
Abstract
Thrombosis is a common problem with potentially severe consequences. Endothelial progenitor cells (EPCs) show great potential as a thrombosis therapy due to their angiogenesis-promoting, thrombus-relieving, and anticoagulant functions. However, cell therapies present more clinical challenges than small molecule solutions. MicroRNAs (miRNAs) are small noncoding single-stranded RNAs with wide-ranging regulatory activities. miRNA-126 is highly enriched in EPCs and endothelial cells. Although increasing research showed that mircoRNA-126 (miR-126) can regulate EPC functions through various pathways and cytokines, summaries of these interactions are rare. Therefore, this brief review of recent findings on the relationship between miRNA-126 and EPC function will attempt to clarify the role of miR-126 in thrombosis through regulation of EPCs, with the goal of exploring alternative therapies for thrombotic diseases.
Introduction
Thrombosis is a common disorder that can occur independently or concurrently with other diseases. Deep vein thrombosis (DVT), a frequent and serious complication of surgery, can result in pulmonary hypertension and pulmonary embolism. Thrombosis is of particular concern for diabetic patients since most of them are already in a prethrombotic state (Tangvarasittichai, 2015), which increases their risk for atherosclerotic thrombotic events by two- to fourfolds compared with nondiabetic patients.
Approximately 80% of deaths in patients with type 2 diabetes are due to thrombotic events (Gerber and Rutter, 2017). Similarly, cerebral infarction caused by cerebral thrombosis is a serious threat to human health and life, with high morbidity and mortality.
Endothelial cells are essential components of blood vessels and are derived from differentiation of endothelial progenitor cells (EPCs), a type of stem cell first identified in the bone marrow in 1997. As EPCs are increasingly studied, they are gradually being distinguished into two different subtypes, namely, myeloid angiogenic cells (MACs) and endothelial colony forming cells (ECFCs), which are also referred to as early EPCs and late EPCs (Medina et al., 2017, Medina et al., 2011; Prasain et al., 2015; Yoon et al., 2005).
A growing body of evidence highlights critical roles for EPCs in angiogenesis, and endothelial dysfunction and repair. EPCs may provide alternative means of treating cardiovascular diseases because of their proangiogenic and protective cytokine release potential (Huang and Huang, 2022; Zeng et al., 2021).
MicroRNAs (miRNAs) are small noncoding RNAs that regulate biological processes in animals and plants at the epigenetic level. These complex miRNA networks are involved in health (regulating cellular function, differentiation, and development [Plotkin and Wallace, 2021; Sengupta et al., 2021]) and disease (including cancer [Sempere et al., 2021], diabetes [Wonnacott et al., 2022], and cardiovascular disease [Song et al., 2020b]). In addition, they can also play a diagnostic role in diseases as diagnostic markers (Blanco-Domínguez et al., 2021).
Among them, mircoRNA-126 (miR-126) is highly enriched in EPCs and endothelial cells (Chistiakov, et al., 2016), and can affect EPCs through various mechanisms. This article briefly reviews the process of thrombosis and the role of EPCs in thrombosis and elucidates the mechanism by which miR-126 regulates EPC function. In addition, we also discuss alternative strategies for the treatment and prevention of thrombosis.
Thrombosis
Thrombus can be divided into arterial thrombus and venous thrombus depending on the site of origin. Arterial thrombus is a platelet-rich thrombus formed on the basis of atherosclerotic plaque rupture and high shear flow. When endothelial cells are injured, vascular barrier function is impaired and endothelial integrity and permeability are altered (Gimbrone and García-Cardeña, 2016). At the same time, nitric oxide and prostacyclin decrease and thromboxane and vascular hemophilia factor increase to mediate platelet aggregation and adhesion to promote thrombosis (Kaur et al., 2018).
Unlike arterial thrombosis, venous thrombosis occurs under low shear flow and mostly around intact endothelial wall. Venous thrombi are rich in fibrin and encase a large number of red blood cells in addition to activated platelets. Inflammation, hypoxia due to low blood flow, and other states allow endothelial cell activation. Activation of endothelial cells increases the levels of vascular hemophilia factor, P-selectin, intercellular adhesion factor 1, and vascular adhesion molecule 1, which in turn cause platelet and leukocyte adhesion (Dhami et al., 2022; Horváth et al., 2020; Li et al., 2021 ; Preston et al., 2019).
Attached leukocytes become activated and initiate expression of TF that, in turn, activates the coagulation cascade (Von Brühl et al., 2012), leading to the activation of thrombin and the conversion of fibrinogen to fibrin. These processes combine to promote venous thrombosis.
EPCs for Thrombosis Resolution
In mice models of arterial thrombosis, MACs and ECFCs transplants can accelerate thrombosis resolution (Carneiro et al., 2019). This may be related to PAR-1 signaling in these cells, their ability to promote angiogenesis, and their anticoagulant effects (Smadja et al., 2008, Smadja et al., 2006). The anticoagulatory effects of MACs may be related to inhibition of platelet aggregation through P-selectin and contribute to a reduction in thrombus formation (Abou-Saleh et al., 2015). Furthermore, ECFCs can improve the microenvironment by regulating the changes of VEGF, monocyte chemoattractant protein (MCP)-1, and ANG-1 in the vascular lumen and accelerate the resolution of thrombosis (LI et al., 2007; Meng et al., 2010).
In recent years, more advanced research to improve thrombosis treatment with EPCs include engineering MACs to overexpress platelet-derived growth factor-D (Zhang and Luo et al., 2021), and using MACs and ECFCs-derived exosomes. The MACs-derived exosomes can reduce the length and weight of venous thrombosis in DVT mice by inhibiting TXNIP, suppress apoptosis and inflammatory responses, decrease fibrinogen and thrombin–antithrombin complex levels, and increase fibrinolytic enzyme (Feng et al., 2022). In addition, ECFCs-derived exosomes can suppress iron-induced endothelial cell death by inhibiting SP1 and activating AMPK pathways (Xia et al., 2022).
miR-126 Regulates the Function of EPCs
miRNAs are involved in a variety of cellular functions, including angiogenesis, and miR-126, in particular, has been shown to enhance this aspect (Urbich et al., 2008). Some effects of miR-126 are mediated by the PI3K/Protein Kinase B (Akt) pathway. An enzymatic subunit of PI3K, PIK3R2, can be inhibited by miR-126, leading to reduced apoptosis, increased proliferation and angiogenesis, and improved cerebral collateral circulation (Hao et al., 2021; Song et al., 2020a; Tang et al., 2021; Zhang et al., 2021).
Similarly, miR-126 inhibition of PIK3R2 improves the function of EPCs under hypoxic conditions (Zhang and Xu et al., 2022), and promotes ECFCs angiogenesis and accelerates thrombolysis in a rat model of DVT (Meng et al., 2015). In addition, miR-126 can promote MACs angiogenesis, proliferation, and migration through sprouty-related EVH1 domain containing 1 (Spred-1) targeting of rat sarcoma (Ras)/extracellular regulated protein kinases (ERK) pathways (Meng et al., 2012; Sabzevari et al., 2020).
In addition to promoting angiogenesis, miR-126 overexpression can increase the homing of ECFCs to arterial thrombi by promoting C X C chemokine receptor 4 (CXCR4) expression and pathways involving ERK/VEGF and Akt/endothelial nitric oxide synthase, thereby protecting against vascular injury (Pei et al., 2020; Wang et al., 2018). Hyperglycemia and advanced glycation end products (AGEs) can lead to dysfunction of MACs by increasing inflammation, inhibiting the PI3K/Akt pathway and Ras/ERK pathways, and reducing miR-126 expression. Restoring miR-126 expression may partially protect EPCs in this condition (Li et al., 2016; Meng et al., 2012).
Similarly, miR-126 can also improve MACs function under oxidative stress by regulating the PI3K/AKT signaling pathway (Wu et al., 2021). Besides targeting PIK3R2 and Spred-1, miR-126 has been shown to regulate ERK1/2 (Wu et al., 2021), Notch (Kong et al., 2020), and the stromal cell-derived factor 1 (SDF-1)/CXCR7 signaling pathway (Li et al., 2021 ; Shan and Ma, 2018) to promote MACs or ECFCs proliferation and migration, and enhance vascular repair and regeneration. These studies suggest that miR-126 regulates not only EPCs by numerous mechanisms but also across a wide range of activities, including proliferation, migration, homing, and angiogenesis (Fig. 1).
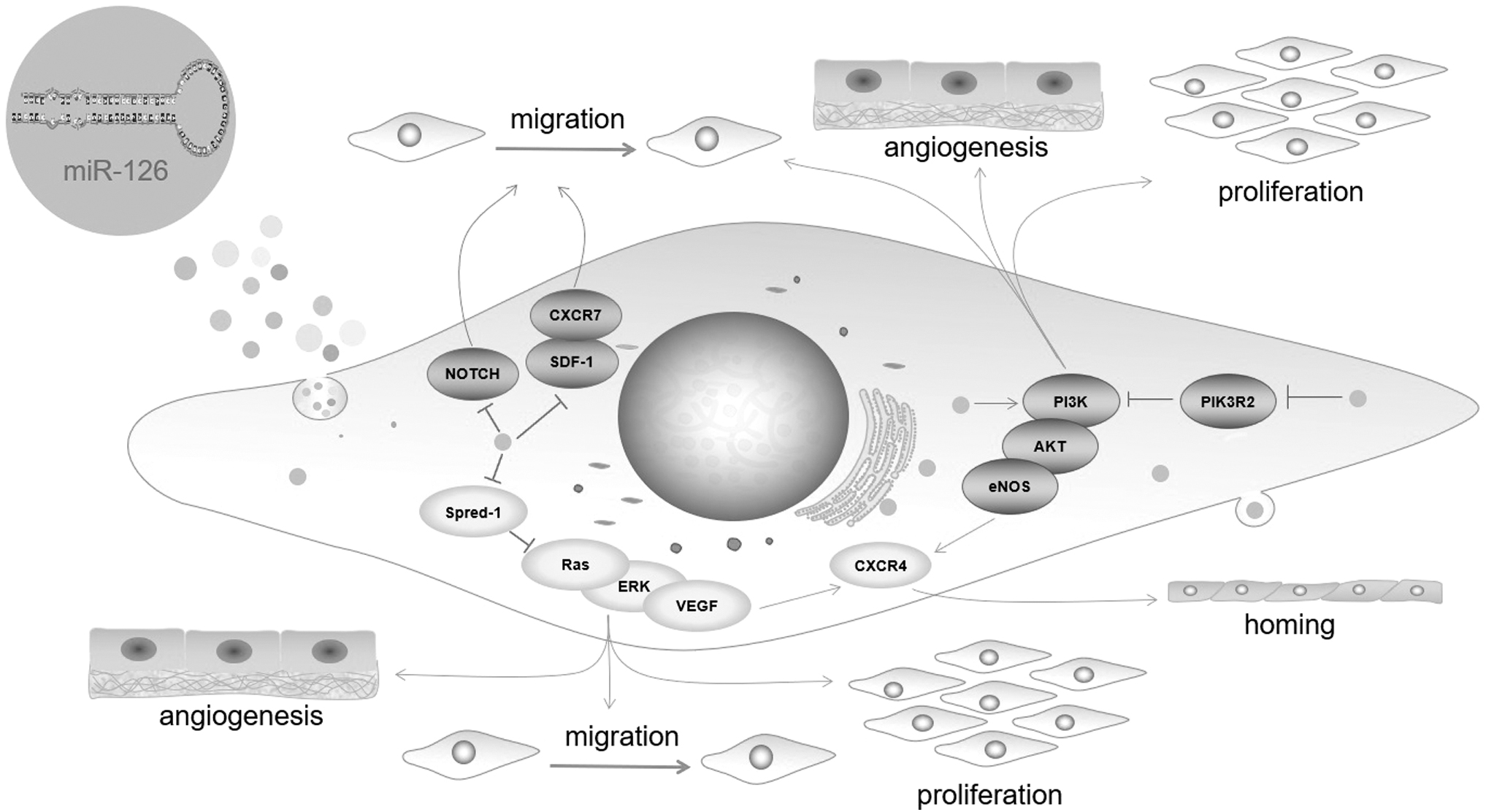
miRNA-126 regulates EPC-related signaling pathways. MicoRNA-126 regulates EPC proliferation, migration, homing, and angiogenesis by targeting PIK3R2 and Spred-1. The SDF-1/CXCR7 and Notch channels regulate EPC migration. EPC, endothelial progenitor cell; miRNA, microRNA; PIK3R2, phosphoinositide-3-kinase regulatory subunit 2; SDF-1, stromal cell-derived factor 1; Spred-1, sprouty-related EVH1 domain containing 1.
miR-126 can also affect EPC functions by regulating cytokine expression. In vitro overexpression of miR-126-3p in EPCs upregulates numerous cytokines including angiopoietin, granulocyte colony-stimulating factor (G-CSF), hepatocyte growth factor (HGF), insulin growth factor-1 (IGF-1), interleukin (IL)-3 and -10, matrix metalloproteinase inhibitor-1 (TIMP-1) and -2, and VEGF-A, while reducing others such as IL-8, MCP-1 and -2, macrophage inflammatory protein-1β (MIP-1β), and tumor necrosis factor (TNF)-α and -β (Li et al., 2018).
Among them, miR-126 can target PIK3R2 and Spred-1 to regulate VEGF, and can also target PIK3R2 to regulate angiopoietin (Hao et al., 2021; Sessa et al., 2012). In addition, miR-126 can target sphingosine-1-phosphate receptor 2 (SIPR2) to regulate TNF-α, IL-10, and glycogen synthase kinase-3β (GSK-3β) to regulate TNF-α, IL-8 (Fan et al., 2020; Guo et al., 2022). These cytokines regulate many aspects of EPC function, from survival to migration, and angiogenesis.
Cytokines that are upregulated by miR-126 generally promote ECFCs or MACs survival, proliferation, and differentiation, and include angiopoietin (Sudchada et al., 2011), HGF (Zhang et al., 2022), IGF-1 (Chen et al., 2021), IL-10 (Short et al., 2022), and VEGF (Yang et al., 2022). Migration is essential for tissues or organs for growth and repair. Cytokines that promote ECFCs or MACs migration include G-CSF (Perdomo et al., 2022), HGF (Zhang et al., 2022), IL-10 (Short et al., 2022), and VEGF (Yang et al., 2022). Angiogenesis is an important aspect of thrombus resolution. Cytokines that promote the angiogenic functions of ECFCs or MACs include IGF-1 (Chen et al., 2021), IL-3 (Moldenhauer et al., 2015), and IL-10 (Short et al., 2022).
Among these cytokines, angiopoietin, HGF, G-CSF, and IL-3 promote the function of ECFCs, whereas VEGF promotes the function of MACs. Conversely, cytokines that are downregulated by miR-126 tend to inhibit functions of ECFCs and MACs. Proinflammatory cytokines such as IL-8 (Ha et al., 2021) and MCP-1 (Shoeibi, 2021) may have adverse effects on ECFCs, and MIP-1β (Zhao and He, 2021) may have adverse effects on MACs, and they may impair the proliferation and angiogenesis of both types of cells. Tumor necrosis factor, in particular TNF-α, has been shown to impair proliferation, migration, adhesion, and angiogenesis of ECFCs (Zhang et al., 2020).
Less is known about the effect of TNF-β, but based on the structural and functional similarities between these two proteins (Li et al., 2021 ), they are likely to have similar effects on ECFCs. However, the functions regulated by miR-126 in ECFCs are likely to be much more complex. For example, miR-126 overexpression increases antivascular factors such TIMP-1 and -2, which correlate negatively with activities of ECFCs (Ray et al., 2020), whereas proangiogenic factors, such as MCP-2 (Litwin et al., 2015), are reduced by miR-126 overexpression. Despite the seemingly contradictory evidence, these studies, nevertheless, suggest that miR-126 can regulate EPC function by altering cytokine expression (Fig. 2).
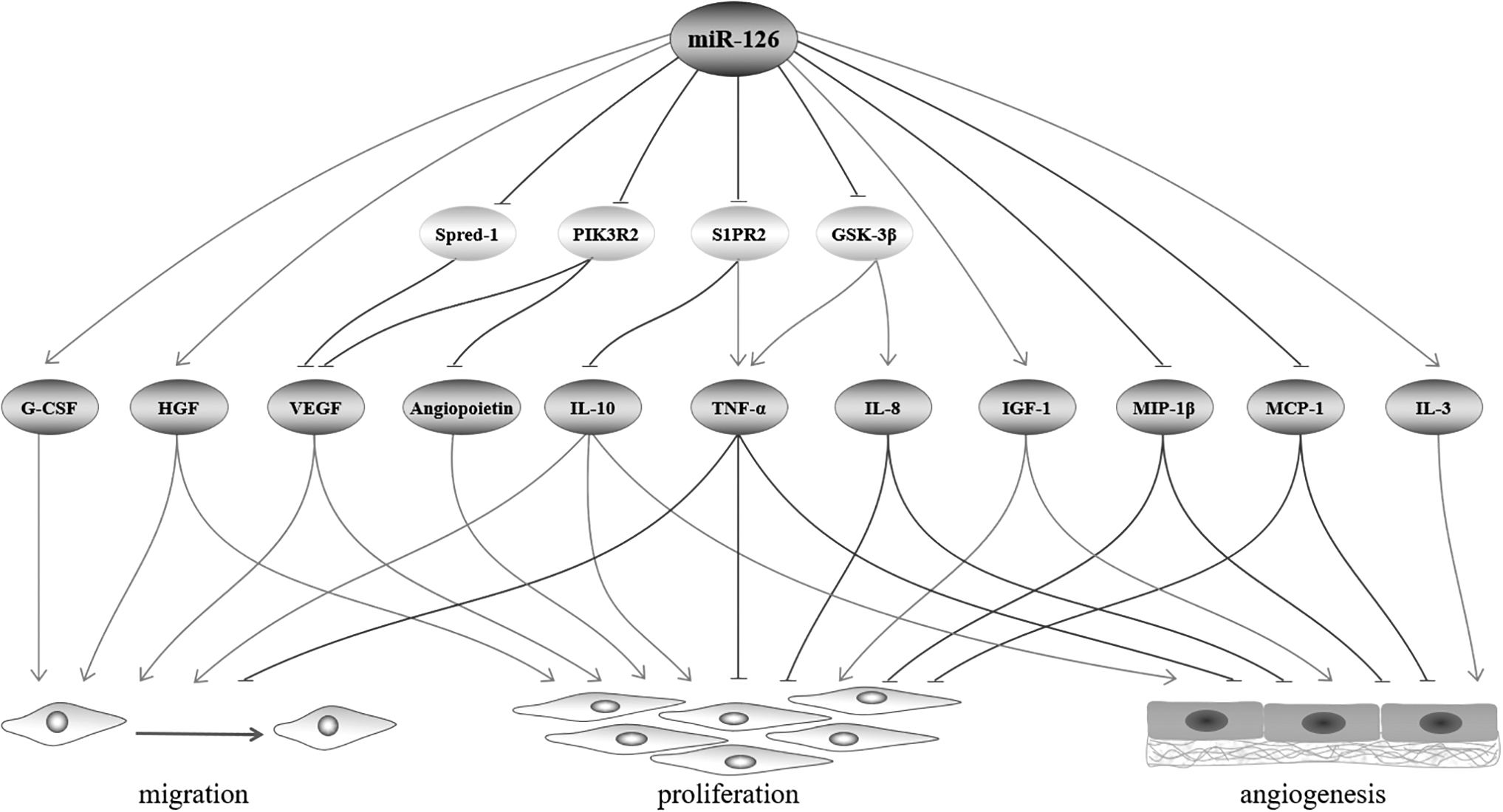
miRNA-126 affects EPC function by regulating cytokine expression. Including upregulation of G-CSF, HGF, VEGF, IL-10, angiopoietin, IGF-1, and IL-3 and downregulation of IL-8, MCP-1, MIP-1β, and TNF-α. G-CSF, granulocyte colony-stimulating factor; HGF, hepatocyte growth factor; IGF-1, insulin growth factor-1; IL, interleukin; MCP-1, monocyte chemoattractant protein-1; MIP-1β, macrophage inflammatory protein-1β; TNF-α, tumor necrosis factor-alpha.
Conclusions
In this brief review, we first describe the process of arterial and venous thrombosis and second highlighted the importance of EPCs in thrombosis. The antithrombotic effects of EPCs stem from their ability to repair endothelial cells, promote angiogenesis, inhibit platelet aggregation, and improve the vascular microenvironment. In addition, the EPC-derived exosomes can help to relieve blood clots. We also provided an overview of the direct and indirection ways in which miR-126 can regulate EPC functions.
Direct signaling from miR-126 can regulate EPC proliferation and homing, migration, and angiogenesis. These EPC activities are also indirectly regulated by the cytokines produced upon miR-126 signaling. Both pro- and antiangiogenic, as well as pro- and anti-inflammatory cytokines contribute to the fine tuning of EPC functions. In summary, we found that the regulation of EPCs by miR-126 is wide ranging, and largely positive toward thrombus resolution (Table 1).
Function and Target of MicroRNA-126
GSK-3β, glycogen synthase kinase-3β; PIK3R2, phosphoinositide-3-kinase regulatory subunit 2; SDF-1, stromal cell-derived factor 1; SIPR2, sphingosine-1-phosphate receptor 2; Spred-1, sprouty-related EVH1 domain containing 1.
Perspectives
EPCs are important players for the mitigation of thrombosis. Increasing research on miRNAs is providing more insights into the interactions between these small molecules and EPC function. In this study, we found abundant evidence that miR-126 can regulate the proliferation, mobilization, migration, and recruitment of EPCs, as well as activate their proangiogenic functions to drive thrombotic regression. Based on the mentioned evidence, it is reasonable to speculate that miR-126 could mitigate thrombosis by enhancing the activities of EPCs.
Therefore, novel therapies for thrombosis may be derived from miR-126 or the molecules that promote miR-126 expression. However, the mechanism of action and effect of miR-126 on different subtypes of EPC differ and need to be further differentiated. In addition, most of the existing studies were conducted in animals and clinical evidence is required to support the feasibility of targeting miR-126 for thrombosis management.
Footnotes
Authors' Contributions
Q.X., D.W., Y.S., J.H., and X.Q.H. contributed to the conception and design of the study. Q.X. and D.W. searched the relevant literature. Q.X. wrote the article. X.Q.H. provided advice and was responsible for revising the article. All authors have read and approved the final version of the article.
Availability of Data and Materials
Data sharing is not applicable to this article, as no data sets were generated or analyzed during this study.
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the National Natural Science Foundation of China (Grant No. 81273568).