TLR4, TLR5, TLR7, and TLR11 subfamilies. The TLR7 subfamily is divided into three branches: TLR7, TLR8, and TLR9. In addition, OfTLR1, OfTLR2a, OfTLR2b, OpTLR1, OpTLR2, and OpTLR14 belong to the TLR1 subfamily; OfTLR3 belongs to the TLR3 subfamily; OfTLR5 and OpTLR5 belong to the TLR5 subfamily; OfTLR7 and OfTLR9 belong to the TLR7 subfamily; and OfTLR23a, OfTLR23b, and OfTLR21 belong to the TLR11 subfamily.
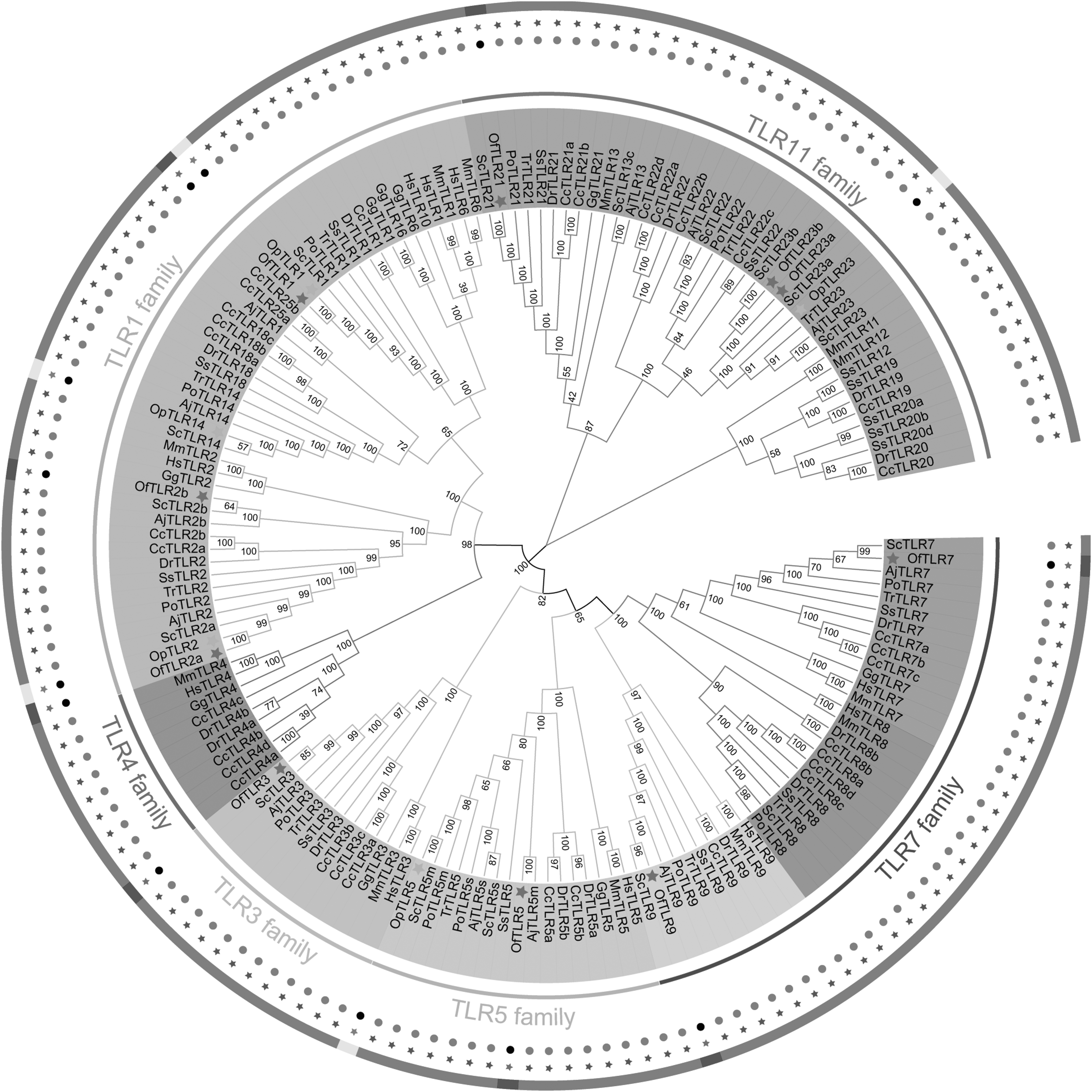
Phylogenetic analysis tree of TLR gene family in Oplegnathidae. There are 160 amino acids involved in the tree. Twelve species were selected to build the evolutionary tree. The abbreviations of species are as follows: Hs: Homo sapiens, Mm: Mus musculus, Gg: Gallus gallus, Of: Oplegnathus fasciatus, Op: Oplegnathus punctatus, Dr: Danio rerio, Aj: Argyrosomus japonicus, Po: Paralichthys olivaceus, Cc: Cyprinus carpio, Tr: Takifugu rubripes, Sc: Siniperca chuatsi, and Ss: Salmo salar. Green stars represent the TLR genes in O. fasciatus and yellow stars represent the TLR genes in O. punctatus. Three circles in the shape of dots, stars, and interrupted color segments from the inside out are used to mark TLR genes in O. fasciatus and O. punctatus. In O. punctatus, the position of TLR genes is represented as black dots, red stars, and yellow color segments, while in O. fasciatus, the position of TLR genes are represented as black dots, green stars, and blue color segments. Different color modules are used to distinguish between different gene families (Supplementary Table S1).
OfTLRs and OpTLRs were always clustered with ArgTLRs and ScTLRs, and the results revealed that they are more closely related. However, O. fasciatus is not the closest relative of O. punctatus. It was obvious that seven species of bony fish were separated from the two species of mammals and birds. Overall, the results of phylogenetic analysis supported the annotation of the O. fasciatus and O. punctatus TLR gene families, and these families were classified with other vertebrate homologous genes, with high bootstrap support value.
Tissue distribution of the TLR gene family in O. punctatus
To understand the expression patterns of these genes in the immune system tissues, quantitative expression analysis of tissue was carried out. The expression of five TLR genes differed in O. punctatus. OpTLRs were widely expressed in all tested tissues, but there were differences in expression level in each tissue (Fig. 7).
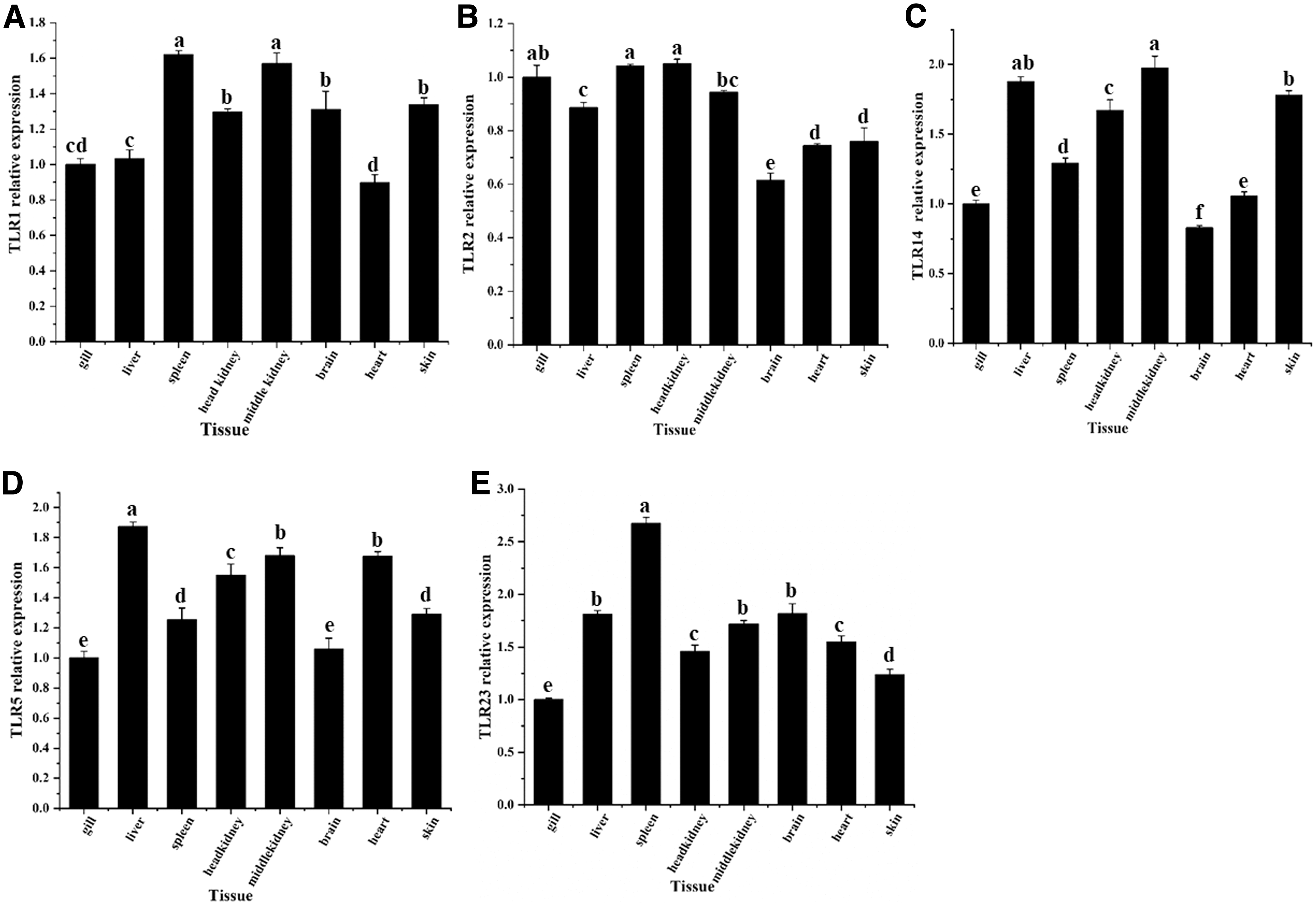
Expression of TLR genes in different tissues of adult O. punctatus. (
The TLR1 family is composed of OpTLR1, OpTLR2, and OpTLR14, which were expressed in the examined tissues. OpTLR1 was highly expressed in the spleen and middle kidney and showed lowest expression in the heart of O. punctatus. The mRNA expression level of OpTLR2 was highest in the spleen and head kidney, and lowest in the brain. The expression pattern of OpTLR14 was similar to those of OpTLR1 and OpTLR2; OpTLR14 was highly expressed in the middle kidney, and expressed at the lowest level in the brain.
Members of the TLR5 family were mainly expressed in the liver, as shown in Figure 8. OpTLR5 displayed high expression levels in the liver, while its expression levels in the spleen and head kidney were much lower than those in the liver (p < 0.01).
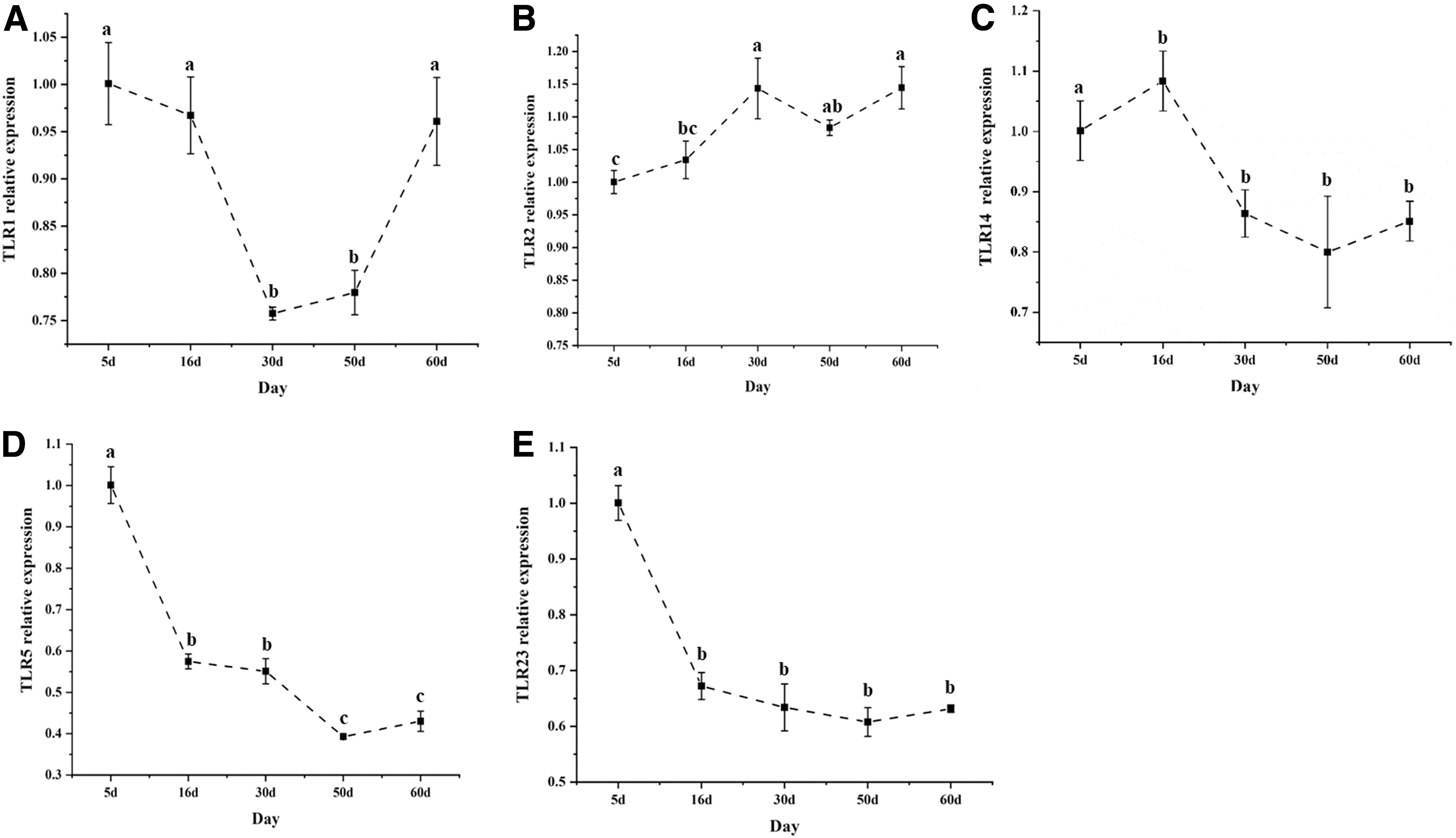
Expression of TLR genes during the developmental period of O. punctatus. (
TLR23, the only member of the TLR11 family, was mainly expressed in the spleen, followed by the liver and brain, and had the lowest expression level in gills.
Expression patterns of OpTLR genes during development
The relative expression of OpTLR genes in O. punctatus was detected at different developmental stages. The goal was to understand the point during development when these genes play a role. The expression of OpTLR1, a member of the TLR1 family, first decreased and then increased, reaching its lowest value at 30 days, and gradually increasing thereafter. The expression level of OpTLR2 and OpTLR14 first increased and then decreased and reached their highest values at 30 and 16 days, respectively. However, the final expression level (60 days) of OpTLR2 was higher than the initial expression level (5 days), while the final expression level (60 days) of OpTLR14 was very significantly lower than its initial expression level (5 days) (p < 0.001).
Furthermore, the expression trends of OpTLR5 and OpTLR23 families were consistent: both decreased sharply from 5 to 30 days and then increased steadily from 50 to 60 days, but were always lower than the initial expression level (5 days) (p < 0.001).
Discussion
In lower vertebrates, such as fish, innate immunity is essential to the fish immune response because it is the main line of defense against the attack of pathogenic microorganisms (Rajme-Manzur et al, 2021). In innate immune responses, the response to an antigen is based on encoded PRRs, which produce an immune response by recognizing PAMPs and DAMPs (Tan et al, 2014). Among the PRRs of innate immunity, TLRs are critical and the earliest studied receptor for recognizing pathogen molecules (Iwasaki and Medzhitov, 2004; Takeda and Akira, 2015). Over a long evolutionary history, the number of TLRs has changed, with different species evolving different numbers of TLRs through adaptation. However, the structure of TLR genes (containing TIR, TM, and LRR domains) is conserved across species (Zhao et al, 2020).
Common carp (Cyprinus carpio) and Atlantic cod (Gadus morhua) have been found to have 27 TLR genes (Gong et al, 2017; Star et al, 2011). Twenty-one TLRs have been identified in grass carp (Ctenopharyngodon idella) and channel catfish (Ictalurus punctatus) (Liao et al, 2017; Zhao et al, 2013a). For mandarin fish (Siniperca chuatsi), Japanese meagre (Argyrosomus japonicus) and black rockfish (Sebastes schlegelii), 16, 11, and 17 TLRs have been identified and characterized, respectively (Cao et al, 2020; Han et al, 2022; Wang et al, 2021). According to a recent study by Sousa et al (2022), who compared TLR members in temperature regions, the Antarctic Nototheniidae species and non-Antarctic (temperate) species in the sister lineage of Bovichtidae, no major TLR gene family expansion and contraction occurred, except in Dissostichus mawsoni.
In all Notothenioidei (except D. mawsoni) and other teleosts, two TLR2 genes appeared to be duplicated and to have arisen during genome duplication events specific to teleosts, with paralogs termed TLR2a and TLR2b. At the same time, the authors concluded that the ancestral genes of TLR5 originated in the TLR5 and TLR5s subfamilies and replicated during adaptive radiation in teleosts. During the long evolutionary process, TLR genes were conserved across most teleosts (Sousa et al, 2022). Li (2020) found that several gene families related to immunity contracted in a comparative genomic study, including O. punctatus.
In this study, we identified 10 OfTLR and 5 OpTLR genes at the genome-wide level. According to the number and variety of TLRs in Oplegnathidae, we concluded that the TLR gene family of Oplegnathidae has contracted significantly. A similar species-specific gene deletion occurred in Cynoglossus semilaevis, according to the study by Sousa et al (2022). Therefore, it is speculated that this may be one of the reasons why Oplegnathidae is susceptible to disease or that TLR gene contraction is the result of adaptive evolution.
According to the amino acid sequence alignment results and the phylogenetic tree, there were many differences between OfTLR2a and OfTLR2b, and they may perform different functions. In terms of domain composition, although both OfTLR2a and OfTLR2b contain TIR domains, there is one more LRR-N-terminal domain in OfTLR2a than in OfTLR2b, and the number and location of LRRs in the two are different. OfTLR2a and OfTLR2b were on different branches in the phylogenetic tree.
Therefore, although gene duplication occurred in TLR2 during the long evolutionary process, their functions were different from those of existing genes, similar to the conclusion reached in a study of Japanese meagre (A. japonicus) (Han et al, 2021). This type of phenomenon was also observed for the TLR5 family of O. fasciatus and O. punctatus. In the phylogenetic tree, OfTLR5 and OpTLR5 were not on the same branch, and amino acid sequence alignment revealed different bases in the conserved region, of the two genes.
qRT-PCR was used to detect the expression of each TLR gene in the tissues of healthy O. punctatus. The tissue expression distribution showed that all the OpTLRs were widely expressed in all tested tissues. However, these TLR genes were highly particularly expressed in immune system tissues. Due to adaptive immunity, teleost fish have more TLRs than mammals (Liao and Su, 2021).
In addition, TLR6 and TLR10 are absent from the teleost fish TLR1 family, which differs from the pattern in higher vertebrates (Su and Yu, 2019). TLR1 in fish is said to be more closely related to TLR1, TLR6, and TLR10 in mammals than to other TLRs in birds and mammals (Kawai and Akira, 2007). In mammals, however, TLR14 is missing from the TLR1 family. In contrast, TLR14 has been identified in most teleost fishes, such as golden pompano (Trachinotus ovatus) (Wu et al, 2019), orange-spotted grouper (Epinephelus coioides) (Li et al, 2016), and Miiuy croaker (Miichthys miiuy) (Zhang et al, 2014).
In O. punctatus, TLR1, TLR2, and TLR14 belonging to the TLR1 family are expressed in immune system tissues such as the spleen, gill, head kidney, middle kidney, and liver, and there are low levels of expression in the brain and heart. The results of this study were consistent with those for orange-spotted grouper (E. coioides) (Wei et al, 2011) and rainbow trout (Oncorhynchus mykiss) (Palti et al, 2010).
These results revealed TLR1 mRNA expression in all tissues, mainly in the spleen and head kidney. Furthermore, in Japanese flounder (Paralichthys olivaceus), the expression of TLR2 was detected in relevant immune system tissues, such as the spleen and head kidney (Hirono et al, 2004). The expression of TLR14 was not the highest in the immune system tissue, conclusion also reached in O. punctatus. According to the above studies, the TLR1 family plays an important role in the immune response of teleost fish; members of this family might not only participate in a crucial immune response in teleost fish but also perform other physiological functions that have not been discovered.
TLR5 is present in both mammals and teleost fish, and it has been shown to recognize flagellin components of bacteria and be responsible for flagellin-mediated NF-κB activation (Tsujita et al, 2004). In Japanese flounder (P. olivaceus), TLR 5m was highly expressed in the gill, head kidney, heart, and liver. In addition, TLR5s was highly expressed in the brain, head kidney, and heart (Hwang et al, 2010). Real-time PCR was used to examine the expression of TLR5 in pufferfish (Fugu rubripes), revealing that TLR5 was widely expressed (Oshiumi et al, 2003). In this study, the same conclusion was also reached: TLR5 is widely expressed in all tissues. Furthermore, TLR5 was highly expressed in the liver, followed by the middle kidney and heart. In a study of rainbow trout (O. mykiss), TLR5m was ubiquitously expressed, whereas TLR5s were predominantly expressed in the liver (Tsujita et al, 2004).
Another study found that both TLR5-1 and TLR5-2 are highly expressed in the liver and gonads, while TLR5-2 is more highly expressed in the kidney in channel catfish (I. punctatus) (He et al, 2019). In teleost fish, intense activation of NF-κB occurs through the combination of two forms of TLR5. The advantages of this flagellin recognition system are comparable to those of TLR4 in LPS recognition (Akashi et al, 2003; Poltorak et al, 2000). However, there is only one TLR5 in O. punctatus, and its expression pattern was the same as those in the above fish. Its mechanism of action in the downstream pathway will need to be further explored.
Among numerous studies on teleost fish, some “fish-specific” TLRs were identified: TLR5s, TLR18, TLR19, TLR20, TLR23, TLR25, TLR26, TLR27, and TLR28 (He et al, 2019; Wang et al, 2015). Only TLR23 is found in O. punctatus, where it is mainly expressed in the spleen, followed by the liver, middle kidney, and brain. TLR23 is highly expressed in the brain, which differs from the expression pattern of other TLR genes.
This expression pattern has also been found in other species; for example, TLR23 mRNA levels were highest in the brains of Japanese meagre (A. japonicus) (Han et al, 2021). In Atlantic cod (G. morhua), the expression level of TLR23a was higher compared with TLR23b, both of which were expressed in the head kidney, kidney, spleen, gills, blood, and testis. In addition, TLR23a transcripts were also more abundantly expressed in the liver, heart, and brain (Sundaram et al, 2012). To date, there have been few studies on TLR23, and its mechanism in fish needs to be further studied. Based on the above studies, it is speculated that TLR23 is also involved in other physiological processes in O. punctatus.
The expression levels of TLRs vary at different stages of fish development, indicating that the types of TLRs that play a dominant role at different stages of development are also distinct. In the early stages of common carp (C. carpio), that is, within the first 24 h, only the TLR2-1 gene was expressed at a high level. At 36 h, each TLR gene began to be expressed significantly. However, most genes had low or no expression at 48 h. Over the next 3 to 7 dph, the expression of different genes varied. At 3 dph, TLR1 and TLR5 began to be highly expressed; TLR3, TLR9, and TLR19 showed a high expression level at 5 dph; and a large number of TLR genes, especially TLR1, TLR8, and TLR19, were highly expressed again at 7 dph (Gong et al, 2017).
In this study, we found that all OpTLR genes maintained high expression levels at 5 dph, except for OpTLR2, and the expression levels of TLR14, TLR5, and TLR23 decreased significantly at the late development stage, while the expression levels of TLR1 and TLR2 remained high. It can be concluded that some genes are expressed in the early stage of development, and few genes are expressed in the later stage of development. It is hypothesized that maternal immunity plays an important protective role during the early posthatching period (5–16 dph). Then the expression of TLRs reaches a minimum level at 30 and 50 dph. Then, TLR self-expression began because maternal immunity has been exhausted.
On the other hand, studies in Drosophila and some mammals have shown that TLR genes are closely associated with development. Combining these results, the results of this article, we hypothesize that TLR genes show higher expression during early development and are likely to play a role in development (Anderson et al, 1985; Sousa et al, 2022; Souza and Lerner, 2019). According to the research of He (2011) and Xiao (2014), the prelarval mortality rate was higher before 60 dph, during which prelarvae were susceptible to the studied virus. This phenomenon coincided with the expression of TLR genes: TLR expression was low when the immunity of the prelarva was weak.
In addition, members of the TLR1 family are involved in almost the whole development process, suggesting that TLR1 plays an irreplaceable immunity-related role in the development process. Such a role of the TLR1 family has also been demonstrated in other studies, such as in pufferfish (Tetraodon nigroviridis) (Wu et al, 2008) and channel catfish (I. punctatus) (Zhang et al, 2013b).
Conclusion
In summary, based on the genome and other available data, we identified 10 TLR genes in O. fasciatus and 5 TLR genes in O. punctatus, representing the first systematic collation and analysis of TLR genes in Oplegnathidae. Compared with those of other bony fishes, the TLR gene family of Oplegnathidae fishes contracted significantly. The characteristics of a total of 10 OfTLRs and 5 OpTLRs were analyzed, such as their chromosomal localization, protein domains, motif composition, conserved regions, and phylogenetic relationships. The OpTLRs were widely expressed in all tested tissues, but the expression level was higher in immune system tissues.
Notably, all OpTLR genes were expressed in the early stage of development. Furthermore, except for OpTLR2, f OpTLRs showed higher expression levels in the early stage of development, which might be due to the depletion of maternal immunity. Since the self-expression of OpTLRs began in late development, the expression of TLRs was low at that time. Research on TLR genes in Oplegnathidae will provide basic information for further understanding the immune response of Oplegnathidae and other fishes.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the National Key Research and Development Program (2018YFD0901204), the Key Special Project for Introduced Talents Team of the Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou) (GML2019ZD0402), Key Deployment Projects of Center for Ocean Mega-Science, Chinese Academy of Sciences (Fron-tier Cross-category, COMS2020Q05), CARs for Marine Fish Culture Industry (CARS-47), the Major Agricultural Application Technology Innovation Project of Shandong Province (SD2019YY011), the STS project (KFZD-SW-106, ZSSD-019, 2017T3017, and KFJ-STS-QYZX-020), the Qingdao National Laboratory for Marine Science and Technology (2018SDKJ0502-2 and2015ASKJ02), and the National Natural Science Foundation of China (No. 31672672).
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.