
Abstract
Human dermal fibroblasts (HDFs) can be reprogrammed through different strategies to generate human induced pluripotent stem cells (hiPSCs). However, most of these strategies require high-cost materials and specific equipment not readily accessible in most laboratories. Hence, liposomal and virus-based techniques can replace with polyethylenimine (PEI)-mediated transfection to overcome these challenges. However, few researchers have addressed the PEI's ability to transfect HDFs. This study used PEI reagent to transfer oriP/EBNA1-based vector into HDFs to produce hiPSC lines. We first described conditions allowing the efficient transfection of HDFs with low cytotoxicity and without specific types of equipment and optimized several parameters relevant to the transfection procedure. We then monitored the effect of different N/P ratios on transfection efficiency and cytotoxicity using flow cytometry and fluorescent microscopy. By the results, we found that transfection efficiency was greatly affected by plasmid DNA concentration, PEI concentration, order of combining reagents, serum presence in polyplexes, and the duration of serum starvations. Moreover, using the optimized condition, we found that the N/P ratio of 3 achieved the highest percentage of HDFs positive for green fluorescent protein plasmid (∼40%) with minimal cell toxicity. We finally generated hiPSCs using the optimized protocol and oriP/EBNA1-based vectors. We confirmed hiPSC formation by characterizing tests: alkaline phosphatase staining, immunocytochemistry assay, real-time PCR analysis, in vitro differentiation into three germ layers, and karyotyping test. In conclusion, our results indicated that 25 kDa branched PEI could efficiently transfect HDFs toward generating hiPSCs via a simple, cost-effective, and optimized condition.
Introduction
In the last two decades, there has been a growing interest in nanoparticle delivery systems as promising nonviral reagents for generating induced pluripotent stem cells (iPSCs) (Rony et al., 2015; Sivandzade and Cucullo, 2021) due to their great potential to enhance clinical regenerative medicine and stem cell research (Liu et al., 2015). Among these vectors, cationic lipids are commonly administered in the iPSC technique. However, the cost of such commercial reagents such as Lipofectamine can be a limiting factor for laboratories facing financial problems. In addition, several other successful nanoparticulated delivery vehicles have been introduced in the literature for generating iPSCs, including polyketal nanoparticles, cationic bolaamphiphile, poly-b-amino esters, and calcium phosphate nanocomposite (Cao et al., 2013; Khan et al., 2013; Sohn et al., 2013) [see Liu et al. (2015) for a comprehensive review].
Although the research on a greater understanding of such nanoparticles has been conducted with promising results, the optimized protocol and the properties enabling these nanoparticles to overcome several barriers to intracellular DNA delivery still need further investigations (Guo et al., 2017; Lin et al., 2021). For example, despite the successful reprogramming ability of poly-b-amino esters, integrated transgenes were found in clones of human induced pluripotent stem cells (hiPSCs) and further investigations are still required to evaluate the feasibility of poly-b-amino ester-plasmid as a nonviral reagent (Liu et al., 2015). Moreover, evidence have shown more potency of polyethylenimine (PEI) for delivering genes of interest into cells in comparison to one of the most common nanoparticulated delivery vehicles; calcium phosphate (D'Mello et al., 2016).
Given above, despite the common usage of different nonviral reagents for generating iPSCs from a wide range of somatic cells, cationic polymers such as PEI, which is one of the golden standard reagents for nucleic acid delivery applications (Meleshko et al., 2017; Srisomboon et al., 2021), have been less investigated in iPSC technology. PEI belongs to one of the most studied and effective families of manufactured polycations, which are available with different sizes (kDa) and structures (linear or branched) depending on the particular synthesis procedure (Lázaro-Martínez et al., 2015). A great interest has been shown toward PEI due to its ability to condense DNA into compact particles, uptake into compact particles, release from endosomal compartment into the cytoplasm, and uptake of the DNA into nucleus (Liu et al., 2020).
Wang et al. (2017) showed that the total amount of plasmid DNA (pDNA) within the cytosol was similar among cultured cells after transfection, showing PEI's prominent ability to penetrate cell membranes (Wang et al., 2017). Seo et al. (2013) also evaluated the efficiency of poly (D,L-lactide-co-glycolide) [PLAG]/PEI/DNA nanoparticles in transfection and virus production in comparison with liposome-DNA complexes (Seo et al., 2013). The authors showed superior transfection efficiency and viral yield in PLAG/PEI/DNA group when compared with liposome-DNA complexes, particularly in multiple gene transfections.
These properties make PEI a good alternative for the delivery of desired genes and nucleic acids (Rodier et al., 2019). However, despite the known considerable strength of PEI in the cell transfection process, not much attention has been given to the application of PEI in iPSC generation, which is a novel and gold strategy in personalized as well as regenerative medicine. In a comparable attempt, Drozd et al. (2015) investigated the generation of hiPSCs from cells of fibroblastic and epithelial origin by means of episomal reprogramming vectors via four reagents; Lipofectamine, PEI, FuGENE6, and FuGENE HD. At the preliminary stages, they observed the highest expression levels achieved from PEI and FuGENE6 reagents (Drozd et al., 2015). Although PEI gave the highest transfection efficiency, the authors decided to use FuGENE6 for further experiments in their study due to lesser cytotoxic effects.
Therefore, PEI dose-dependent toxicity on cells and tissues has remained unclear and controversial (Craciun et al., 2019). The cytotoxicity and efficiency of PEI have been linked to PEI's physicochemical properties such as molecular weight, branching ratio, and particle size (Neu et al., 2005; Mintzer and Simanek, 2009). Most techniques have focused on refining PEI size, branch density, and deacylation to reduce PEI-mediated cell toxicity. But these methods are labor-intensive and time-consuming (Jiang and Salem, 2012; Sawant et al., 2012). Besides such methods, the way the polyplex is prepared can also significantly impact transfection efficiency and cytotoxicity (Zhao et al., 2009).
So considering several parameters such as DNA amount, PEI amount, the ratios of moles of the amine groups of cationic polymers to those of the phosphate ones of DNA (N/P ratio) assists further optimizing transfection efficiency and cytotoxicity of polyplexes. It is well documented that PEI exhibits the highest transfection efficiency among nonviral vectors under adjusted parameters and conditions (Srisomboon et al., 2021; Zhu et al., 2021), although there are few data on the optimized protocol for generating hiPSCs administrating PEI. So, to better clarify the properties of PEI/pDNA complexes for generating hiPSCs from human dermal fibroblast (HDF), we intended to optimize several conditions regarding PEI/pDNA complexes, which is missing in the current literature.
In this study, we investigated the influence of different parameters relevant to the formation of PEI-pDNA complex, such as pDNA concentration, PEI concentration, order of combining reagents (i.e., PEI, pDNA), serum content of polyplexes, and serum starvation of cells before transfection. Then, after optimizing the polyplex preparation, we studied the influence of N/P ratio on HDF transfection to find the most efficient protocol for generating hiPSC lines, in which 25 kDa branched PEI and oriP/EBNA1-based vectors were used.
Therefore, in a novel attempt, we aimed to develop an efficient protocol for the PEI-mediated transfection of HDF toward hiPSCs comparable in ease to other nonviral commercial transfection reagents such as lipofectamine. We believe that our findings provide experimental evidence that could be usefully applied in other experiments in the area of safe and cost-effective iPSC generation.
Materials and Methods
HDF culture
Primary HDFs were purchased from Royan Institute, Stem Cell Biology and Technology Department (Chen et al., 2007; Totonchi et al., 2009). IRB approval code is 96-03-87-31967 registered in Iran University of Medical Sciences.
PEI/pGFP (green fluorescent protein) complexes
One mg branched PEI (molecular weight of 25 kDa) was purchased from Polysciences, Inc., and dissolved in 1 mL sterile water and filtered with a 0.25 μm syringe filter. Green fluorescent protein plasmid (pGFP) (#64904; Addgene) was used to detect the most efficient PEI-mediated gene delivery (Ye et al., 2007). Then, immunofluorescence imaging and flow cytometry were performed to detect transfected cells.
At the first step, we used N:P ratio of 12 to test different conditions and to find the best-optimized protocol relevant to complex formation. For this purpose, pDNA concentration, PEI concentration, order of combining reagents, fetal bovine serum (FBS) content of complexes, and duration of serum starvation before transfection were adjusted (see Table 1 for more details on the different conditions of complex formation). Based on the optimized protocol of complex formation, we assessed the impact of different N/P ratios on transfection efficiency and cell toxicity. We prepared different N/P ratios of PEI/pGFP complexes, including 3, 4, 6, 8, 10, and 12.
Different Conditions of Complex Formation to Reach the Best Optimized Condition
PEI concentration of 40 μg/mL was obtained by diluting PEI in distilled water in a ratio of 1:25 (pGFP volume adjusted to the PEI volume in each group to keep the N/P ratio of 12).
FBS, fetal bovine serum; pDNA, plasmid DNA; PEI, polyethylenimine; PEI/pDNA, PEI added to content of pDNA microtube; pDNA/PEI, pDNA added to content of PEI microtube.
Transfection efficiency assay: Immunofluorescence imaging and flow cytometry
Transfection efficiency of different conditions and N/P ratios was assessed by immunofluorescence imaging under GFP filter and flow cytometry technique 48 h post-PEI/pGFP transfection. The gating was performed based on the untransfected cells considering as the control group. The GFP-positive HDFs were investigated statistically to determine the efficiency of gene transfection. Finally, the data were analyzed with Flow Jo software version 10. All tests were performed in triplicate.
Cell cytotoxicity evaluation: MTT assay
The impacts of different conditions and N/P ratios on cellular viability were evaluated by MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) reduction assay according to the standard protocol. The cell viability was reported in terms of the control group (considered 100%) and was calculated based on the following formula: cell viability (%) = (OD of polymer-treated sample/OD of untreated sample) × 100. The data are shown as the mean value ± standard error of the mean (SEM). Each treatment was run in triplicate.
hiPSCs generation
In brief, the fourth passage of HDFs were seeded on Geltrex (Life Technologies)-coated 24-well plates the day before transfection. When HDFs reached 70–80% confluence, they were transfected by oriP/EBNA1-based vector genes presented in Figure 1 [plasmid information and sequences are available from Yu et al. (2009); Supplementary Material]. Based on the optimized protocol and N/P ratio, we prepared PEI/pDNA complex via diluting 1 μL PEI (1 mg/mL) in distilled water to the volume of 25 μL and concentration of 40 μg/mL. We also diluted the 1 μg pDNA in distilled water to reach the volume of 25 μL. The contents of the PEI tubes and pDNA tubes were mixed by gentle pipetting. The reagents were incubated at room temperature for 30 min.

oriP/EBNA1-based vector genes, including OCT4 (NM_00701), SOX2 (NM_003106), NANOG (NM_024865), LIN28 (NM_024674), c-Myc (NM_002467), and KLF4 (NM_004235).
The solution of the PEI/pDNA complexes was added to 250 μL starved culture (without FBS and Pen/Strep) and added in a dropwise manner directly to cells grown in four wells of a 24-well plate. After incubating for 4 h, the medium was replaced with fresh high glucose Dulbecco's Modified Eagle Medium (DMEM) with 10% FBS. Following this, the medium was replaced every other day. On day 4 posttransfection, the morphology of HDFs changed from spindle to rounded shape. At this time, the medium was replaced with human embryonic stem cell culture media (PluriSTEM; Millipore) containing fresh 100 ng/mL basic-fibroblast growth factor (Royan Institute). From then on, the medium was refreshed every day. HiPS colonies emerged 18–25 days after transfection.
iPSC characterization
Alkaline phosphatase staining
Alkaline phosphatase (AP) Staining Kit (Stemgent) was performed according to the manufacturer's protocol to confirm generated iPSCs. Images were captured by Olympus CKX41 microscope with an Olympus DP72 camera (magnification × 100).
Immunofluorescence staining
iPSCs colonies were fixed by 4% paraformaldehyde for 15 min at room temperature and washed three times with phosphate buffered saline (PBS). Then the cells were treated with 0.2% Triton X-100 for 15 min at room temperature and blocked using 5% bovine serum albumin and 0.1% Triton X-100 for 1.5 h at room temperature. Following removal of blocking buffer, colonies were incubated with primary antibodies against KLF4 (ab26648), OCT4 (ab18976), SOX2 (sc-365964), and cMYC (sc764) at 4°C overnight. The next day, colonies were incubated with secondary antibodies conjugated with (Alexa Fluor 488) (1:500; Invitrogen; ThermoFisher Scientific, Inc.) in the dark for 1 h at room temperature. After three washing steps, 4′,6-diamidino-2-phenylindole was applied to stain the cell nuclei. Images were captured within 30 min using Olympus BX50 fluorescent microscope.
Cytogenetic analysis (Karyotype)
The normal chromosomal constitution was verified by conventional karyotyping of generated iPSCs. Conventional cytogenetic analysis was performed on hiPSCs using conventional QFQ-banding at 450 bands resolution according to the International System for Human Cytogenetic Nomenclature. A minimum of 10 metaphase spreads were analyzed for each sample and karyotyped using a chromosome imaging analyzer software (Chromowin Software; Tesi Imaging).
Embryonic body formation
The hiPSCs were harvested using Dispase (1 mg/mL; StemCell Technologies) when they reached 80–90% confluence. Then the cells were resuspended in differentiation medium (high glucose DMEM, 2 mM
Real-Time PCR Primer Sequences
Real-time PCR analysis
Real-time PCR was performed to evaluate the expression level of pluripotency genes (oct4, sox2, nanog) to confirm generated iPSCs lines. Moreover, gata4, msx1, and sox1 genes were checked in hiPSC-derived differentiated cells to ensure trilineage differentiation (endoderm, mesoderm, and ectoderm, respectively). The real-time PCR was performed with appropriate primers for exogenous pluripotency markers and lineage-specific markers. The sequences of all primers are shown in Table 2. The relative expression level of β-actin was used for normalization.
Statistics
Statistical analyses were performed using GraphPad Prism version 8.0.1. All descriptive results were expressed as mean ± standard error of at least two to three independent experiments. p-Value of <0.05 was considered as statistically significant.
Results
Determining the best condition for transfection of HDFs
Based on our previous experiments, PEI 25 kDa at N/P ratio of 12 could efficiently transfect standard cell lines such as human embryonic kidney (HEK) cells. Thus in the current study, different experiment conditions were adjusted based on PEI/pGFP ratio of 12 at the first stand. Fluorescent microscopy, flow cytometry, and MTT assay was performed to find the most efficient condition with few cytotoxicity effects. The following parameters were set and led to find an optimized condition for PEI-mediated transfection (Fig. 2 and Table 3):
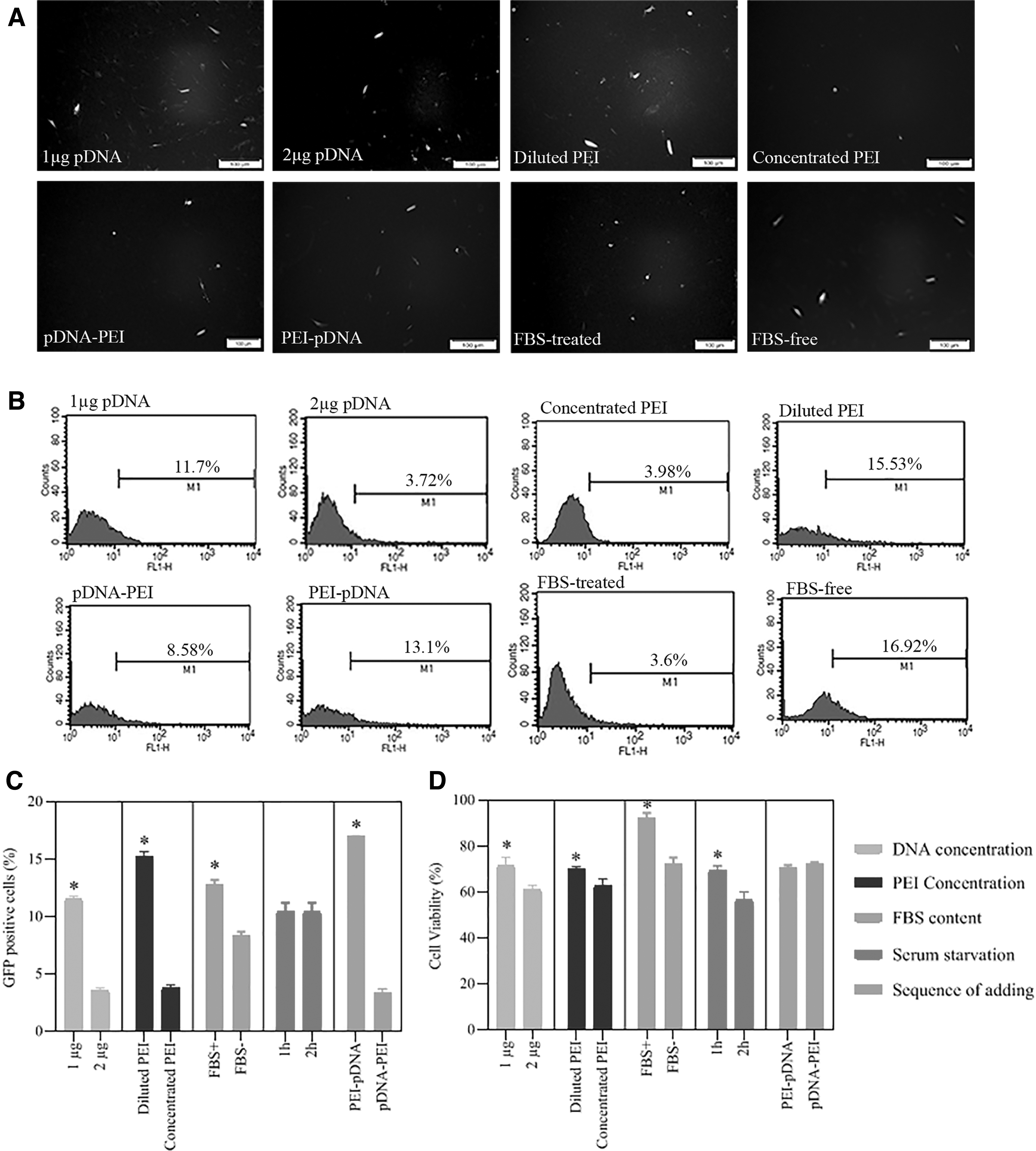
Initial optimization of PEI transfection conditions. HDF cells were assessed by fluorescence microscopy, flow cytometry, and MTT assay 48 h after transfection.
Mean Percentage of Cell Viability and Cytotoxicity of Human Dermal Fibroblast Cells Transfected with Different Conditions
P/D, PEI/pDNA; D/P, pDNA/PEI.
A significant difference (p < 0.05).
Plasmid concentration
The fluorescence microscopy and flow cytometry method showed that the PEI/pDNA complex with 1 μg pDNA efficiently transfected HDFs. In contrast, using 2 μg pDNA decreased the transfection efficiency. Moreover, the HDF samples treated by PEI/pDNA complex containing 2 μg DNA showed less than 70% cell viability.
PEI concentration
We denoted that rediluting PEI in distilled water before complex formation to reach 40 μg/mL concentration considerably increased transfection efficiency and cell viability compared to concentrated PEI (1 mg/mL).
The sequence of adding reagents
Our results showed that the order of adding reagents affects the efficiency of transfection. Adding PEI to pDNA increased the transfection efficiency, while the vice versa order decreased the rate of transfection. However, the cell viability percentage did not differ significantly between samples treated by PEI/pDNA versus pDNA/PEI complexes.
10% FBS treatment during PEI/pDNA complexation
MTT assay showed that FBS significantly reduced the cytotoxicity of PEI/pDNA complexes. However, fluorescent microscopy and flow cytometry assay determined that the presence of the FBS in polyplexes environment during complex formation dramatically decreased the transfection efficiency.
HDFs serum starvation before transfection
Based on the fluorescent microscopy and flow cytometry results, 1 and 2 h serum starvation before transfection showed similar transfection efficiency. Nevertheless, 2 h FBS-free culture before transfection significantly decreased the cell viability measured by MTT assay 48 h posttransfection.
Determining the best N/P ratio according to the adjusted condition
At the next step, the optimized condition was chosen to measure the transfection efficiency among different N/P ratios of PEI/pGFP complexes (i.e., 3, 4, 6, 8, 10, and 12). GFP-positive cells were determined using flow cytometry and fluorescent microscopy. Our results showed that the transfected cells with N/P ratio of 3 displayed the highest percentage of the cell expressing GFP (an average of ∼40%) [(Fig. 3B, C) and Supplementary Material: Supplementary Fig. S1; Lipofectamine vs. PEI at N/P ratio 3]. When we increased the ratio of PEI/pGFP, the amount of GFP-positive HDF significantly decreased in comparison to N/P ratio of 3 (p = 0.04). However, the reduction in transfection efficiency followed by increasing the N/P ratio has not occurred linearly (Fig. 3B, C). Among N/P ratios of 4, 6, 8, 10, and 12, N/P ratio of 4 achieved higher transfection efficiency (29.1%).
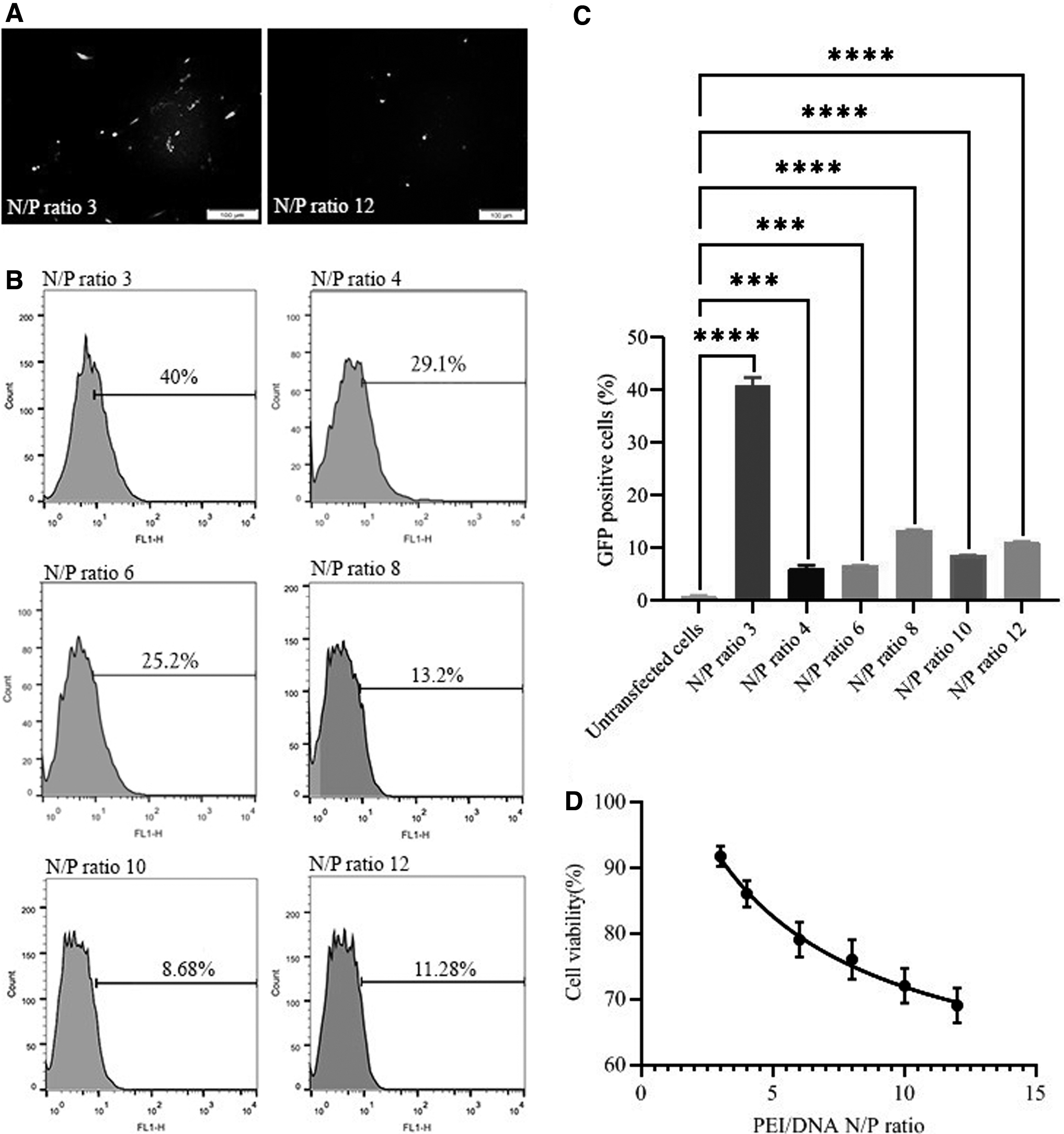
Moreover, the fluorescent microscopy results were consistent with the flow cytometer findings. Fluorescent images represented that the N/P ratio of 3 caused more GFP-expressing cells when compared to N/P ratio of 12 (Fig. 3A).
Cytotoxicity of PEI complexes
MTT assay showed that increasing N/P ratio significantly decreased the cell viability index (p = 0.001) (Fig. 3D). The cell viability of all HDF groups is represented in Table 4. Results indicated a substantially higher cell survival rate in the HDFs transfected with N/P ratio of 3 than other N/P ratios
Mean Percentage of Cell Viability and Cytotoxicity of Human Dermal Fibroblasts Transfected with Different N/P Ratios
Generation and characterization of hiPSCs
Complexes which assures a balance of considerable transfection efficiency and acceptable toxicity (Fig. 2), was chosen to further transfect HDFs to investigate the potency of this protocol to generate hiPSCs through transfection with oriP/EBNA1 vector. On day 14, posttransfection colonies emerged. Colonies exhibited hESC-like morphology with refractive edges and three-dimensional growth (Fig. 4A). An AP assay was performed to distinguish the fully reprogrammed hiPSC colonies. AP staining illustrated that 1–3% of rounded colonies expressed AP enzyme. The colonies with hESC colonies morphology could be better noticeable from non-iPS colonies upon passaging and were individually and manually picked for expansion and analysis (Fig. 4A).
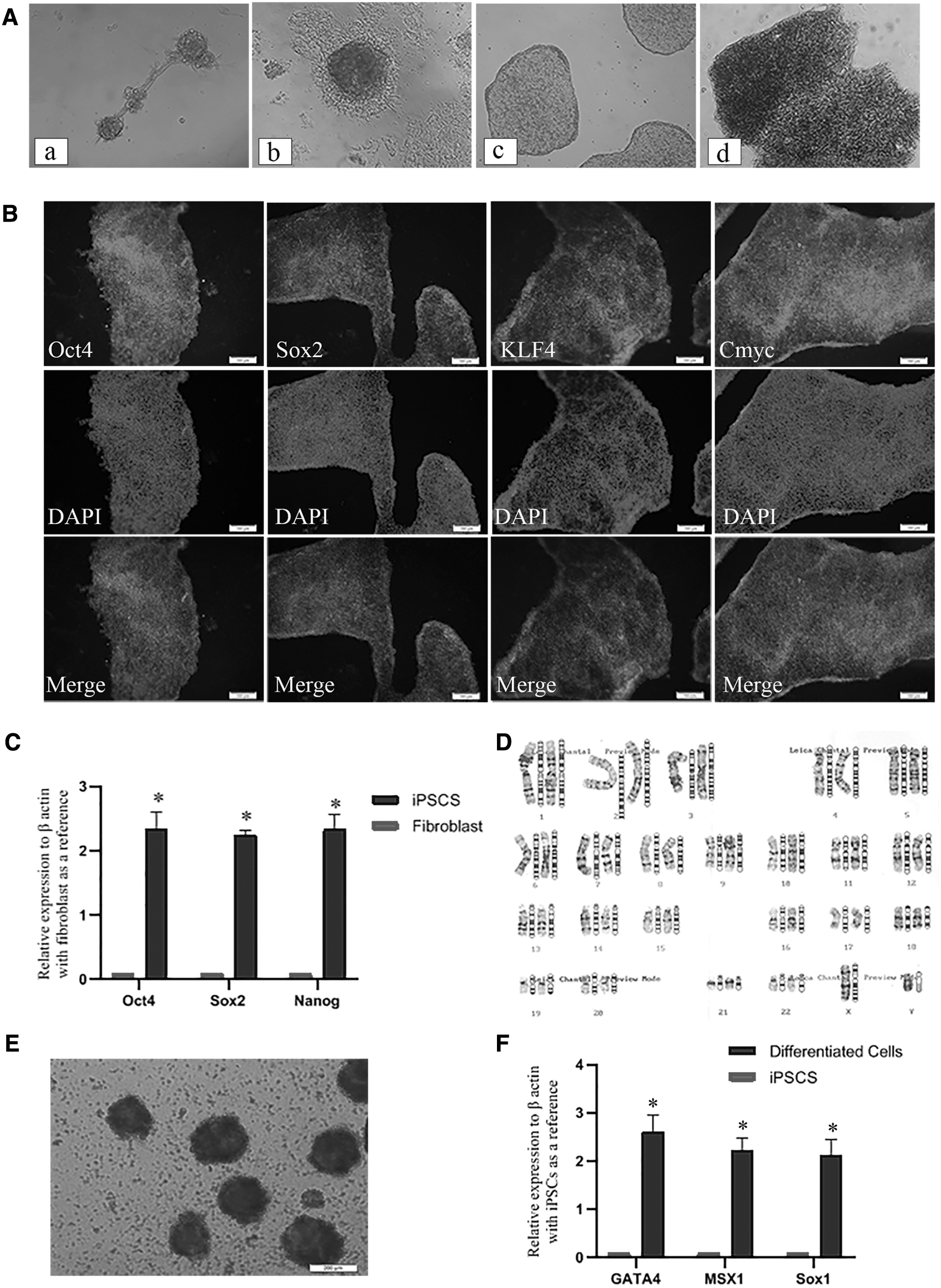
Under sterile conditions and using a 1 mL syringe, hiPSC colonies were broken into pieces on days 30–35 and transferred onto a 1% Geltrex-coated 12-well plate containing 0.5 mL PluriSTEM. Colonies were attached to the culture plate 48 h after colony transferring, and from then on, the medium was replaced every day. When the colonies covered 80–90% of the surface area of the culture plate, they were considered ready for multiple passaging to obtain hiPSC-lines ready for characterization.
Immunofluorescence staining was administrated for the expanded colonies after 8–10 passages. Immunofluorescence microscopic images displayed the expression of pluripotency markers, including KLF4, OCT4, SOX2, and cMYC (Fig. 4B). Furthermore, real-time PCR analyses showed that all chosen clones expressed the endogenous reprogramming and pluripotency factors of nanog, sox2, and oct4. Relative expression level (REL) for these pluripotency genes were markedly increased in the iPSCs compared to the parental HDFs (Fig. 4C). The genetic stability of the hiPSCs was also confirmed by karyotyping (Fig. 4D). The established hiPSC line exhibited a normal karyotype of 46 XY as confirmed by chromosomal G-band analysis at passages 8–10.
Moreover, the differentiation potential of generated hiPSCs was examined using EB formation and spontaneous differentiation (Fig. 4E). The expression of three germs layer markers, including msx1 (mesoderm), gata4 (Endoderm), and sox1 (ectoderm), was checked by real-time PCR analysis. All differentiated markers were upregulated in hiPSC-derived differentiated cells in comparison to progenitor hiPSCs (Fig. 4F).
Discussion
Although PEI is a stable, easy to handle, and inexpensive cationic polymer (Godbey et al., 1999a; Xu et al., 2021), very few studies pointed out the ability of this reagent in generating hiPSCs. Moreover, the optimized protocol required for complexation of PEI/pDNA polyplexes for generating hiPSCs cells was missing in the literature. In the current investigation, we represented a cost-effective and optimized protocol for transfecting HDFs through a gold standard reagent (i.e., PEI) and oriP/EBNA1-based vectors to produce hiPSCs. Previous evidences have confirmed the capability of PEI to transfect several types of cells such as HEK293 (Choi et al., 2016), mesenchymal (Tong et al., 2013), neural (Shakhbazau et al., 2011), and embryonic (Bartman et al., 2015) stem cells. However, with the aim of generating hiPSCs, our investigation is of the pioneers setting and standardizing several parameters relevant to HDF transfection process using PEI25K.
Regarding parameters relevant to complex formation, our results revealed that using 2 μg pDNA implies a lower transfection rate and higher cell death in comparison to 1 μg pDNA, which may be related to the aspect ratio of PEI/pDNA. Higher concentrations of pDNA require a higher amount of PEI for modification and permeabilization of endosomal membrane, which causes a higher cytotoxic effect of polyplexes on cell viability (Moghimi et al., 2005; Liang and Lam, 2012). In addition, regardless of PEI dosage, an excess of pDNA itself can also contribute to cellular toxicity (Juckem, 2012; Chopra et al., 2020).
In addition, we found that a lower concentration of PEI (dissolved in a low ionic solvent such as distilled water) could prominently increase both transfection efficiency and cell viability compared to concentrated PEI. Besides the lesser toxic effect of PEI at lower concentrations (Zintchenko et al., 2008), low-concentrated branched PEI has different formation and hydration properties, facilitating transport and diffusion of the polyplexes through cell membrane (Rudolph et al., 2000). Moreover, we hypothesized that diluted PEI might provide more free branches than a concentrated state, advancing DNA-binding affinity and improving cellular uptake (Zhou et al., 2016). In addition, diluted PEI might form a thicker layer around pDNA than concentrated PEI, which enhances the protonation state of PEI/pDNA polyplex structure and provides an electrostatic and physical barrier to DNases with cationic surfaces (Godbey et al., 1999b).
Our results also represented that adding PEI to pDNA can increase transfection efficiency relative to the converse order related, in part, to the electrostatic interactions between the positive charges of cationic polymers and negative charges of pDNA (Lucotti et al., 2014). Previous evidence have also shown that stoichiometry of both components in a binary complex and the properties of final complex depend not only on the nature of the regent or polymer (including molecular weight, charge densities, stereochemistry, etc.) but also on the preparation conditions such as ratio of mixing (N/P ratio), PH of the medium, the ionic strength, and order of mixing reagents [see Fukuda and Kikuchi, 1979; Chen et al., 2012; Hsu and Uludağ, 2012; Pezzoli et al., 2017) for some examples and a detailed description].
Pezzoli et al. (2017) showed that mixing order and even dropping mode of polyplex (PEI/pDNA) formation significantly gave rise to the formation of cationic nanoparticles with different hydrodynamic diameter (DH ), PDI, and ζ-potential. For example, the mixing order of “pDNA solution added to PEI” resulted in nanoparticles with ζP between +22 and +30 mV and DH between 143 and 161 nm. Also, PDI was invariably low (PDI <0.2) indicating a quite monodisperse particle populations in comparison to other mode of polyplex formation. It is worthy of note that different contextual conditions such as type of solvent, PH of medium, and N/P ratio could affect the aforementioned results (Chen et al., 2012; Pezzoli et al., 2017).
Given above, even subtle changes in the sequence of addition of reagents prominently influence the physiochemical properties of PEI-polyplexes (Pan et al., 2013; Pezzoli et al., 2017; Zhang et al., 2017). However, so far, there were few data regarding the order of mixing PEI/pDNA polyplexes required for generating hiPSCs form HDF cells.
Furthermore, we found that the presence of serum in polyplexes significantly decreased transfection efficiency due to the interfering role of serum in dissociating and disrupting the structure of PEI/pDNA complexes (Forcato et al., 2017). However, the MTT test showed that the existence of FBS prominently increased the cell viability, which may be attributed to the potential of serum proteins in absorbing free PEIs (Forcato et al., 2017). Previous studies also confirmed that the presence of FBS in polycations-based transfection reduces both cytotoxicity and transfection efficiency (Pezzoli et al., 2016). Therefore, in the current study, using FBS during polyplex formation was not favored by transfection efficiency and is not recommended in HDF cell type.
Apart from the impact of FBS in the structure of polyplexes, it is evident that FBS in cell culture media before transfection can affect the efficiency of PEI/pDNA delivery to the cells. Serum starvation induces cell cycle synchronization (G0/G1 proliferation delay), which eventually increases transfection efficiency (Shin et al., 2008; Tong et al., 2016). Moreover, the competitive interaction of serum proteins and polyplexes with cell surface reduces the efficiency of transfection. Herein, we examined the impact of 1 h versus 2 h serum starvation on HDFs' transfection efficiency and cell viability. We discovered that both groups showed the same transfection efficiency. Yet, 2 h serum starvation decreased cell viability in comparison to 1 h starvation. Nevertheless, the optimized serum starvation period differs in other studies depending on the studied cell types (Oya et al., 2003).
Given together, our optimized protocol consists of 1 μg pDNA, diluted PEI (40 μg/mL), PEI to pDNA addition sequence, serum-free polyplexes, and 1 h serum starvation of HDFs. Besides, since N/P ratio plays an essential role in influencing the degree of complexation, particle diameter, transfection efficiency, and cytotoxicity of carriers (Zhao et al., 2009), in the next step, we investigated the effect of N/P ratio on transfection efficiency and cell viability using the optimized protocol. According to previous documents implying that nearly all DNA is bound to PEI by ratios higher than 2 (Zhao et al., 2009; Patnaik et al., 2010), we set N/P ratio of 3 as the minimum evaluated N/P ratio. In addition, since N/P ratios higher than 12 expose cells to more toxic effects (Kang et al., 2010; Cheraghi et al., 2017), we set N/P ratio of 12 as the highest evaluated N/P ratio in our investigation.
In a novel finding, we observed that the PEI/pDNA ratio of 3:1 led to the highest transfection efficiency and cell viability in comparison to other higher N/P ratios representing the acceptable strength of low-dose PEI in successfully carrying a large-sized plasmid (∼20 kb) into HDFs. We hypothesized that increased binding ability and cytotoxic effect of PEI at higher N/P ratios may justify in part such finding.
The maximum efficacy at a low-dose N/P ratio found in this study differs from previous studies, which used higher polyplex N/P ratios to improve the transfection efficiency even at the cost of cell viability (Zhao et al., 2009; Cheraghi et al., 2017; Forcato et al., 2017; Zhang et al., 2020). For example, Cheraghi
They reported that according to Dynamic Light Scattering results, by increasing N/P ratio, polyplexes showed more positive charges, which in turn produced more transfection efficiency and less cell viability. They postulated that among ratios of 3, 6, 12, 18, and 24, N/P ratio of 12 could establish an optimized ratio between transfection efficiency and cytotoxicity of PEI/plasmid nanoparticles. In another investigation, Zhang et al. (2020) assessed the effect of extracellular nanovesicles (EVs) in PEI-mediated transfection of GFP-encoding plasmid into HEK293T cells, A549 cells, and in the zebrafish embryos (Zhang et al., 2020). They suggested a very low transfection efficiency for PEI25K and PEI60K at N/P ratios under 32.
Due to the negative charge of both EV and DNA, the authors suggested that by increasing N/P ratio higher than 32, zeta potential (surface charge of polyplexes) increases, and subsequently, transfection efficiency will enhance. However, to achieve an acceptable transfection efficiency, they also observed an unavoidable decrease in cell viability by increasing N/P ratio.
We concluded that there could be two possible explanations regarding our finding of acceptable transfection efficiency in a low N/P ratio. First, our optimization led to significant transfection even at a low N/P ratio of 3. In line with our results, evidence shows that the complex formation protocol can strongly influence transfection efficiency and cytotoxicity (Ogris et al., 1998). In other words, reagent/DNA ratio and complex formation conditions is of paramount importance for obtaining maximum efficacy with the least side effects in cell transfection process (Sou et al., 2013).
Second, in the current study, we used low ionic solvent to dissolve PEI. Evidence have shown that PEI/pDNA complexes in high ionic conditions (e.g., HEPES buffered 150 mm saline ) exhibited a strong tendency for aggregation at low N/P ratios, which can be avoided by increasing the N/P ratio to 6 or higher (Ogris et al., 1998), while polyplexes in low ionic solvents represented an opposite behavior (Ogris et al., 1998). Therefore, by administrating the optimized conditions and N/P ratio, which we presented in the current study, we circumvented issues with either transfection success or high cytotoxicity.
Finally, PEI polyplexes containing oriP/EBNA1 vector were used to generate iPSCs from HDFs through the optimized condition and N/P ratio. In this study, the efficiency of colony formation was about 2 colonies per 150,000 input cells. Although efficiency was low, it is still sufficient to establish iPSC lines from HDFs (Yu et al., 2009). In the same vein, it has been previously denoted that despite the development of numerous methods of introducing the reprogramming factors into the somatic cells, only a small percentage of cells may completely express the pluripotency factors and end up the road toward pluripotency state (Plath and Lowry, 2011). Our colonies exhibited positive iPSC characterization tests indicating that iPSC colonies and lines had been successfully developed.
However, despite the common administration of lipotransfection in the generation of iPSCs, there is evidence showing that lipofection has its own adverse effects on transfected cells (Sharma et al., 2018). For example, evidence has been demonstrated that lipid-based methods have been linked with causing cell cycle arrest, proinflammatory reactions, and increased apoptosis (Sharma et al., 2018). Although PEI exhibits cytotoxicity issues in cellular transfections, in our study, we represented that a low-dose ratio and an optimized polyplex preparation condition helped minimize polyplexes' toxicity effects on our cell type studied (i.e., HDFs).
In addition, there is evidence showing a relatively better efficacy of PEI in comparison to common lipid-based reagents in transfecting different cell types (Supplementary Material: Supplementary Fig. S1). For example, some types of cells such as pig fetal fibroblast, Chinese hamster ovary cells, and HEK293T cells have undergone better transfection administrating PEI in comparison to lipid-based reagents (e.g., Effectene and Lipofectamine) (Densmore et al., 2000; Maurisse et al., 2010; Sou et al., 2013; Hasanzadeh et al., 2021).
Penacho et al. (2009) in an investigation improved transfection of HeLa cells (an epithelial-like adherent cell line derived from human tissue) via transferrin-associated lipoplexes by means of precondensation of DNA with PEI of low molecular weights (2.7, 2.0, and 0.8 kDa) at various N/P ratios. Their results showed that cationic liposomes, in which the carried DNA was precondensed with low-weight PEI, become highly efficient in mediating transfection. Interestingly, they argued that one of the possible explanations could be that these complexes enter into the cells through different pathways (involving clathrin versus caveolin). In other words, PEI most likely takes advantage of its intrinsic biophysical properties to escape from the endosome to the cytosol (Penacho et al., 2009).
In addition, the overall manufacturing cost of cell transfection is also a crucial factor for determining the transfection method (Sou et al., 2013). We tried to show that polycations such as PEI, which has a well-documented strength in cell transfection and on the contrary has lower costs and is more accessible to laboratories, could perform comparably to other high-cost nonviral commercial reagents such as lipofectamine. Soe et al. (2013) compared three transfection reagents, including lipofectamine 2000, TransIT-PRO, and linear 25 kDa PEI, regarding cost-effectiveness and efficacy of transient expression of enhanced GFP in Chinese hamster ovary cells. They observed a higher number of mRNA copies per viable cell and higher protein titer in PEI than lipofectamine group.
They also stated that the cost of lipofectamine reagent would be the highest followed by TransIT-PRO and then PEI. Two grams of PEI supplies enough reagent for 2 L solution or 20,000 reactions (six well plate) (Bartman et al., 2015). For the same price, a very lower number of reactions (∼ 60) can be performed, if expensive reagents such as lipofectamine are used. In fact, using Lipofectamine imposes much more cost on laboratories. So, PEI could serve as a cost-effective alternative for large-scale generation of iPSCs, which can be a serious limiting factor for laboratories facing financial problems.
Despite different valuable investigations on PEI properties, to our knowledge, there was no study using PEI25K as a successful carrier for transfecting human fibroblast using oriP/EBNA1 vectors for obtaining hiPSCs. Herein, we introduce a cost-effective and straightforward method to use PEI/pDNA polyplexes in generating iPSCs. In this protocol, we overcome the cytotoxicity of PEI-mediated complexes via optimizing transfection conditions and minimizing PEI/pDNA ratio. Our method can be greatly useful in research projects, especially for laboratories with financial issues. Availability and Simplicity use of PEI reagents makes it a suitable candidate for large-scale transfection approaches.
Conclusion
In this study, we represented that PEI25K can transfect HDFs to generate hiPSCs with minimal cytotoxicity. Owing to high molecular weight (>25 kDa), PEI exhibits toxicity issues, which are, in part, due to the absorption of anionic serum proteins onto polyplex surface between cationic polymers and anionic plasma proteins of serum (Wadhwa et al., 2020). However, we assumed that a low-dose ratio and an optimized polyplex preparation condition helped minimize polyplexes' toxicity effects on our cell type studied (i.e., HDFs).
Our protocol is safer, less costly, and less complicated than conventional viral and lipid-based nonviral methods. However, we recommend future studies to control for other nonviral reagents such as lipid based reagents to better clarify the potency of PEI in generating hiPSCs from different cell sources. In addition, optimizing different other parameters relevant to the transfection process using polyplexes with the aim of hiPSCs generation, is recommended in future studies. We could propose that by setting the vital parameters, using PEI on iPSC generation will be a very important primary step toward the extension of the field. At the later phases, we can further improve and optimize PEI efficacy through PEI derivatives for iPSC generation.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by Iran University of Medical Sciences (code: 31967).
Supplementary Material
Supplementary Figure S1
Supplementary Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.