
Abstract
Hepatocellular carcinoma (HCC) is a common malignancy that is associated with poor prognosis in humans. Despite the development of targeted drugs, overall survival remains a significant challenge, and new therapeutic strategies are urgently needed. The aim of this study was to investigate the function of miR-552-5p in ferroptosis and the underlying mechanism, as well as to explore novel strategies for HCC treatment. CCK8 assay results showed that the viability of Huh-7 and Hep3B cells decreased significantly after transfection of the miR-552-5p inhibitor. In addition, we found that glutathione levels were depleted, intracellular Fe2+ levels were elevated, and the mean fluorescence intensity of C11-BODIPY was increased after miR-552-5p transfection. Transmission electron microscopy revealed that mitochondria became smaller and mitochondrial membrane intensity was increased in the inhibitor+RSL3 group. Mechanistically, a dual-luciferase reporter assay confirmed that miR-552-5p interacted with the 3′ untranslated region (3′ UTR) of acyl-CoA synthetase long-chain family member 4 (ACSL4) mRNA. qPCR and Western blotting results verified that miR-552-5p negatively regulated ACSL4 expression. In addition, we found that overexpression of ZNF8, which is a transcription factor, reduced intracellular miR-552-5p levels and enhanced sensitivity to ferroptosis. miR-552-5p reduces sensitivity to ferroptosis by targeting the 3′ UTR of ACSL4 in HCC. The ZNF8-miR-552-5p-ACSL4 axis is involved in regulation of ferroptosis in HCC, and these findings may provide a new therapeutic target for treatment of HCC.
Introduction
Hepatocellular carcinoma (HCC) is a common malignancy that is associated with poor prognosis in humans. In 2020, nearly 830180 new HCC-related deaths were recorded, accounting for 8.3% of all cancer-related deaths, and this mortality rate ranked third after those of lung cancer (18%) and colorectal cancer (9.4%) (Sung et al., 2021). The latest research predicts that the number of patients who are newly diagnosed with liver cancer will exceed 1 million by 2025.
The main risk factors for HCC are HBV, HCV, and other viral infections and alcoholic and nonalcoholic steatohepatitis (Llovet et al., 2021). Novel therapies for treating liver cancer, such as molecular targeted therapies (regorafenib, lenvatinib, and sorafenib) and antitumor immunotherapies (anti-PD1, anti-PDL1, and anti-CTLA4), have significantly improved the survival rate of patients with advanced liver cancer (Sun et al., 2022).
However, nearly 30% of patients experience sorafenib resistance within 6 months and have a poor prognosis (Tang et al., 2020). Similar results are observed after treatment with other molecular targeted drugs and immune checkpoint inhibitors, and development of drug resistance is a great challenge in the treatment of HCC (Bagchi et al., 2021). Therefore, exploring the mechanisms and seeking novel molecular targets are urgently needed.
miRNAs are a class of small noncoding RNAs that are 17–25 nucleotides long and widely expressed in animals, plants, and viruses (Fabian et al., 2010; Lee and Dutta, 2009). miRNAs are a main component of miRISCs (miRNA-induced silencing complexes), which generally inhibit protein synthesis by suppressing translation and/or by facilitating mRNA degradation (Behm-Ansmant et al., 2006).
miRNAs play vital roles in the biological processes of cancer cells, including cell cycle progression, invasive metastasis, programmed cell death, and angiogenesis (Lee and Dutta, 2009). miR-552 is a novel cancer-related microRNA with a highly conserved sequence that is located on chromosome 1p34.3, and miR-552 is found in both the cytoplasm and nucleus (Miao et al., 2016; Zou et al., 2020). miR-552-5p is highly expressed in various tumors and facilitates tumor development and progression. Overexpression of miR-552-5p inhibits the expression of WIF1 and contributes to breast cancer cell proliferation, migration, and invasion (Jiang et al., 2022).
In pancreatic cancer, Linc00261 inhibits metastasis by regulating the miR-552-5p/FOXO3 axis (Chen et al., 2020). In HCC, miR-552-5p is upregulated and its expression is inversely correlated with patient survival (Li et al., 2018). miR-552-5p directly interacts with the 3′ untranslated region (3′ UTR) of PTEN mRNA and activates the PI3-K/Akt signaling pathway, which contributes to tumorigenesis and progression of HCC. High expression of miR-552-5p mediates sorafenib resistance in HCC by suppressing apoptosis (Han et al., 2020).
Ferroptosis is an iron-dependent regulation of necrosis and its morphological features mainly include mitochondrial alterations that are observed by electron microscopy (Mou et al., 2019). In recent years, research on ferroptosis and cancer has attracted widespread attention, especially in fields focused on novel mechanisms underlying ferroptosis (Jiang et al., 2021; Wu et al., 2021). Loss of CPLX2 function enhances HCC sensitivity to sorafenib-induced ferroptosis (Li et al., 2023).
miR-214-3p increases sensitivity to ferroptosis by targeting the 3′ UTR of ATF4, and this finding has provided a novel strategy for treatment of HCC (Bai et al., 2020). In pancreatic cancer, USF2 inhibits sensitivity to ferroptosis, and this finding has provided new insights into the mechanism underlying ferroptosis regulation (Chen et al., 2022).
Previous studies have shown that miR-552-5p promotes cell invasion and metastasis and enhances sorafenib resistance by inhibiting apoptosis in hepatocellular carcinoma.
However, the function and specific mechanism of miR-552 in ferroptosis, which is a regulated form of necrosis, in HCC remain elusive. In this study, we elucidated the specific mechanism by which miR-552-5p reduces sensitivity to ferroptosis by targeting the 3′ UTR of acyl-CoA synthetase long-chain family member 4 (ACSL4). In addition, we demonstrated that ZNF8 decreased the intracellular miR-552-5p levels and increased the sensitivity to ferroptosis in HCC. Therefore, this study may reveal a new therapeutic target for treatment of hepatocellular carcinoma.
Materials and Methods
Cell culture and transfection
The Hep3B, HepG2, SK-Hep1, and Huh-7 hepatocellular carcinoma cell lines were purchased from Cellcook Biologicals Company (Guangzhou, China), and THLE-2 was obtained from Jennio Biotech Company (Guangzhou, China). The cell lines were cultured at 37°C in a 5% CO2 incubator. The ferroptosis inducers, RSL3 (1 μM) and erastin (10 μM), and inhibitors, ferrostatin-1 (Fer-1, 1 μM) and Z-VAD-FMK (10 μM), were obtained from MedChemExpress (New Jersey). Cells were analyzed after 24 h of drug treatment.
The hsa-miR-552-5p inhibitor and mimics, si-ACSL4 and oe-ZNF8 (NM_021089.3), were purchased from GenePharma (Shanghai, China). Lipofectamine 2000 reagent (Thermo Fisher) was used to enhance the efficacy of transfection, according to the manufacturer's instructions. The effect of gene silencing or overexpression was verified by three RT-qPCR experiments. The sequences were as follows: miR-552-5p mimics (sequence: 5′-GUUUAACCUUUUGCCUGUUGG-3′); inhibitor (sequence: 5′-CCAACAGGCAAAAGGUUAAAC-3′); and si-ACSL4 (sequence: 5′-GUCCAAGAGAUGAAUUAUATT-3′).
RNA extraction and RT-qPCR assay
Total RNA was extracted with the TRIzol reagent (Invitrogen). cDNA was synthesized using the HiScript III RT SuperMix for qPCR (Vazyme, Nanjing). qPCR was performed with the ChamQ Universal SYBR qPCR Master Mix (Vazyme, Nanjing) according to the manufacturer's instructions. GAPDH or U6 was used to normalize the results, and relative expression levels of target genes were calculated with the 2−ΔΔCt method.
The sequences of primers used in our experiment were as follows:
hsa-miR-552-5p sequence: 5′-CGCGTTTAACCTTTTGCCTGTTGG-3′;
ACSL4 forward: 5′-AATACCTGGACTGGGACCGA-3′, and reverse: 5′-GCTGGACTGGTCAGAGAGTG -3′;
PCBP1 forward: 5′-TCCATGACCAACAGTACCGC -3′, and reverse: 5′-TACACCCGCCTTTCCCAATC -3′;
SLC39A8 forward: 5′-CACCCATGTGAGGATCGGC -3′, and reverse: 5′-CAATAGCCAGCCCCACAAAA -3′;
ZNF8 forward: 5′-AGCAAGGGACCGAGCTATGG -3′, and reverse: 5′-GTGACATGGGATGGCTCCTCT-3′; and
GAPDH forward: 5′-GCACCGTCAAGGCTGAGAAC -3′, and reverse: 5′-TGGTGAAGACGCCAGTGGA -3′.
Western blotting analysis
Proteins were extracted using the radioimmunoprecipitation assay buffer (RIPA) lysis buffer (Beyotime, Shanghai). An appropriate amount of protein from each sample was separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and subsequently transferred to polyvinylidene difluoride membranes (Millipore, MA) at room temperature. The membranes were blocked for 2 h with 5% fat-free milk, and primary antibodies were incubated for 8 h at 4°C.
After incubation with the anti-rabbit IgG secondary antibody (1:5000; Elabscience, Wuhan), the membranes were incubated with ECL-enhanced contrast solution (Vazyme, Nanjing). The following primary antibodies were used in our experiment: anti-GAPDH (1:1000, Cell Signaling Technology), anti-GPX4 (1:1000, Abcam); anti-FTH1 (1:1000, Abcam); anti-ACSL4 (1:3000, Proteintech); and anti-ZNF8 (1:1000, Signalway antibody).
Cell Counting Kit-8 assay
Cell viability was measured with Cell Counting Kit-8 (Elabscience, Wuhan). Cells (5 × 103 cells/well) were seeded in 96-well plates. After 24 h of RSL3 treatment, the CCK-8 reagent was added to each well and cells were incubated for 2–4 h. The optical density (OD) value was measured at 450 nm by a microplate spectrophotometer (BioTek).
Colony formation assays
HCC cells (3 × 102 cells/well) were seeded in 12-well plates. Cells were cultured with a medium supplemented with 10% fetal bovine serum (FBS) for 2 weeks, and the medium was changed every 4 days. Cells were incubated with 4% paraformaldehyde for 20 min at room temperature, followed by staining with 0.1% crystal violet (Solarbio, Beijing).
FerroOrange staining
Cells were washed three times with the medium and then stained with FerroOrange working solution (Dojindo, Japan) for 30 min in the dark at 37°C. The fluorescence signal intensity was observed under a confocal microscope.
C11-BODIPY assay
C11-BODIPY (ABclonal, Wuhan) was used to assess intracellular lipid peroxidation. Cells were incubated with 50 μmol/L C11-BODIPY reagent according to the manufacturer's protocol. After trypsin digestion, cells were resuspended in phosphate-buffered saline supplemented with 5% FBS. Finally, the fluorescence signal intensity was measured using flow cytometry.
Glutathione assay
We used the glutathione assay kit (JianCheng, Nanjing) to determine the relative intracellular levels of glutathione (GSH). According to the instructions, the absorbance values of each well were measured with a microplate spectrophotometer (BioTek) at 405 nm. Relative GSH levels were calculated according to a standard curve and protein concentration.
Transmission electron microscopy analysis
Cells were harvested, an electron microscope fixative (Srevicebio, Wuhan) was added, and cells were incubated at 4°C for 24 h. Samples were fixed, dehydrated, embedded, sectioned, and stained. Morphological changes in mitochondria were observed by transmission electron microscopy (TEM) (JEM1400, Japan).
Dual-luciferase reporter assay
The mimics or NC and firefly/Renilla luciferase vector were cotransfected with the wild-type (WT) or mutant sequence of the ACSL4 mRNA 3′ UTR into Hep3B cells. The oe-ZNF8 plasmid or pcDNA3.1 and firefly/Renilla luciferase vectors with partial sequences of the WT or mutant pri-miR-552 promoter were transfected into Hep3B cells. Cells were incubated for 48 h and subsequently harvested. Luciferase activities were measured by a dual-luciferase reporter assay system (Promega Corp., Madison, WI).
Tumor xenograft assays
Four-week-old, male, BALB/c nude mice were obtained from Vital River Laboratories (Beijing, China). Twelve mice raised in pathogen-free conditions were randomly divided into three groups of four mice each. Hep3B cells (1 × 107) that had been transfected with antagomir-552, agomiR-552, or negative control were injected into the left flanks of nude mice. The mice were treated with 5 mg/mL RSL3 solution every 2 days for 2 weeks. The tumor volume was measured every 4 days with Vernier calipers and calculated as follows: 0.5 (length × width2).
After 4 weeks of observation, mice were sacrificed and photographed. Finally, the terminal weight of the tumors was determined. Mice received humane care, and the animal experiments in this study were approved by the Animal Care and Use Committee of Shandong Provincial Qianfoshan Hospital (approval no.: S725).
Bioinformatic analysis
To identify downstream targets of miR-552-5p, TargetScan and miRDB (
Statistical analysis
GraphPad Prism 8 software was used for statistical analysis of data in this study. We used Student's t test to analyze the differences between two groups, and results are presented as the mean value ± standard deviation. p < 0.05 was considered statistically significant; *p < 0.05, **p < 0.01, and ***p < 0.001.
Results
miR-552-5p negatively regulates ferroptosis in HCC
A CCK8 assay was performed to assess the HCC cell line viability after RSL3 treatment. We found that overexpression of miR-552-5p significantly enhanced cell survival. As expected, cell viability was significantly decreased after inhibitor transfection (Fig. 1A). In addition, cell viability was significantly restored after Fer-1 was induced, while the Z-VAD-FMK group did not change significantly compared with the RSL3 group (Fig. 1B). Meanwhile, we also observed no significant difference in cell viability in the Erastin group compared with the RSL3 group (Fig. 1B).

miR-552-5p is involved in regulation of ferroptosis in HCC.
Similar results were observed in the colony formation assay (Fig. 1C). These results suggest that miR-552-5p plays an important role in ferroptosis in HCC. The levels of intracellular GSH, Fe2+, and lipid peroxidation were measured to elucidate whether miR-552-5p plays a role in the ferroptosis pathway.
The results showed that the relative levels of GSH were reduced in the inhibitor+RSL3 group compared with the NC + RSL3 group and mimic+RSL3 group (Fig. 1D). We observed a significant increase in the fluorescence signal intensity after transfection with the inhibitor, according to confocal microscopy. Overexpression of miR-552-5p decreased intracellular Fe2+ levels compared with the control (Fig. 2A). Moreover, a C11-BODIPY assay was used to measure lipid peroxidation levels. Overexpression of miR-552-5p reduced lipid peroxide generation. We also discovered that intracellular lipid peroxide levels were significantly increased in the inhibitor+RSL3 group compared with other groups (Fig. 2B).
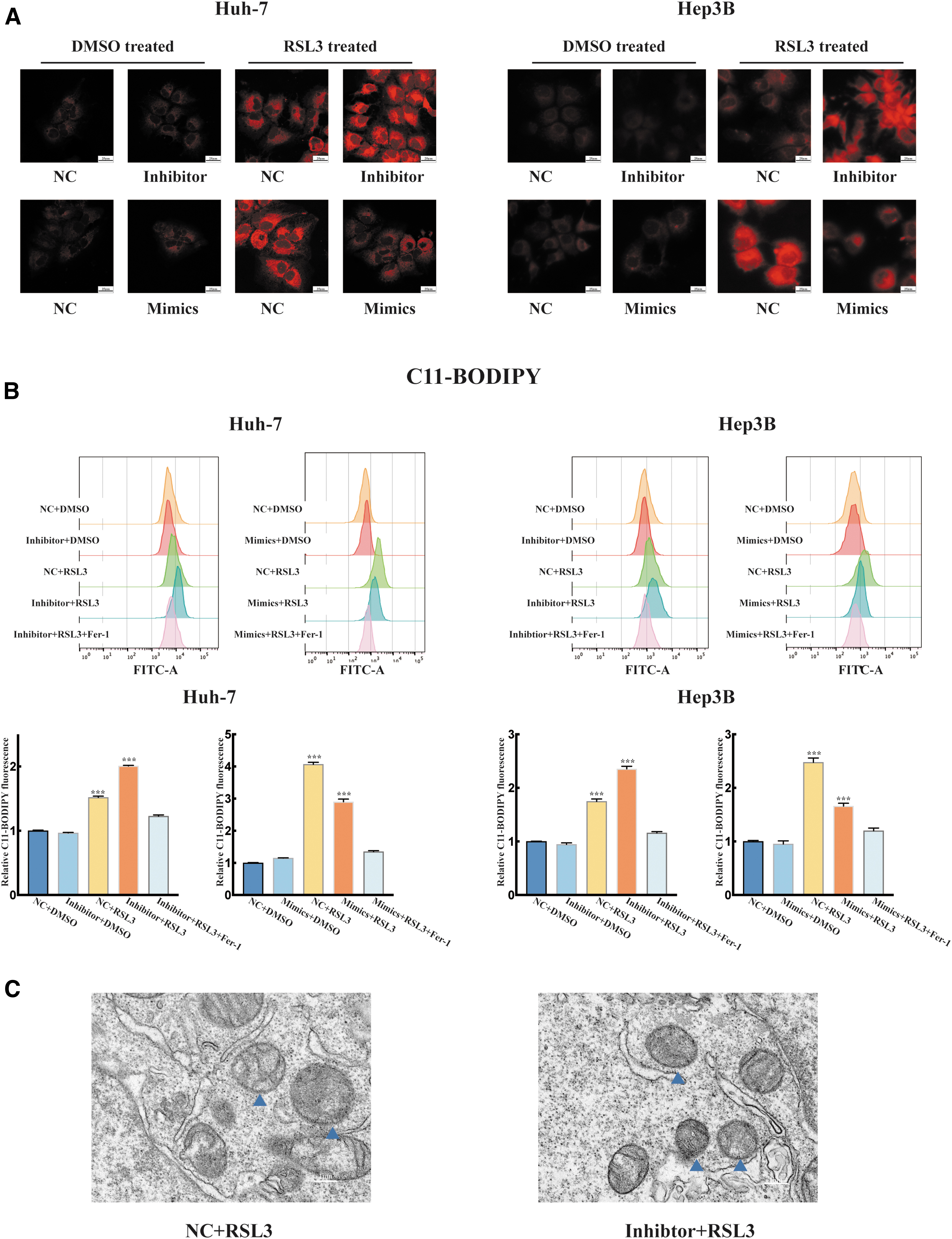
miR-552-5p suppresses sensitivity to ferroptosis in vitro.
TEM results demonstrated significant changes in mitochondria and increased membrane density in miR-552-5p knockdown cells that were treated with RSL3 (Fig. 2C).
miR-552-5p regulates ferroptosis by directly targeting the ACSL4 3′ UTR
We further explored the specific mechanism by which miR-552-5p participates in the ferroptosis pathway. As shown in Figure 3A, we predicted the downstream targets of miR-552-5p using Kyoto Encyclopedia of Genes and Genomes pathway, TargetScan, and miRDB (
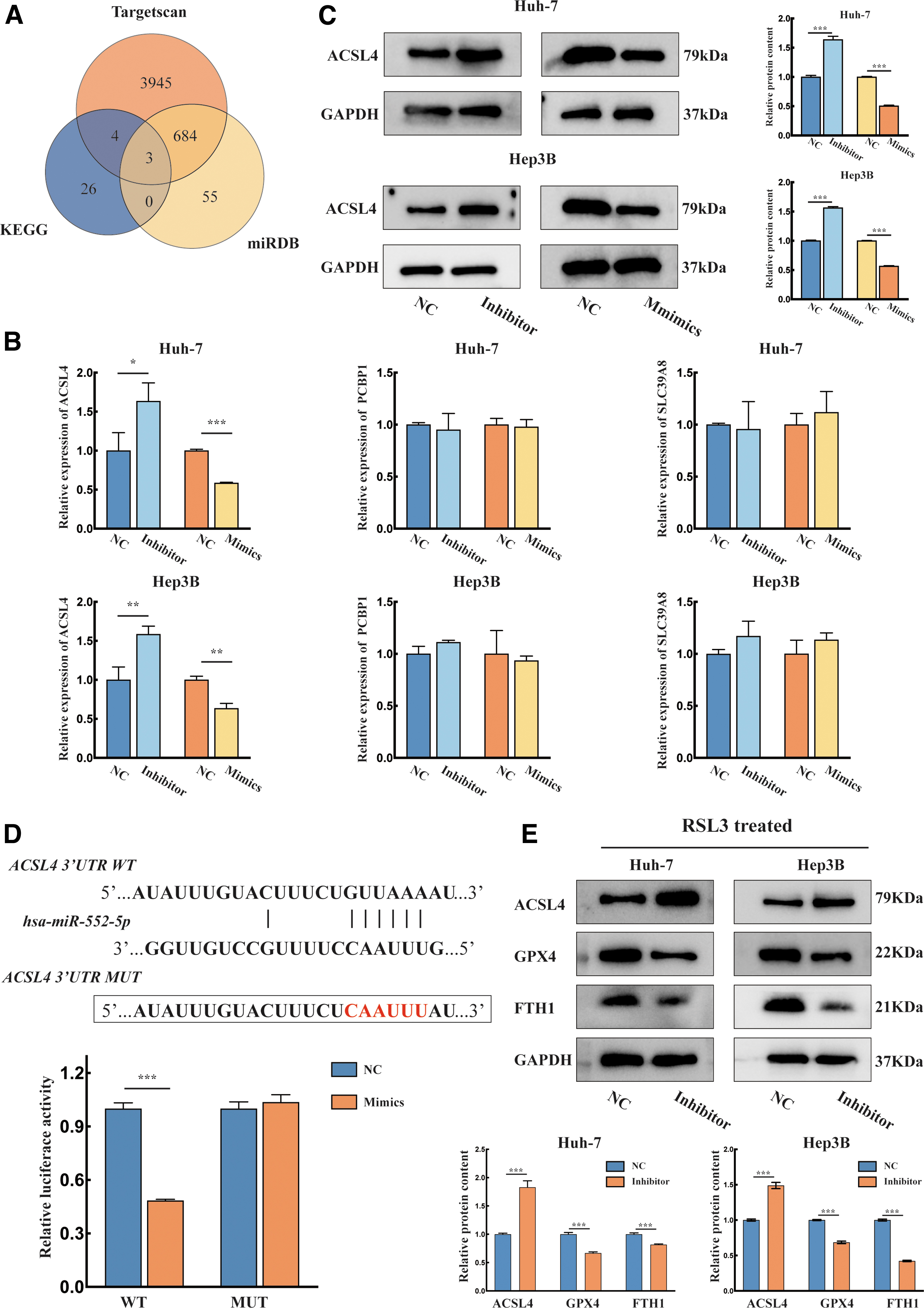
miR-552-5p interacts with the 3′ UTR of ACSL4 mRNA in HCC.
The qPCR assay revealed that miR-552-5p negatively regulated ACSL4 expression, while the levels of PCBP1 and SLC39A8 were not significantly altered (Fig. 3B). Then, we found that changes in ACSL4 protein expression were similar to the qPCR results, suggesting that ACSL4 is a target of miR-552-5p (Fig. 3C). Subsequently, dual-luciferase reporter assays were used to verify the direct interaction between miR-552-5p and ACSL4. The results showed that transfection with the mimics significantly decreased the luciferase fluorescence intensity of the WT plasmid. However, the luciferase intensity of the mutant plasmid was not significantly changed (Fig. 3D).
Western blotting assays were used to examine the key genes involved in the ferroptosis pathway. GPX4 and FTH1 expression significantly decreased after inhibitor transfection (Fig. 3E). These results suggest that miR-552-5p regulates ferroptosis in HCC by targeting the 3′ UTR of ACSL4 mRNA.
Downregulation of ACSL4 reversed miR-552-5p inhibitor-mediated ferroptosis
To elucidate whether miR-552-5p suppresses ferroptosis by regulating ACSL4 in HCC, we silenced ACSL4 and transfected the cells with the inhibitor. The CCK8 results showed that transfection with the miR-552 inhibitor facilitated the effects of RSL3 on cell viability, whereas the effect was significantly restored when ACSL4 was silenced (Fig. 4A). The intracellular GSH levels were reduced in the inhibitor group compared with the si-ACSL4 and inhibitor cotransfected group (Fig. 4B).
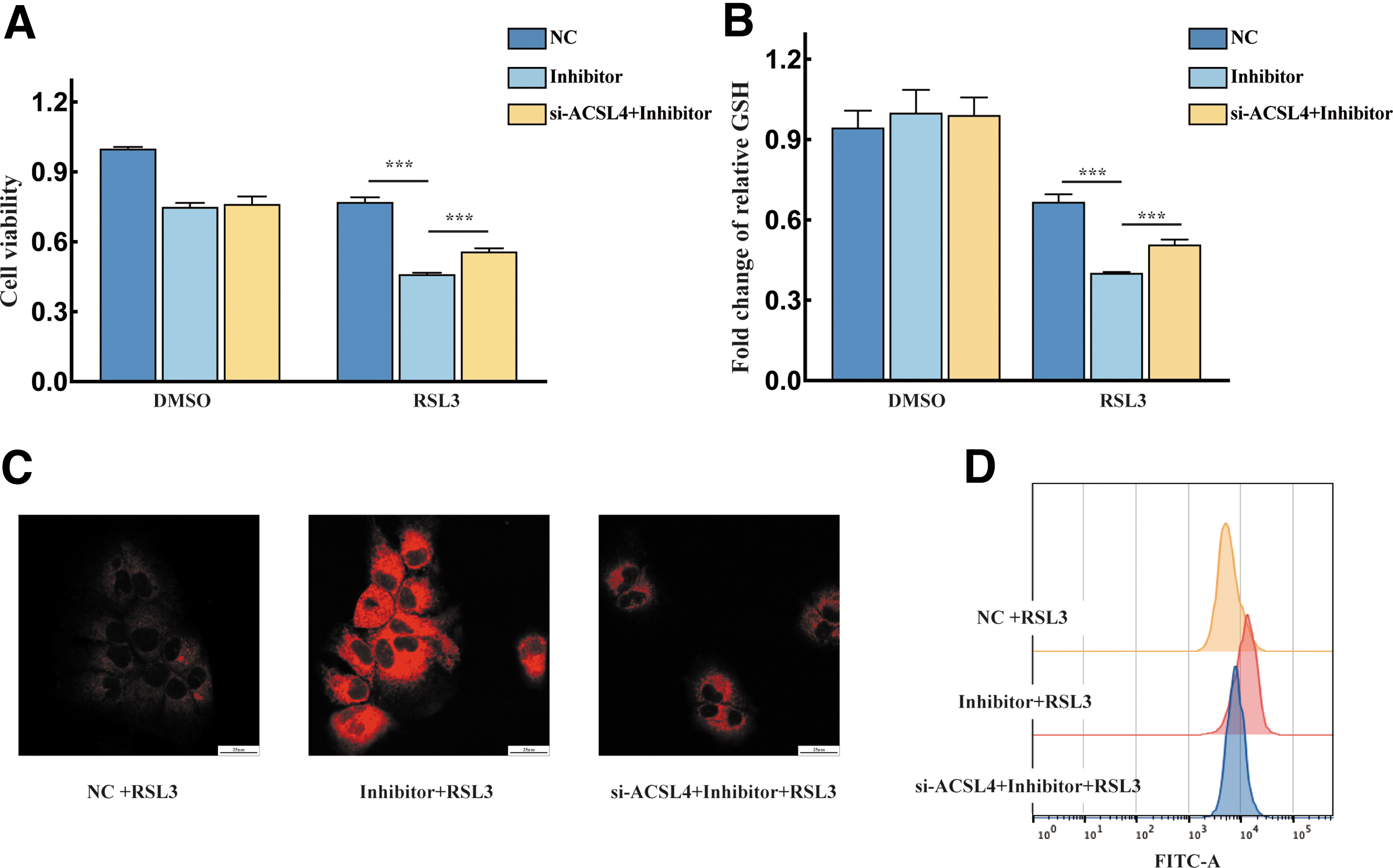
Loss of ACSL4 function suppressed the effect of the miR-552-5p inhibitor on ferroptosis.
The FerroOrange staining assay revealed that the highest intracellular Fe2+ levels were observed in the inhibitor+RSL3 group (Fig. 4C). The C11-BODIPY results showed that lipid peroxidation was restored by si-ACSL4 transfection (Fig. 4D). All these data showed that miR-552-5p negatively regulates ferroptosis by targeting ACSL4.
Inhibition of miR-552-5p contributes to sensitivity to ferroptosis in vivo
To evaluate the inhibitory effect of miR-552-5p on the ferroptosis pathway in vivo, we established a subcutaneous xenograft model. BALB/c nude mice were randomly divided into three groups: the NC+RSL3 group, agomir+RSL3 group, and antagomir+RSL3 group (Fig. 5A). Our results showed that the terminal tumor weight was significantly increased in the agomir+RSL3 group compared with other groups (Fig. 5B).

Biological functions of miR-552-5p in vivo.
Tumor volumes were measured every 4 days, and a clear trend was observed: the mean tumor volume increased significantly in the agomir+RSL3 group, but decreased in the antagomir+RSL3 group compared with the NC+RSL3 group (Fig. 5C).
This evidence further showed that inhibition of miR-552-5p promotes ferroptosis in vivo.
ZNF8 downregulates miR-552-5p and enhances sensitivity to ferroptosis in HCC
Transcription factors were predicted using the UCSC Genome Browser database, and a score >600 was used as the screening criterion; ZNF8 had the highest score (Fig. 6A). Subsequently, we performed an intergenic correlation analysis with the GEO dataset (GSE57555) and found that ZNF8 had a negative correlation with miR-552-5p.
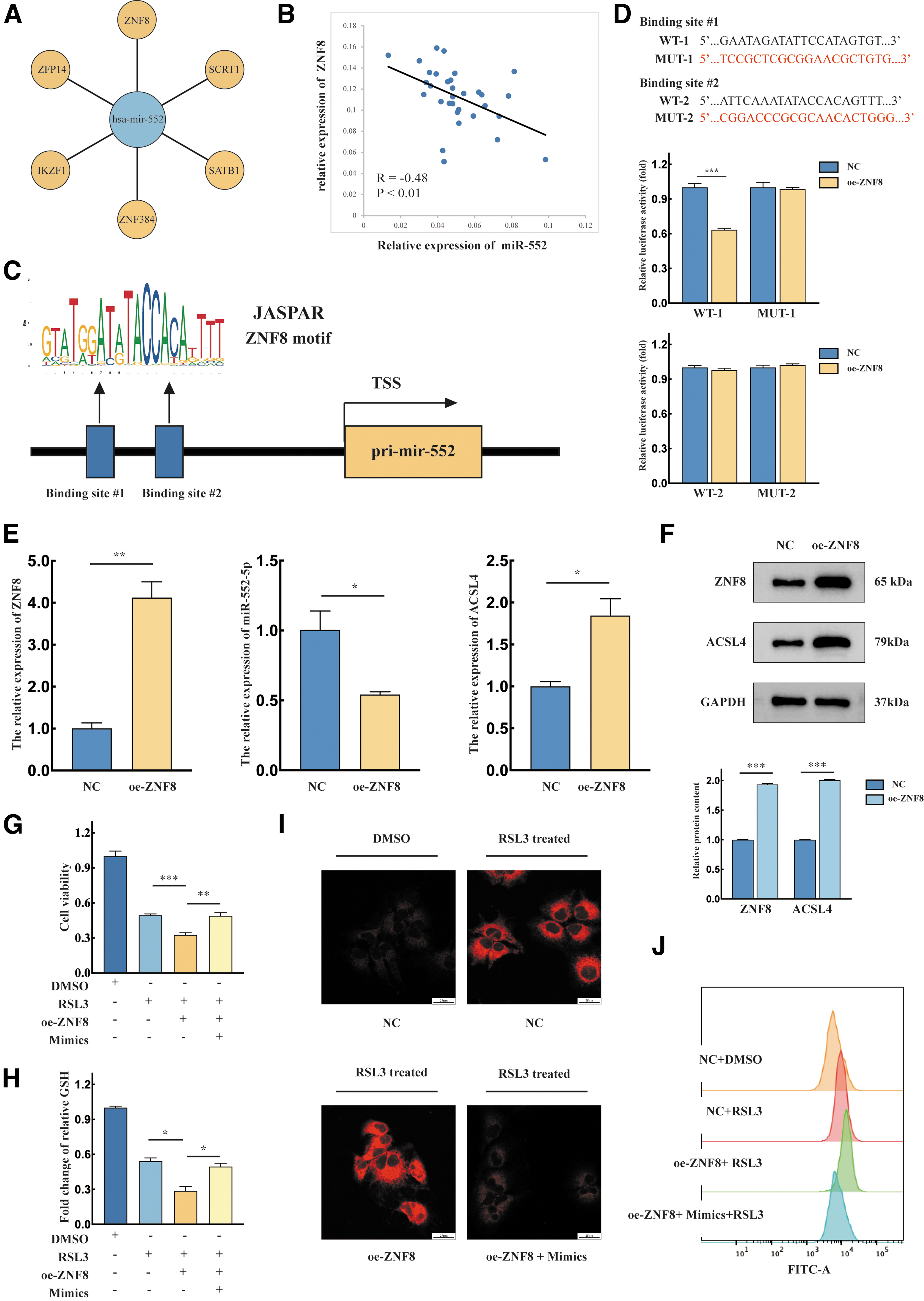
ZNF8 inhibits pri-miR-552 transcription and enhances sensitivity to ferroptosis.
Among the many transcription factors, ZNF8 showed the most significant correlation coefficient (R value) (Fig. 6B). Subsequently, we investigated the ZNF8 gene expression. The levels of ZNF8 in Huh-7 and Hep3B were elevated compared with normal liver cells (THLE-2) (Supplementary Fig. S1). To explore the specific site at which ZNF8 binds to the pri-miR-552 promoter region, we performed predictions using the JASPAR database and verified these findings with a dual-luciferase reporter assay (Fig. 6C).
The relative luciferase activity of WT 1 decreased, while that of mutant 1 was not significantly altered. In addition, the fluorescence intensity of neither WT 2 nor mutant 2 was significantly different after overexpression of ZNF8 (Fig. 6D). Western blotting and qPCR were performed to verify the function of ZNF8 in regulating miR-552-5p. We found that miR-552-5p decreased significantly and ACSL4 expression was increased (Fig. 6E). Western blotting results revealed that ACSL4 increased after oe-ZNF8 transfection (Fig. 6F).
Next, we investigated the function of ZNF8 in ferroptosis in Huh-7 cells. A CCK8 assay was performed, and results showed that the viability of ZNF8-overexpressing cells was decreased after RSL3 treatment. However, the cell viability recovered after cotransfection with the mimics and oe-ZNF8 plasmid (Fig. 6G). GSH assays revealed that overexpression of miR-552-5p reversed GSH depletion (Fig. 6H). When FerroOrange probe staining was examined by confocal microscopy, we observed the highest fluorescence intensity in the oe-ZNF8 group (Fig. 6I). A similar trend was also observed in the C11-BODIPY assay (Fig. 6J).
In conclusion, these collective results demonstrated that ZNF8 regulates the expression of ACSL4 by repressing miR-552-5p transcription and enhances sensitivity to ferroptosis in HCC.
Discussion
Ferroptosis, which was first elucidated by Stockwell in 2012, is recognized as an iron-dependent form of nonapoptotic cell death (Mou et al., 2019; Stockwell et al., 2017). MicroRNAs are implicated in different biological processes, including proliferation; apoptosis; and metabolic, transcriptional, and post-transcriptional gene regulation. Recently, studies have proven that miRNAs also play an important role in ferroptosis (Xie and Guo, 2021; Zuo et al., 2022).
The upregulation of miR-127-5p inhibits JAM3 expression in meningiomas by increasing intracellular reactive oxygen species (ROS) levels and Fe2+ cation concentrations (Zhang et al., 2022). Overexpression of miR-494-3P induces ROS production and promotes ferroptosis, leading to damage to the substantia nigra and striatum (Ma et al., 2022). In Parkinson's disease, miR-150-5p regulates intracellular SLC7A11 expression and alleviates ferroptosis by binding to BAP1 (Zhao et al., 2022). miR-147a binds directly to the 3′ untranslated regions (3′ UTRs) of its target gene SLC40A1, inhibits iron export through ferroportin, and promotes ferroptosis (Xu et al., 2022).
Several previous studies have reported that miR-552-5p expression is upregulated in HCC cells and strongly correlated with HCC patient survival (Li et al., 2018). miR-552-5p, which is an oncogene, plays an important role in promoting liver tumor-initiating cells. miR-552-5p is involved in regulation of SF-induced apoptosis (Han et al., 2020).
Our study demonstrates for the first time that miR-552-5p is also involved in regulation of ferroptosis, which is a regulated form of programmed cell death that is distinct from apoptosis. Cell viability was significantly decreased when we transfected cells with the miR-552-5p inhibitor. When miR-552-5p expression was upregulated, the opposite effect on cell viability was observed.
In addition, we observed that cells treated with RSL3 showed a rapid accumulation of intracellular Fe2+ and lipid peroxides after inhibition of miR-552-5p, which was accompanied by a significant decrease in GSH levels. More direct evidence was obtained by TEM, which showed that the mitochondrial volume decreased and membrane density increased. Collectively, this evidence reveals a novel function of miR-552-5p in inhibiting HCC sensitivity to ferroptosis.
ACSL4, which is located on the X chromosome, is the founding member of the five-member ACSL family (Quan et al., 2021). ACSL4 prefers polyunsaturated fatty acids (PUFAs), such as arachidonic acid and eicosapentaenoic acid, as substitutes to form CoA-PUFAs (Doll et al., 2017; Quan et al., 2021).
ACSL4 promotes PUFA phosphatidylethanolamine (PUFA-PE) synthesis in the cell membrane. Subsequently, LOXs directly oxidize PUFAs and disrupt the structural integrity of biofilms (Latunde-Dada, 2017; Mou et al., 2019). ACSL4 is a key positive regulator of ferroptosis. Therefore, targeting ACSL4 is a promising direction for development of a new class of cancer therapeutics.
Mechanistically, we demonstrated for the first time that miR-552-5p interacts with the ACSL4 3′ UTR through a dual-luciferase reporter assay. Silencing of miR-552-5p resulted in upregulation of ACSL4 at both the protein and mRNA levels. miR-552-5p inhibited PUFA-PE synthesis and reduced the sensitivity to ferroptosis by regulating ACSL4 expression in HCC. In addition, intracellular GPX4 and FTH1 expression was decreased by inhibitor transfection, which revealed the function of miR-552-5p in suppressing the ferroptosis pathway.
Wang et al. reported that miR-552 localizes to both the cytoplasm and nucleus. miR-552 effectively suppresses CYP2E1 expression at both the transcriptional and post-transcriptional levels (Miao et al., 2016). However, it has not been elucidated whether miR-552 regulates ACSL4 expression through transcriptional or post-transcriptional mechanisms, and this question needs to be further explored.
Zinc finger proteins (ZFPs), which are transcription factors, are involved in various biological functions such as lipid metabolism, immune response, and cell differentiation. Different species of ZFPs have distinctive DNA binding motifs (Li et al., 2022). ZFPs regulate the transcription of genes by zinc finger domain recognition or direct binding to the promoter regions of target genes.
ZNF8 is located on the long arm of chromosome 20 q13 and is ubiquitously expressed in human cells (Lania et al., 1990). Kai Jiao et al. (2002) discovered that ZNF8 negatively regulates the BMP signaling pathway during spermatogenesis. ZNF8, which is a transcriptional enhancer, facilitates the expression of ENTPD3-AS1 in renal cell carcinoma (Wang et al., 2021).
In this study, the genomic correlation analysis showed that ZNF8 was negatively correlated with miR-552-5p expression. Our study confirmed that ZNF8, which is a transcriptional repressor, downregulated miR-552-5p expression and enhanced intracellular ACSL4 expression. Moreover, we verified the function of ZNF8 in HCC. We observed that the intracellular Fe2+ and lipid peroxide levels increased and GSH levels decreased after overexpression of ZNF8.
Additionally, the rescue experiment proved that the phenotypes could be reversed by transfection with the miR-552-5p mimics. Thus, ZNF8 enhances sensitivity to ferroptosis by repressing miR-552 transcription.
Conclusions
In summary, we demonstrated for the first time that miR-552-5p inhibited the sensitivity of HCC to ferroptosis by interacting with the 3′ UTR of ACSL4 mRNA. We verified the function of ZNF8, which is involved in inhibiting the transcription of miR-552-5p (Fig. 7). The ZNF8-miR-552-5p-ACSL4 axis is involved in regulation of ferroptosis in HCC.

Schematic illustration of the mechanism by which the ZNF8-miR-552 axis regulates ACSL4-mediated ferroptosis. ZNF8, which is a transcriptional repressor, downregulated miR-552-5p expression. miR-552-5p inhibited the sensitivity of HCC to ferroptosis by targeting with the 3′ UTR of ACSL4 mRNA. AA, arachidonoyl; AdA, adrenoyl; LPCAT3, lysophosphatidylcholine acyltransferase 3; PE, phosphatidylethanolamine.
In addition, ferroptosis-based therapeutic approaches are being developed. We will further investigate whether selective knockdown of miR-552-5p combined with iron nanoparticles or other drugs that induce ferroptosis (a therapeutic approach based on ferroptosis) is effective.
Footnotes
Authors' Contributions
H.Y. designed the experiments, performed the experiments, analyzed the data and made the figures, revised the article, and wrote the initial draft. W.S. performed the experiments, revision of the article, and visualization. T.B. and J.S. participated in the investigation, resources, and data curation. Z.L., J.L., and H.W. were involved in project administration, funding acquisition, and supervision. All the authors have read the final article and support its publication.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (grant nos. 82172830, 81802414, and 82002504) and Natural Science Foundation of Shandong Province (grant no. ZR2020QH228).
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.