
Abstract
Psychotropic drugs have long been known to possess antimicrobial activity against several groups of microorganisms. Although this property has been extensively studied both alone and when combined with antibiotics against antimicrobial-resistant bacterial and fungal species, relatively little attention has been given to their ability to contribute to the emergence of antimicrobial resistance (AMR). We have recently reported the acquisition of multidrug resistance in Escherichia coli after exposure to gut-relevant concentrations of the antipsychotic quetiapine. Considering these observations, this review attempts to establish if a relationship between psychotropics and AMR in microorganisms has been defined in the scientific literature.
Introduction
Antimicrobial activity is observed in many drug classes with human cellular targets. Medications such as nonsteroidal anti-inflammatory drugs, proton pump inhibitors, statins, and psychotropics have shown growth inhibitory effects in microorganisms (Ait Chait et al, 2020; Imhann et al, 2015; Maier et al, 2018; Paes Leme and da Silva, 2021). Of particular interest are psychotropics, which comprise a broad category of pharmacological interventions for mental health disorders and are generally categorized as antidepressants (ADs), antipsychotics (APs), mood stabilizers, anxiolytics, or stimulants.
The relationship between psychotropics and their antimicrobial properties is well documented, dating back to 1959 when the AP chlorpromazine was discovered to have inhibitory effects on mycobacterium (Amaral et al, 2000). Since then, the use of select tricyclic antidepressants (TCAs), APs, selective serotonin uptake inhibitors (SSRIs), and benzodiazepines (BZDs) as treatment adjuncts for bacterial and fungal infections have been thoroughly reviewed (Alkhalifa et al, 2022; Butts et al, 2019; Caldara and Marmiroli, 2021; Gillard et al, 2018; Kathwate et al, 2015; Pulsen et al, 2014).
According to the National Center for Health Statistics (2022), the use of psychotropics has surged since the beginning of the COVID-19 pandemic, and today nearly one in four American adults are prescribed these medications (U.S. Census Bureau, 2022). Antimicrobial resistance (AMR) and the emergence of multidrug-resistant (MDR) organisms are a concurrent global threat. In 2019 alone, an estimated 4.95 million deaths worldwide were attributed to bacterial AMR, warranting concern for all agents with intrinsic antimicrobial activity (Murray et al, 2022). Given the widespread usage and long treatment duration, the relationship between psychotropic drugs and emergence of AMR is particularly relevant if it has been defined in the scientific literature.
In Vitro Studies
In vitro experiments have been the most prevalent source of evidence establishing a relationship between psychotropics and the emergence of MDR organisms, and through a variety of mechanisms, exposure to colonic estimated concentrations of psychotropics have led to emergence of MDR phenotypes (Supplementary Table S1).
A study by Jin et al (2018) exposed Escherichia coli to the SSRI fluoxetine for 30 days at concentrations between 5 and 100 mg/L and reported a dose- and time-dependent decrease in susceptibility for several classes of antibiotics, including fluoroquinolones, aminoglycosides, β-lactams, tetracycline, and chloramphenicol. Whole genome sequencing of fluoxetine-exposed isolates revealed chromosomal mutations in AMR genes; marR, soxR, rob, sdiA, cytR, and crp (Supplementary Table S2). Transcriptomic analysis identified increased multidrug efflux encoding genes, including acrA, acrB, and tolC and tsx.
Higher expression of the multidrug efflux pump AcrAB-TolC increase antibiotic efflux and reduces susceptibility to aminoglycosides, β-lactams, fluoroquinolones, tetracyclines, and chloramphenicol (Pradel and Pagès, 2002), suggesting that AMR among the fluoxetine-exposed isolates was triggered by the enhancement of antibiotic efflux. Fluoxetine exposure also induced higher levels of reactive oxygen species (ROS) in E. coli (Jin et al, 2018). Under duress, bacteria also activate antioxidant systems that lead to the upregulation of superoxide dismutase (sodA) and ROS-sensing transcription factors (soxR) (Imlay, 2013), which was also observed in the fluoxetine-exposed E. coli (Jin et al, 2018).
Shi et al (2021) also observed that, compared with the effects of either agent alone, concurrent treatment with the serotonin–norepinephrine reuptake inhibitor duloxetine (25 mg/L) and antibiotic chloramphenicol (4 mg/L) led to increased ROS production in E. coli and upregulation of efflux pathway genes associated with MDR phenotypes, including MlaFEDB ATP-binding cassette (ABC) transporter and AcrAB-TolC (Shi et al, 2022). Also reported were significant increases in minimal inhibitory concentrations (MICs) of 12 antibiotics, including tetracycline, cefazolin, cefoxitin, ampicillin, and chloramphenicol. One-day exposure to duloxetine alone at 100 mg/L concentration also induced resistance to cefoxitin and chloramphenicol.
We recently reported the emergence of MDR in E. coli after exposure to gut-relevant concentrations of the AP quetiapine (10 mg/L, 100 mg/L), including mutations in the multidrug resistance repressor (marR) gene (Kyono et al, 2022). MarR is a transcriptional repressor widely conserved among bacteria and is important for adaptation to environmental stressors such as antibiotics, toxic substances, and ROS (Deochand and Grove, 2017; Hao et al, 2014). When the effects of marR mutations were individually reconstructed in the parent genetic background, they did not confer the same level of AMR as isolates exposed to quetiapine for 6 weeks, despite the robust activation of the acrAB-tolC efflux pump genes (Kyono et al, 2022). Interestingly, however, the reconstructed MarR mutant strains demonstrated increased growth fitness in the quetiapine-containing media compared with the parent strain.
The AP haloperidol and BZD diazepam have also been implicated in MDR emergence. Exposure of diazepam (0.03–0.3 mM) to E. coli and Klebsiella pneumoniae resulted in increased expression of AMR transcriptional regulators, marA in E. coli, and ramA in K. pneumoniae, and decreased susceptibility to chloramphenicol, quinolones, tetracycline, and β-lactam antibiotics (Tavío et al, 2004). E. coli grown in the presence of subinhibitory concentrations of haloperidol (0.4 mM) revealed 2- to 32-fold increases in MICs to tetracycline, chloramphenicol, cefoxitin, nalidixic acid, norfloxacin, aztreonam, amikacin, and gentamicin (Tavío et al, 2012). In addition, exposure to haloperidol (0.04 mM) and diazepam (0.25 mM) were sufficient to induce expression of the soxS gene and other AMR genes, such as sdiA, acrB, acrD, acrF, and tolC, and the downregulation of the outer membrane porin, ompF (Tavío et al, 2012).
A recent study by Butts et al (2019) reported that decreased antimicrobial susceptibility from psychotropic medication was also observed in the fungal pathogen Candida albicans (Butt et al, 2019). Exposure to the AP aripiprazole (5 μM) induced the expression of the ABC efflux transporters CDR1 and CDR2, consistent with activation of the transcriptional regulator Tac1p, thereby increasing the antifungal fluconazole MICs by 32-fold.
The majority of the cited in vitro studies in bacteria report increased activation of the SoxRS system when bacteria are exposed to a wide variety of redox-cycling compounds (Fig. 1) (Gu and Imlay, 2011). Many SoxRS-induced genes encode proteins that function to exclude these compounds from the cytoplasm by processes that inhibit entry through membrane porin modifications (WaaZY, sRNA MicF) and cellular efflux (AcrAB, TolC), which can otherwise disturb enzymatic activity and DNA integrity (Gu and Imlay, 2011). Although some studies suggest endogenous ROS production from exposure to traditional antibiotics can contribute to bactericidal activity (Dwyer et al, 2014; Memar et al, 2018), this idea has been challenged (Keren et al, 2013; Liu and Imlay, 2013).
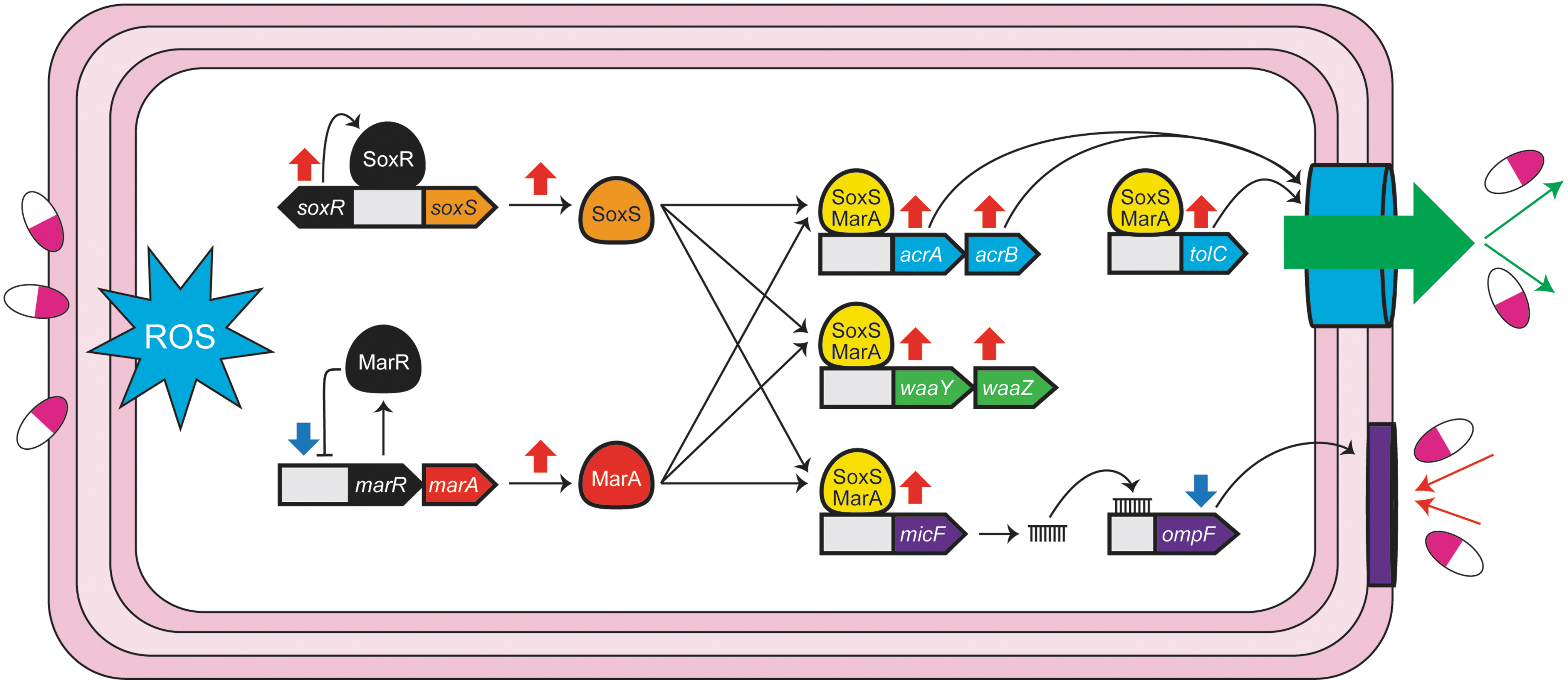
Model of psychotropic antimicrobial activity and antimicrobial resistance emergence based on available in vitro studies. ROS induced by psychotropic drug exposure leads to the activation of stress response pathways, primarily orchestrated by MarA and SoxS. This will lead to a reduced level of psychotropic drugs by increasing the efflux pump (AcrAB-TolC) to exclude the drug from the cytoplasm and also by reducing the membrane porin (OmpF) to inhibit the further drug entry into the cytoplasm. ROS, reactive oxygen species.
In psychiatry, however, ROS has been implicated in the pathophysiology of psychiatric disorders such as schizophrenia. For instance, decreased antioxidant activity has been suggested to contribute to tardive dyskinesia resulting from chronic treatment with APs (Agostinho et al, 2007). The APs olanzapine, clozapine, and haloperidol are reported to induce ROS formation in the whole blood of rats, which can be reduced with the addition of the antioxidant vitamin C (Heiser et al, 2010). Considering reports of ROS production from psychotropic exposure, the role of the endogenous ROS and activation of the SoxRS system in the antimicrobial activity of psychotropic medication should be further explored.
Animal Studies
Bacterial response to antibiotics and other medications is significantly affected by the presence of other interacting microbes, such as the gut microbiome; however, such interactions rarely considered when determining antibiotic sensitivity (Bretler et al, 2019). Although psychotropic medications are routinely associated with changes in gut microbiome composition and function (Ait Chait et al, 2020; Cussotto et al, 2019), little is known on how these changes reflect differences in AMR gene representation. One study in rats reports increased abundance of genes contributing to MDR as well as aminoglycoside and tetracycline resistance in fecal samples after oral exposure to amitriptyline (25 mg/kg/day) or fluoxetine (12 mg/kg/day) for 6 weeks (Zhang et al, 2021).
All treatment groups exhibited similar AMR gene abundance; however, enriched genes in amitriptyline-treated rats, aph3iiiA, tetM, lsA, catb2, bcR, and emrD, are reported to contribute to resistance to aminoglycosides, tetracycline, and macrolide–lincosamide–streptogramin. Indicator AMR genes abundant in fluoxetine-treated rats were categorized as multidrug (mdtK, mdtP, mdtH, mdtG, and acrA), aminoglycoside (aph3iiiA), tetracycline (tetM), and chloramphenicol (cml_e8). It is unclear whether dosing in rat models translates to humans; however, max doses for amitriptyline (150–300 mg) and fluoxetine (40–80 mg) would convert to ∼20–40 and 0.5–1 mg/kg, respectively, in an average 70 kg adult (Fluoxetine, Lilly USA, 2015; Amitriptyline, Merck & Co., 1999).
Human Studies
Psychotropics are linked to potentially adverse alterations in the human gut microbiome in psychiatric populations (Bretler et al, 2019; Cussuto et al, 2019; Flowers et al, 2017). Although the mechanism of this interplay is not defined, most gut microbes reside in the terminal ileum and colon, which is an area of detectable psychotropic concentrations (Thomas and Saadabadi, 2021; Maier et al, 2018). In a study by Maier et al (2018) reviewing the impact of nonantibiotics on gut bacteria, average colonic concentrations of human-targeted medications are estimated to be magnitude higher than whole-blood concentrations where drug levels are routinely drawn. Recent studies have started to investigate the consequence of psychotropic treatment on AMR development in human cohorts.
A 2021 study by Gashaw et al (2021) reported that E. coli isolated from stool of over 100 patients using psychotropic medications exhibited significantly higher resistance against amoxicillin-clavulanic acid, cephalosporins, meropenem, ciprofloxacin, and tetracycline than over 100 healthy controls. The odds of isolating extended spectrum beta-lactamase (ESBL)-producing Enterobacteriaceae (odds ratio [OR] = 2.3, 95% confidence interval [CI]: 1.4–4.0) and MDR organisms (OR = 5.4, 95% CI: 1.5–29.8) were also higher for psychotropic-treated patients.
A large-scale meta-analysis described the associations between prescription drug use and the gut microbiome in a cohort of healthy controls and those with inflammatory bowel disease (IBD) (Vich Vila et al, 2020). In TCA users with IBD, an increase in AMR genes were observed (n = 454; two-sided Wilcoxon-test, false discovery rate = 0.04), including statistically significant increase in the abundance of multidrug efflux pump genes tolC and mdtP. However, not all studies have positively associated psychotropics with AMR. A small study in 37 patients reported no association between psychotropic treatment and the risk of urine colonization with ESBL producing Enterobacteriaceae (Bachtarzi et al, 2019).
Translational Potential of Future Research
Accumulating evidence links psychotropics to AMR and the emergence of MDR organisms. However, due to several significant limitations in current data, the link between psychotropic drug exposure cannot be confirmed.
Many reported in vitro studies describe the development of genetic mutations or transcriptional adaptations occurring during cell replication; however, horizontal gene transfer is the primary means for AMR dissemination in bacterial communities (Liu et al, 2012; Wang et al, 2019). Epidemiological studies linking psychotropic use and infection have developed primarily in parallel to, but not interactively with, studies that examine AMR dynamics in microbial communities, creating an avenue for future research.
Only one large-scale population study links psychotropics with AMR genes in the gut microbiome (Vich Vila et al, 2020), thus limiting the generalizability of gathered data. Also diminishing data relevancy is the inconsistent pharmacokinetic consideration of relevant doses and drug concentrations impacting gut microbiota.
Psychotropic medications have several side effects that can independently contribute to increased infection risk. For instance, some cause urinary retention and sedation that can increase risk of urinary tract infections or aspiration pneumonia (Chung et al, 2016; Dzahini et al, 2018; Ferraris et al, 2021; van Strien et al, 2017). APs and ADs are also reported to depress inflammatory processes of the immune system (Eyre et al, 2016; May et al, 2019).
Despite these limitations, future study in this area may significantly impact antimicrobial stewardship in psychiatric populations and in long-term care facilities (LTCF). Infection control in psychiatric units face unique challenges due to treatment environments that encourage patient interaction. Psychiatric patients have a high incidence of chronic infection related to increased rate of substance use, inadequate follow-up, lack of insurance coverage, difficulties navigating the medical system, and socioeconomic risk factors, which may further increase interface with antibiotics (Kiraly et al, 2008). Further augmenting this risk is the increased AP prescribing for agitation in elderly patients in LTCFs with dementia (Jensen-Dahm et al, 2020; Resnick et al, 2021). Despite a concerted effort to reduce AP use within this population, an equivalent shift toward the use of mood stabilizers and ADs is reported, largely to address symptoms of dementia depression that occurs in later stages of life (Obuobi-Donkor et al, 2021; Resnick et al, 2021).
As advanced age and psychiatric comorbidities are notably associated with increased infection risk and negative outcomes from infection, MDR from psychotropic use may escalate these risks without the intention or knowledge of prescribing intensivists. Currently, little information exists offering specific recommendations to prescribers who are typically not infectious disease specialists on treating infections commonly reported in psychiatric patients. As psychotropic use continues to expand, it is critical to investigate if drug-related changes promote AMR development in microorganisms.
Footnotes
Acknowledgment
L.E. was supported by the Riback Fellowship at the University of Illinois at Chicago.
Authors' Contributions
Conceptualization, methodology, and writing—original draft by L.E. Writing—original draft and writing—review and editing by A.J. Writing—original draft, writing—review and editing, and visualization by Y.K. Conceptualization, methodology, writing—original draft, and writing—review and editing by S.F.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding information to disclose.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.