
Abstract
Circular RNAs (circRNAs) are generally formed by the back-splicing of precursor mRNA. Increasing evidence implicates the important role of circRNAs in cardiovascular diseases. However, the role of circ-insulin-like growth factor 1 receptor (circIGF1R) in cardiomyocyte (CM) proliferation remains unclear. Here, we investigated the potential role of the circIGF1R in the proliferation of CMs. We found that circIGF1R expression in heart tissues and primary CMs from adult mice was significantly lower than that in neonatal mice at postnatal 1 day (p1). Increased circIGF1R expression was detected in the injured neonatal heart at 0.5 and 1 days post-resection. circIGF1R knockdown significantly decreased the proliferation of primary CMs. Combined prediction software, luciferase reporter gene analysis, and quantitative real time-PCR (qPCR) revealed that circIGF1R interacted with miR-362-5p. A significant increase in miR-362-5p expression was detected in the adult heart compared with that in the neonatal heart. Further, heart injury significantly decreased the expression of miR-362-5p in neonatal mice. Treatment with miR-362-5p mimics significantly suppressed the proliferation of primary CMs, whereas knockdown of miR-362-5p promoted the CMs proliferation. Meanwhile, miR-362-5p silencing can rescue the proliferation inhibition of CMs induced by circIGF1R knockdown. Target prediction and qPCR validation revealed that miR-362-5p significantly inhibited the expression of Phf3 in primary CMs. In addition, decreased Phf3 expression was detected in adult hearts compared with neonatal hearts. Consistently, increased Phf3 expression was detected in injured neonatal hearts compared with that in sham hearts. Knockdown of Phf3 markedly repressed CMs proliferation. Taken together, these findings suggest that circIGF1R might contribute to cardiomyocyte proliferation by promoting Pfh3 expression by sponging miR-362-5p and provide an important experimental basis for the regulation of heart regeneration.
Introduction
Adult mammals have limited ability to regenerate in injured heart tissue, which leads to heart failure that is the main morbidity and mortality worldwide (Bergmann et al., 2009; Bui et al., 2011; Hsieh et al., 2007; Narula et al., 1996). However, the neonatal mouse heart can complete cardiac regeneration on apical resection at postnatal day 1 (p1) (Hsieh et al., 2007; Porrello et al., 2011), but this capacity will be lost at p7, which means the regenerative potential of the heart will be lost along with the development and maturation of the heart (Porrello et al., 2013; Porrello et al., 2011). Therefore, it is very important to study the mechanism of the regeneration capacity of neonatal mouse hearts and find a new target to overcome heart failure.
CircRNAs are covalently closed single-stranded loop structures that are produced by the back-splicing of eukaryotic precursor mRNA (pre-mRNAs) during transcription. Unlike canonical linear splicing, the back-splicing promotes a downstream splice-donor site to covalently link to an upstream splice-acceptor site, thus forming a ring structure with a 3′,5′-phosphodiester bond. CircRNAs are generally more stable than their linear counterparts, which is attributed to circRNAs having neither 5′ 3′polarities nor polyadenylated tails.
Thus, the median half-life of CircRNAs ranges from 18.8 to 23.7 h (Enuka et al., 2016). CircRNAs were initially detected by Sanger et al. (1976) more than 50 years ago, but the functional characterization of circRNAs is still elusive. Until recent years, the biological functions of circRNAs were deeply uncovered with the advancement of RNA deep sequence and ribosomal RNA depletion technology (Salzman et al., 2012).
Accumulating evidence has shown that circRNAs play important roles in a variety of biological processes, such as sponge of microRNA (miRNA), regulating transcription, and acting as protein binding partners. Most of the circRNAs are generally found in the cytoplasm and expressed in a tissue- and cell-specific manner.
To date, researchers have demonstrated that circRNAs are implicated in cardiovascular diseases (Li et al., 2020; Li et al., 2018; Viereck and Thum, 2017; Wu et al., 2021b; Zeng et al., 2021; Zhang and Ren, 2016). It was reported that circRNA_000203 can promote aggravating cardiac hypertrophy by sponging miR-2b-5p and miR-140-3p (Li et al., 2020). A previous study reported that circMAP3K5 could act as a sponge for miR-22-3p to inhibit intimal hyperplasia by suppressing SMCs (human coronary artery smooth muscle cells) (Zeng et al., 2021).
But whether circRNA could regulate heart regeneration has yet to be reported. Recently, an RNA-sequencing revealed that circRNAs are differentially expressed between neonatal and adult hearts, and some abnormal circRNAs may be involved in heart regeneration (Werfel et al., 2016). In this study, we aimed at identifying hub circRNA that could promote cardiomyocyte proliferation. Fortunately, we found a circular RNA, mmu_circ_0001582 (ID from circBase) named circIGF1R that acted as an miR-362-5p sponge to stimulate Phf3 expression to influence cardiomyocyte proliferation. Thus, circIGF1R may act as a potential biological target to combat heart failure.
Materials and Methods
Animals
Neonatal mice (C57BL/6) were used for heart apical resection, as previously described (Wu et al., 2021a). In brief, after anesthesia of neonatal mice, the position between the four and five costal was cut open, following skin incision. The apex of the ventricular chamber was resected by iridectomy scissors. After apical resection, skin and thoracic wall incisions were sutured with 7-0 non-absorbable silk sutures. Sham-operated mice underwent the same procedure without apical resection. Then, all mice were placed on the heating pad until woke up.
The total number of neonatal mice used in this study was 88, and the total number of adult mice used in this study was 6. Neonatal mice were randomly divided into two groups, each of which included three mice: the sham-operated group (Sham) and the apical resection group (dpr). Animal experiment protocols were approved by the Institutional Animal Care and Use Committee of Jinan University (IACUC-2018921-03).
Neonatal mouse ventricular cardiomyocytes and fibroblasts isolation
Primary cardiomyocytes (CMs) were isolated from neonatal mice at p1, as previously described (Wu et al., 2021a). In brief, after separating ventricles from their atria, ventricles were cleaned and cut into pieces, then digested in 0.25% trypsin at 4°C for 12–16 h. Digestions were transferred in 0.075% collagenase and repeated three times at 37°C for 15 mins. Next, the cell suspension was passed through a 70-mm cell strainer and centrifuged at 100 g for 5 mins. After removing the supernatant, all cells were mixed with culture medium and transferred to the plastic dishes for 120 min at 37°C and 5% carbon dioxide [CO2] humidified atmosphere for differential velocity adherent. At last, the supernatant was mostly composed of CMs about 90% and the adherent cells were mostly composed of cardiac fibroblasts (CFs).
Cell culture and transfection
HL-1 (a cardiac muscle cell line) purchased from ATCC (Manassas, VA, USA) was cultured in DMEM containing 10% (v/v) FBS, 100 U/ml penicillin, and 100 μg/ml streptomycin (all from Gibco, Carlsbad, CA, USA), at 37°C in a 5% CO2 incubator. The CMs were cultured in DMEN: M199 (3:1) with 10% (v/v) FBS, 5% BHIH, 100 U/mL penicillin, and 100 μg/mL streptomycin. For specific knockdown of circIGF1R and overexpression of miRNAs, siRNA targeting circIGF1R (si-circIGF1R), Phf3 (si-Phf3), and miRNA mimics (RiboBio, Guangzhou, China) were transfected into cells with Lipofectamine 3000 (Life Technologies) according to the manufacturer's instructions. The sequence of si-circIGF1R used in this study is as follows: 5′-GAA AAT CTG TGG GCC CGG CAT-3′. The sequence of mimics of miRNA used in this study is as follows: mimic-miR362-5p, 3′-UUA GGA ACC UUG GAU CCA CAC UUA-5.
Isolation of RNA for quantitative real time-PCR
Total RNA was isolated from cardiac tissues and CMs using TRI Reagent (Molecular Research Center Inc., USA). ReverTra Ace® qPCR RT Kit with random primers (Toyobo, Japan) was used for cDNA synthesized from total RNA (1 μg). The cDNA synthesis of miRNA was used in the miRNA first-strand cDNA synthesis kit (AgBio, China). The quantitative real time-PCR (qPCR) was performed by using QuantiTect SYBR Green PCR Master Mix (Qiagen GmbH, Hilden, Germany) and the MiniOpticon Real-Time PCR System (Bio-Rad, CA, USA).
GAPDH and U6 were respectively used as the internal standard control to normalize gene expression using the ΔΔCt method for mRNA and miRNA. The number of replicate experiments performed with murine tissue Primer sequences of qPCR used in this study is listed in Supplementary Tables.
Dual-luciferase assays
Cells were seeded in 24-well plates and cultured at about 60% confluence before transfection. To construct wild-type or mutant circIGF1R (546 bp) fragments of pmirGLO plasmids (MiaoLingBio, China), the 500 ng plasmids (wt-circIGF1 and mut-circIGF1R) and 50 nM mimics of mmu-miR-362-5p, mmu-miR-500-5P, and mmu-miR-670-3p were transfected into cells by lipofectamine 3000 (Invitrogen, USA). The Dual-Luciferase Reporter Gene Assay Kit (Yeasen Biotechnology, Shanghai) was used to detect the signals.
Briefly, 200 μL cell lysis buffer was added to cells and incubated for 5 mins on ice. After collecting lysis solutions, 10 μL supernatant was added to 96-well plates for detecting signals of firefly luciferase activities and Renilla luciferase activities separately using Firefly luciferin enzyme reaction solution and Renilla luciferin enzyme reaction solution. The multimode reader (CLARIOstar® Plu, Germany) was used to collect signals. At last, the differences between a firefly and Renilla luciferase activities were calculated to determine relative luciferase activity.
Immunofluorescent staining
The CMs were fixed in 4% paraformaldehyde for 30 min at room temperature, followed by 0.1% Triton X-100 treatment for 10 min, washing three times in PBS (Servicebio, Wuhan, China). EdU (RiboBio) dye liquor was added to cells (1:1000) for incubating for 2 h in a CO2 incubator before paraformaldehyde. Sections were incubated with 5% BSA at room temperature for 30 min. Sections were then incubated overnight at 4°C with primary antibodies against Ki67 (No. 9129; 1:300 dilution; CST), phosphohistone H3 (pH3) (No. 3377; 1:500 dilution; CST), and α-Actin (SAB3300072; 1:300 dilution; Sigma) respectively.
After washing three times in PBS, the sections were incubated with an anti-mouse secondary antibody conjugated to Alexa Fluor 488 (Jackson Immuno Research) and with an anti-rabbit secondary antibody conjugated to Cy3 (Proteintech Group, USA) for 1 h at room temperature to identify the proportion of pH3+ α-Actin+ , EdU(5-ethynyl-2-deoxyuridine)+ α-Actin+ , and Ki67+ α-Actin+ CMs. Next, sections were counterstained with DPAI (Sigma-Aldrich, USA). Images were captured by a laser-scanning confocal microscope (LSM 700, Zeiss). All experiments were performed in triplicate and repeated at least three times.
Fluorescence in situ hybridization
The circIGF1R probe was designed and synthesized by RiboBio. The sequence of the circIGF1R probe specifically targeted the head-to-tail junction position of circIGF1R. The fluorescence in situ hybridization (FISH) procedure was performed using the Ribo™ Fluorescent in Situ Hybridization Kit (RiboBio). In brief, primary CMs or tissues were incubated with 4% paraformaldehyde for 10 min and washed with PBS (Servicebio), followed by 0.1% Triton X-100 treatment for 10 min.
After incubation with the post-hybridization solution in a humid chamber at 37°C for 30 min, the probe targeting circIGF1R was added to cells at 37°C overnight. Next, hybridization solution I, II, and III was used to wash cells at 42°C, respectively. Cells or tissues were then counterstained with DPAI (Sigma-Aldrich).
Bioinformatics analysis
To predict the targeted miRNA that can interact with circIGF1R, multiple bioinformatic programs including starBase (
The potential target genes of mmu-miR-362-5p were predicted by starBase (
Statistical analysis
The GraphPad Prism 8.4.3 software was used to calculate the statistics. Data are presented as the mean ± the standard error of the mean. All samples were performed at least in triplicate. Comparisons between the two groups were analyzed by unpaired Student's t-test. Among three or more groups, statistical analysis was performed using a one-way analysis of variance followed by Dunnett's multiple-comparison tests. A p < 0.05 was considered statistically significant.
Result
Expression profile of circIGF1R during heart growth and regeneration
The circIGF1R derives from the second exon of IGF1R. The expression of circIGF1R was detected in a range of tissues, indicating that circIGF1R may have conservative functions in multiple organs (Fig. 1A). Initially, we designed divergent primers to detect the circIGF1R in the cDNA but not in genomic DNA from cardiomyocyte line (HL-1) and primary CMs. The sequencing of PCR products identified a head-to-tail loop sequence of the second exon of the IGF1R (Fig. 1B), which was consistent with the circular RNA mmu_circ_0001582 from the circBase. Next, we designed convergent primers to validate the stability between linear transcript and circular transcript. RNase-R resistance assays showed that the circIGF1R was resistant to RNase-R, whereas the linear transcript was completely degraded (Fig. 1C).

circIGF1R is up-regulated after heart apical resection.
After the verification of circIGF1R, we focused on the expression patterns of the circIGF1R during heart growth and regeneration. Interestingly, qPCR showed that expression of the circIGF1R was significantly decreased during heart development (Fig. 1D). CircIGF1R expression was considerably down-regulated in both adult hearts and CMs compared with neonatal mice at p1 (Fig. 1D, E). In addition, circIGF1R expression was decreased in adult CFs. (Fig. 1F).
Further, we detected high circIGF1R expression in injured neonatal hearts at 0.5 dpr and 1 dpr compared with sham groups (Fig. 1H, I). Taken together, the data showed that circIGF1R may play a principal role during heart repair. Moreover, we designed a specific probe that targeted the head-to-tail junction position of circIGF1R to detect the subcellular localization of circIGF1R, and the RNA fluorescence in situ hybridization (RNA-FISH) of the circIGF1R showed that the circIGF1R was mainly located in the cytoplasm of primary CMs (Fig. 1J).
Expression profile of circIGF1R in injured neonatal heart
Given that circIGF1R levels were increased in the injured neonatal heart, the specific probe of circIGF1R was applied in the injured neonatal heart. Interestingly, more circIGF1R expression was observed in the injured area of the injured neonatal heart at 1 dpr, as indicated by fluorescence intensity of the injured area to no-injured area ratio, compared with sham groups (Fig. 2A, B). Combined qPCR and IF results may implicate circIGF1R as a novel regulator in heart regeneration.
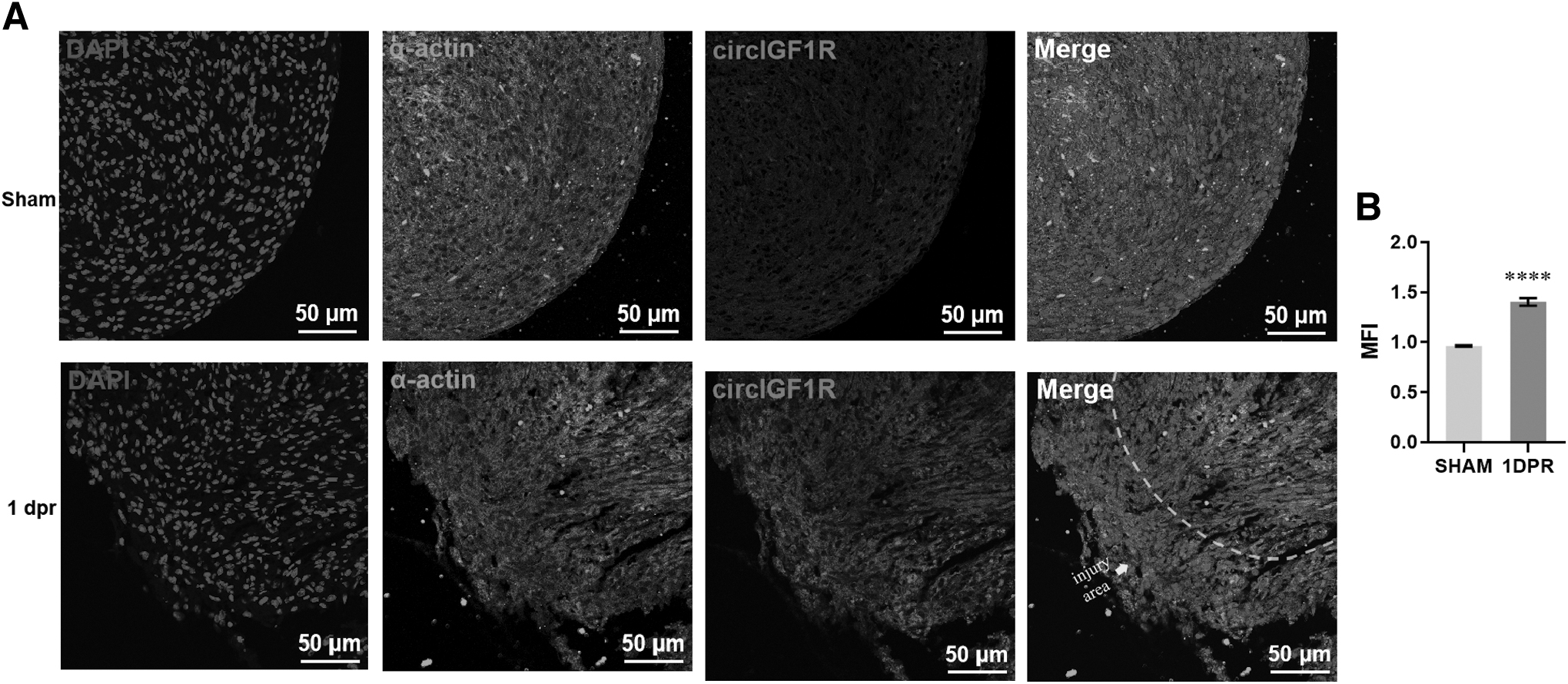
Expression profile of circIGF1R in injured neonatal heart.
Loss of circIGF1R inhibits the proliferation of the neonatal CMs
According to the expression profiles of circIGF1R during heart growth and regeneration, we speculated that there was a positive correlation between circIGF1R and heart regeneration. To test our speculation, we designed a specific siRNA (small interfering RNAs), si-circIGF1R, to specifically knock down circIGF1R instead of the linear transcript in HL-1 and primary CMs (Fig. 3A, B).

Loss of circIGF1R inhibits the proliferation of neonatal cardiomyocytes.
The qPCR result showed that the expression of circIGF1R was pronouncedly decreased in HL-1 after treatment with siRNA, but the expression of IGF1R mRNA was not changed (Fig. 3C, D). In agreement with the HL-1 cell line, specific knockdown of circIGF1R rather than IGF1R mRNA was detected in the primary CMs transfected si-circIGF1R (Fig. 3E, F). Subsequently, we investigated the effect of circIGF1R knockdown on the proliferation of primary CMs using immunofluorescent staining of proliferating markers.
Our data showed that circIGF1R knockdown significantly decreased the number of Ki67+ α-Actin+ CMs compared with the control group in vitro (Fig. 3G, H). In concert with the result, loss of circIGF1R could also inhibit the incorporation of EdU+ α-Actin+ CMs (Fig. 3I, J). To prove complete cardiomyocyte proliferation, late cell cycle marker phospho-Histone H3 (pH3) was further used in this study. Our data showed that the percentage of pH3+ α-Actin+ cells was significantly decreased by si-circIGF1R treatment (Fig. 3K, L), indicating that circIGF1R knockdown suppresses the complete proliferation of CMs in vitro. In conclusion, our results convincingly demonstrated that circIGF1R plays an important role in the proliferation of primary neonatal CMs, which implies the potential importance of circIGF1R for heart regeneration in neonatal mice.
miR-362-5p was downstream of circIGF1R
Since most of the circRNAs execute their biological function as miRNA sponges and circIGF1R mainly locates in the cytoplasm, we wondered whether circIGF1R could promote heart regeneration in such a mechanism. To determine the potential miRNA target of circIGF1R, miRanda, starBase, and RNA hybrid databases were used to preliminarily predict the circIGF1R -targeting miRNAs. The overlapping result revealed that three potential targeting miRNAs, including mmu-miR-500-5p, mmu-miR-362-5p, and mmu-miR-670-3p, were identified (Fig. 4A).

miR-362-5p was downstream of circIGF1R.
Subsequently, we respectively inserted the full-length of circIGF1R and Mut-circIGF1R in the dual-luciferase miRNA target expression vector pmirGLO for dual-luciferase assays (Fig. 4B). The dual-luciferase assays revealed that three potential targeting miRNAs decreased the luciferase activity of the report gene constructed with wild-type circIGF1R (WT reporter) compared with mimic-NC. Among these three potential targets, only miR-362-5p significantly decreased the luciferase activity of the report gene constructed with WT reporter compared with mutant circIGF1R (Mut reporter) (Fig. 4C).
These data indicate that only miR-362-5p out of three potential targets might be the one that can directly bind to circIGF1R. As expected, the expression of miR-362-5p was significantly increased by the knockdown of circIGF1R in primary CMs (Fig. 4D). Next, we use miRNA mimics and inhibitor (mimic-miR-362-5p and si-miR-362-5p) to overexpress and knock down miR-362-5p for further study, verifying in CMs (Fig. 4E, F). Meanwhile, after si-miR-362-5p was treated in cells, expression of miR-362-5p can be rescued by si-miR-362-5p in the circIGF1R-knockdown cells (Fig. 4E).
These data uncovered that miR-362-5p was the direct target of circIGF1R. Subsequently, we performed the expression patterns of miR-362-5p during heart development and regeneration. The qPCR result indicated that miR-362-5p was significantly up-regulated in adult CMs compared with neonatal CMs (Fig. 4H). On the contrary, the expression of miR-362-5p was dramatically down-regulated in the injured neonatal heart at 0.5 dpr and 1 dpr compared with sham groups (Fig. 4I, J). These data revealed that miR-362-5p could negatively control CMs proliferation, whose expression patterns were opposite to the expression profile of circIGF1R during heart growth and regeneration.
miR-362-5p silencing can rescue the proliferation inhibition of CMs induced by circIGF1R knockdown
Because miR-362-5p was up-regulated in adult CMs and down-regulated in the injured neonatal heart, we subsequently explored the effect of miR-362-5p on CMs proliferation. Then, the percentages of Ki67+ α-Actin+ CMs decreased after transfected with mimic-362-5p compared with controls (Fig. 5A, B). Consistent with Ki67 staining, EdU staining also demonstrated that the incorporation of EdU+ α-Actin+ CMs was significantly reduced on miR-362-5p overexpression (Fig. 5C, D).

miR-362-5p silencing can rescue the inhibition of proliferation inhibition of CMs induced by circIGF1R knockdown.
In addition, pH3 staining further demonstrated that miR-362-5p overexpression could significantly inhibit CMs proliferation as demonstrated by the decreased percentage of pH3+ α-Actin+ cells (Fig. 5E, F). We then investigated whether si-miR-362-5p could rescue the inhibition of proliferation following circIGF1R knockdown. The results showed that si-miR-362-5p treatment increased the proportion of EdU-, Ki67-, and pH3-positive CMs and reversed the suppressed proliferation phenotype observed in circIGF1R-knockdown cells (Fig. 5G–L). In conclusion, these results elucidated that circIGF1R acted as an miR-362-5p sponge to regulate CMs proliferation.
Phf3 is a target gene of miR-362-5p and circIGF1R
The function of miRNA was the formation of an RNA-induced silencing complex by specifically binding the 3′-UTRs of mRNA to inhibit gene expression. To identify the potential targets of miR-362-5p, three software including Diana-microt, tarbase, and starBase were used in this study. Overlapping results revealed that Phf3 is the only one identified from each software (Fig. 6A). Previous studies have reported that miR-362-5p can greatly suppress the expression of several target genes, including CYLD, QK, GAS7, Sema3a, and Gadd45a, in various cell types.
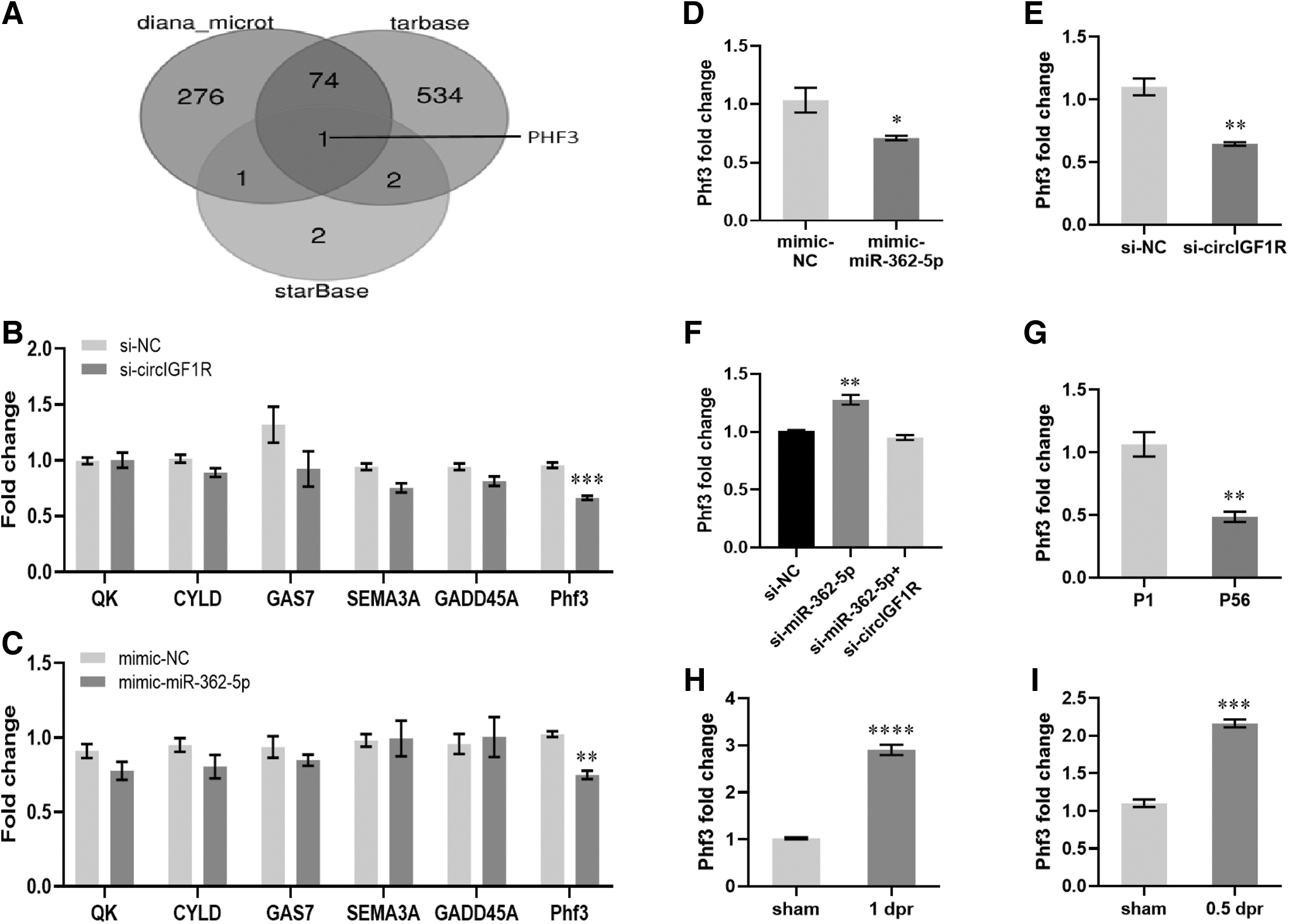
Phf3 is a target gene of miR-362-5p and circIGF1R.
Therefore, these six candidates were further examined in this study. Among these six candidates, only PHD-finger protein 3 (Phf3) was down-regulated by the overexpression of miR-362-5p and by the knockdown of circIGF1R in HL-1, respectively (Fig. 6B, C). Moreover, remarkably decreased expression of Phf3 was also detected in the primary CMs transfected with mimic-362-5p or si-circIGF1R compared with controls (Fig. 6D, E), and si-miR-362-5p treatment rescued the expression of Phf3 in circIGF1R-knockdown cells (Fig. 6F).
Subsequently, we explored the Phf3 expression patterns in the heart and found that the expression of Phf3 was decreased in adult CMs compared with neonatal CMs (Fig. 6G). Consistently, increasing Phf3 expression was detected in neonatal hearts at 0.5 dpr and 1 dpr, respectively (Fig. 6H, I). In summary, our data showed that the expression pattern of Phf3 in the heart was positive, correlated with circIGF1R and that down-regulation of circIGF1R suppressed the proliferation of CMs via miR-362-5p/Phf3 axis.
Knockdown of Phf3 repressed CMs proliferation
Because Phf3 was the common downstream of miR-362-5p and circIGF1R and the expression patterns of Phf3 were consistent with circIGF1R in CMs and injured hearts, we investigated the Phf3 effect on CMs proliferation. SiRNA of Phf3 (si-Phf3)-induced knockdown of Phf3 was verified in primary CMs (Fig. 7A). Subsequently, the IF of Ki67 demonstrated that decreasing CMs was detected after transfected with si-Phf3 compared with controls (Fig. 7B, C). In accordance with Ki67 staining, the staining of EdU also manifested that the incorporation of EdU+ α-Actin+ CMs was significantly reduced on the knockdown of Phf3 (Fig. 7D, E).

Knockdown of Phf3 repressed CMs proliferation.
Moreover, pH3 staining further confirmed that loss of Phf3 could significantly repress CMs proliferation, demonstrated by the decreased percentage of pH3+ α-Actin+ cells (Fig. 7F, G). In closing, our results elucidated that circIGF1R acted as an miR-362-5p sponge to regulate Phf3 expression; as a result, it regulated the proliferation of CMs.
Discussion
Discovering the underlying mechanisms that are involved in cardiac regeneration is helpful to resolve the heart damage caused by myocardial ischemia/reperfusion injury in humans. This is because the ability of regeneration in neonatal mouse hearts occurs within 7 days after birth (Hsieh et al., 2007; Porrello et al., 2011), which means their transcript has greatly different from the adult heart (Werfel et al., 2016). In this study, we focused on a novel circular RNA circIGF1R. Our data revealed that circIGF1R contributes significantly to neonatal CM proliferation, at least in part by functioning as an miRNA sponge to interact with miR-362-5p and then affect Phf3 expression.
The circIGF1R derives from the second exon of IGF1R (insulin-like growth factor I receptor). Although the IGF1R produces a variety of circRNAs, this particular circRNA (mmu circ 0001582 [ID from circBase]) is better investigated and serves a greater function than others. In the current research, we showed that circIGF1R rises within a day of cardiac apex excision and is highly expressed in newborn hearts (Fig. 1). Importantly, knockdown of circIGF1R significantly inhibits the proliferation of primary CMs isolated from the neonatal mouse heart (Fig. 2).
These data strongly indicate that circIGF1R might be involved in CM proliferation. Before our research, some articles also investigated circIGF1R, but the circIGF1R was found in neuronal, non-small cell lung cancer or hepatocellular carcinoma (HCC) (Fu et al., 2019; Shao et al., 2021; Xu et al., 2020). For example, circIGF1R was found to promote the polarization of astrocytes to phenotype A1 by inhibiting autophagy (Shao et al., 2021), implicating that circIGF1R had a function in the suppression of autophagy in neuronal CMs.
And the other article showed that circIGF1R had pro-proliferative and anti-apoptotic effects in HCC (Fu et al., 2019). This research hints that circIGF1R could be involved in apoptosis or autophagy in CMs. In addition, the RNA-sequencing in mouse and human cardiac hypertrophy samples showed that circIGF1R was increased in cardiac hypertrophy compared with sham or normal groups (Tan, 2017; Werfel et al., 2016). All data indicate that circIGF1R participates in the important physiological functions in CMs and the heart.
The research on miRNAs in cardiovascular disease has appeared in lots of articles (Bonauer et al., 2009; Care et al., 2007; Eulalio et al., 2012; van Rooij et al., 2007). In this study, we revealed that miR-362-5p had an important role in CMs. Increasing miR-362-5p expression in adult CMs was detected. On the contrary, the expression of miR-362-5p was decreased in the injured neonatal heart. The functional experiments showed that overexpression of miR-362-5p reduced the proliferation of CMs.
These results uncovered that miR-362-5p played a negative role in CMs. Moreover, the non-coding RNA-sequencing indicated that miR-362-5p was down-regulated in mice hearts 21 days post-myocardial infarction comparing the sham control, which confirmed that miR-362-5p was a negative gene in heart regeneration (Tho et al., 2016). In another report, miR-362-5p was found in neuroblastoma. Increasing miR-362-5p inhibited neuroblastoma progression by targeting NOL4L (Chen et al., 2023) and phosphatidylinositol 3-kinase-C2β (PI3K-C2β) (Wu et al., 2015).
Previous studies have shown that NOL4L and PI3K-C2β are the target genes of miR-362-5p in neuroblastoma (Chen et al., 2023; Wu et al., 2015). In this study, we demonstrated that Phf3 was downstream of miR-362-5p. Phf3 was first reported in research about glioblastoma, and the Phf3 expression was found to be significantly down-regulated in glioblastoma (Fischer et al., 2001). Recently, Phf3 was reported to participate in regulating neuronal gene expression (Appel et al., 2021).
Phf3 was required for proper neuronal differentiation by participating in mRNA stability (Appel et al., 2021). Further, Phf3 interacts with RNA polymerase II via the SPOC domain to prevent the precocious expression of a subset of neuronal genes (Appel et al., 2021). The RNA-seq showed that loss of Phf3 in HEK293T cells led to increased Pol II stalling and reduced elongation rates, causing about 600 genes to be strongly derepressed (>2-fold increase in mature transcript levels) (Appel et al., 2021). In addition, most of the up-regulated transcripts are involved in the regulation of cell proliferation and regulation of cell differentiation, indicating that Phf3 may have stupendous strength in cell proliferation and regulation in CMs.
In conclusion, our results indicate that circIGF1R has the capacity to regulate cardiomyocyte proliferation through the miR-362-5p/Phf3 axis, which provided new insight into heart regeneration. Notably, CM labeling was generally performed by whole-cell staining of CMs in the present study. The strict quantification method had been applied in minimizing the potential interference of proliferating no-CMs. However, most rigorous methods, such as cell fate tracing technology and nuclear staining of CMs, are eager to be used to support our data in the future.
Footnotes
Authors' Contributions
Study design and concept by X.-F.Q. and D.-Q.C. Experimental design and undertaking by J.-H.Z. and X.-F.Q. Cell culture by J.-H.Z., H.-J.L., W.-L.Z., and H.-Y.W.. Additional samples were supplied by C.-Q.L., H.-Y.W., and M.-H.T. Heart apical resection operation was implemented by W.-Y.C. Data analysis by Jun-Hui Zeng, Xu-Feng Qi and Kun Liu. Manuscript written and edited by J.-H.Z., X.-F.Q., and K.L.
Data Availability Statement
The data that support the findings of this study are available from the corresponding authors on reasonable request.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the National Natural Science Foundation of China (Grant Nos. 82070257, 81770240, 81570222), GuangDong Basic and Applied Basic Research Foundation (Grant No. 2023A1515012147), and the Research Grant of Key Laboratory of Regenerative Medicine, Ministry of Education, Jinan University (Grant Nos. ZSYX-M-2019-00009, ZSYXM202004, and ZSYXM202104), China.
Supplementary Material
Supplementary Tables
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.