
Abstract
Constitutive photomorphogenic 1 (COP1), is an E3 ubiquitin ligase that plays a role in the regulation of various cellular processes including cell growth, differentiation, and survival in mammals. In certain conditions such as overexpression or loss of function, COP1 acts either as an oncogenic protein or as a tumor suppressor by targeting specific proteins for ubiquitination-mediated degradation. However, the precise role of COP1 has not been well studied in primary articular chondrocytes. In this study, we investigated the role of COP1 in chondrocyte differentiation. Western blotting and reverse transcription–polymerase chain reaction analysis demonstrated that COP1 overexpression reduced type II collagen expression, promoted cyclooxygenase 2 (COX-2) expression, and reduced sulfated proteoglycan synthesis, as detected by Alcian blue staining. Upon siRNA treatment, revived type II collagen, sulfated proteoglycan production, and decreased COX-2 expression. Phosphorylation of p38 kinase and ERK-1/-2 signaling pathways was regulated by COP1 upon cDNA and siRNA transfection in chondrocytes. The inhibition of the p38 kinase and ERK-1/-2 signaling pathways with SB203580 and PD98059 ameliorated the expression of type II collagen and COX-2 in transfected chondrocytes, thus suggesting that COP1 regulates differentiation and inflammation in rabbit articular chondrocytes via the p38 kinase and ERK-1/-2 signaling pathway.
Introduction
Articular cartilage in the joints of knees and elbows, which comprises soft tissue, is prominent throughout the mammalian body and cushions the bones and facilitates movement (Shi et al., 2019; Sutherland et al., 2015). Unlike other tissues, cartilage does not have a direct blood supply; therefore, growth and self recovery are limited. Cartilage cells, which account for ∼2% of the joint cartilage volume in humans and developed from mesenchymal stem cells (Sophia Fox et al., 2009), produce chondrocyte-specific extracellular matrix (ECM) to maintain matrix integrity (Carballo et al., 2017). Chondrocytes are cells that are responsible for maintaining the ECM and producing the cartilage matrix in cartilage tissue. They are surrounded by collagenous fibers and release substances that make cartilage strong yet flexible. Chondrocytes are found within intervertebral discs and in any form of articular cartilage. Chondrocytes play a crucial role in cartilage formation and maintenance. ECM of chondrocytes is composed of a complex network of macromolecules including proteoglycans, glycosaminoglycans (GAGs), and collagens. These macromolecules provide the structural support and mechanical properties of cartilage (Chen et al., 2005; Lefebvre and Dvir-Ginzberg, 2017). The ECM also plays a critical role in regulating chondrocyte proliferation and differentiation. In this study, we have considered the following chondrocyte differentiation markers such as (type II collagen, SRY-box transcription factor 9 (SOX9), and Aggrecans) because type II collagen is a major component of cartilage and is necessary for the proper formation of the ECM in cartilaginous tissues. Studies have shown that type II collagen can enhance GAG synthesis and mRNA expression of Sox9 and aggrecan (Chen et al., 2005; Lefebvre and Dvir-Ginzberg, 2017). SOX9 is a transcription factor that plays a key role in controlling cartilage-specific gene expression. It is required for chondrocyte differentiation and is necessary for the proper formation of cartilage. Studies have also shown that compressive force can promote chondrogenic differentiation (Lefebvre and Dvir-Ginzberg, 2017).
Aggrecan is an integral component of the ECM in cartilaginous tissues, including the growth plate. It is a major component of cartilage and is necessary for the proper formation of the ECM (Hodax et al., 2019). They all play a key role in the formation of the ECM and the proper functioning of chondrocytes.
Arthritis occurs when one or more joints are inflamed, or the cartilage of the joints is destroyed. Arthritis is characterized by structural and biochemical changes in cartilage, including insufficient ECM synthesis, which leads to the deterioration of cartilage substrate and loss of cartilage cell phenotype (Roseti et al., 2019). There are more than 100 types of arthritis; however, representatively, there are two major types, namely osteoarthritis (OA) and rheumatoid arthritis (RA). Among them, OA is the most common form of degenerative joint disease and is among the longest known and most common diseases in humans. In OA, joint tissue is destroyed, leading to joint pain and rigidity as well as pain and swelling in the cartilage. In addition, injury or abnormal mechanical stress can affect the wrist, elbow, shoulder, and ankle.
Inflammatory reactions are also associated with neurodegenerative diseases, cardiovascular diseases, arthritis, cancer, and stroke (Ferrucci and Fabbri, 2018; Furman et al., 2019). Inflammation processes induce basic acute inflammation symptoms and are essentially protective effects that aid in removing harmful factors, healing damaged tissues, and repairing physiological function (Zotova et al., 2016). Inflammatory cells activated during this process secrete large amounts of reactive nitrogen species (RNS), prostaglandin (PGs) E2, and cytokines (interleukin [IL]-1β, IL-6, and tumor necrosis factor-α) (Chen et al., 2018). PGs play an important role in the inflammatory response that is mediated by cyclooxygenase (COX) enzymes in a cell-specific reaction in response to injury or inflammation (Ricciotti and FitzGerald, 2011). COX is an enzyme produced in the biosynthesis of PGs and is also associated with inflammatory diseases such as OA and RA (Cho et al., 2015).
Isomorphic COX-1 and COX-2 of COXs were identified in eukaryotic cells. COX-1 is expressed in most cell types and plays a role in maintaining homeostasis, but COX-2 is mainly detected in a small number of underlying mammalian tissues such as monocytes, fibroblasts, and endothelial cells and is an enzyme derived by inflammatory cytokines, tumor promoters, and growth factors. Inflammatory media such as COXs and PGs affect the temperament homeostasis of articular chondrocytes by varying their metabolism and the activities of MAP kinases, including ERKs, p38, and JNKs, which are important for COX-2 expression (Caron et al., 2016; Eo and Kim, 2020). Mitogen-activated protein kinase (MAPK) is the basis of intracellular signaling that controls cellular functions by combining cell survival and mitosis, apoptosis, gene expression, cellular proliferation, inflammation, and differentiation (Hu et al., 2019; Jie et al., 2021).
Constitutive photomorphogenic 1 (COP1), which was first recognized in Arabidopsis thaliana, is an important protein that mediates the light-response needed for photomorphogenesis in plants (Kang et al., 2021; Subramanian et al., 2004). COP1 is an E3 ubiquitin-linked enzyme that modifies a specific target protein, priming what decomposes through ubiquitin-proteasome systems (Ko et al., 2019). In adults, it is known to function in pathogenic processes such as tumorigenesis (Ka et al., 2018; Song et al., 2020). Moreover, COP1 is also involved in inducing cell growth and differentiation in lung epithelial cells and the loss of this gene averts cell differentiation causing the formation of defective lungs (Zhang et al., 2016). These findings allowed us to investigate the function of COP1 in chondrocyte differentiation that has not been elucidated yet. Thus, this study aimed to elucidate the role of COP1 in mediating inflammatory reactions using rabbit articular chondrocytes.
Materials and Methods
Materials
COP1 cDNA (GFP-tagged) and pCMV-AC6 vector were purchased from Origene Technologies (Rockville, MD). COP1 siRNA was acquired from COSMOGENETECH (Seoul, Korea). cDNA and siRNA transfection was performed using the TurboFect transfection reagent (Fisher Thermo Scientific, Waltham, MA) according to the manufacturer's instructions. Sodium nitroprusside (SNP) and simvastatin (SVT) were purchased from Sigma-Aldrich (St Louis, MO). SB203580 and PD98059 were purchased from Calbiochem (San Diego, CA). Mouse anti-collagen type II monoclonal antibody for western blot analysis and immunofluorescence staining (Cat. No. MAB8887, RRID: AB_2260779) was purchased from MERCK Millipore, Inc. (Merck & Co, NY). Mouse anti-glyceraldehyde 3-phosphate dehydrogenase (GAPDH) monoclonal antibody (Cat No. SC-166545, RRID: AB_2107299), anti-E3 ubiquitin ligase COP1 monoclonal antibody (Cat No. SC-166799, RRID: AB_2178761), rabbit anti-SOX9 polyclonal antibody (Cat No. SC-20095, RRID: AB_1080067), anti-p38 polyclonal antibody (Cat. No. SC-535, RRID: AB_632138), and anti-ERK2 polyclonal antibody (Cat. No. SC-154, RRID: AB_2141292) were purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX). Mouse anti-COX-2 polyclonal antibody for western blot analysis and immunofluorescence staining (Cat. No. 160106, RRID: AB_2571729) was purchased from Cayman Chemical (Ann Arbor, MI). Mouse anti-Aggrecan monoclonal antibody (Cat. No. NB120-11570, RRID: AB_90460) was purchased from Novus Biologicals (CO). Rabbit anti-phospho-p38 (pp38) monoclonal antibody (Cat. No. 9211S, RRID: AB_331640) and anti-phospho-ERK (pERK) polyclonal antibody (Cat. No. 9101, RRID: AB_2315113) were purchased Cell Signaling Technology (Beverly, MA). Anti-mouse IgG antibody (Cat. No. ADI-SAB-100, RRID: AB_11001642) was obtained from Enzo Life Sciences International (Plymouth Meeting, PA). Anti-rabbit IgG antibody (Cat. No. A0545, RRID: AB_257896) from Sigma Aldrich.
Rabbit articular chondrocyte culture
A total of 17 New Zealand rabbits were used from which primary chondrocytes were extracted from articular joints by carving thin slices of cartilage. These cartilages were further processed with the collagenase II enzyme dissolved in Dulbecco's modified Eagle medium (DMEM). Chondrocytes were purified from collagenase treatment lysate with centrifugation and incubated with the culture media. The culture medium was composed of antibiotics (streptomycin and penicillin) at a concentration of 50 μg/mL and 10% (v/v) fetal bovine calf serum dissolved in DMEM. This specialized culture media was further used to facilitate chondrocyte growth and development, and the media were replaced after 48 h to sustain cell confluency as described previously (Shah et al., 2021). All these animal-based experiments were ethically approved by the Chairman of the Institutional Animal Care and Use Committee, Kongju National University, Republic of Korea under the license number (KNU-2022_04).
Western blot analysis
Total protein was harvested from transfected or treated chondrocytes, and western blot analyses were performed. Equal amounts (20–40 μg) of proteins were separated on 9% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to a nitrocellulose membrane. The membrane was blocked with 5% (w/v) nonfat dry milk in Tris-buffered saline (TBS)-0.01% Tween 20 (TBST, pH 8.0) buffer at 25°C for 1 h and incubated overnight with primary antibodies. After washing three times for 10 min with TBS and polysorbate 20 (TBST), the membrane was incubated for 2 h with appropriate secondary horseradish peroxidase–conjugated antibodies. An enhanced chemiluminescence reagent (Dogen, Seoul, Korea) was used to visualize the reactive bands, which were quantified using the LAS4000 camera system (Fuji Film, Tokyo, Japan).
Transfection
The COP1 vector, empty vector, scrambled siRNA, or COP1 siRNA were transfected to chondrocytes using TurboFect Transfection Reagent according to the manufacturer's instructions and left to incubate for 12–24 h depending on the experiment design. COP1 siRNA sequences were as follows: 5′-GCU GUG GUC UAC CAA UCU ATT-3′ (sense), 5′-UAG AUU GGU AGA CCA CAG CTT-3′ (antisense), Negative control 5′-UUC UCC GAA CGU GUC ACG UTT-3′ (sense), 5′-TTA AGA GGC UUG CAC AGU GCA-3′ (antisense). The efficiency of transfection was determined using reverse transcription–polymerase chain reaction (RT-PCR).
Transfected cell treatment
Transfected chondrocytes were treated with 1 mM of SNP, 50 μM of SVT, and 30 μM each of SB203580 (p38 inhibitor) and PD98059 (MEK inhibitor) for 24 h. These concentrations and the duration of SNP, SVT, SB203580, and PD98059 treatment were selected to best represent the effect of each reagent on chondrocytes.
Reverse transcription–polymerase chain reaction
Total RNA was isolated using TRIZOL reagent (Invitrogen Life Technologies, Carlsbad, CA) according to the manufacturer's instructions. Total RNA (0.5 μg) was used for cDNA synthesis with the Maxime RT-PCR PreMix Kit (Intron, Seongnam, Republic of Korea). The following primers and conditions were used for PCR; for COP1 (279-bp product), 5′-TGAGAAGCCGATGTTGGAGTC-3′ (sense) and 5′-GCGTAAGAGACAGCCTTTCG-3′ (antisense) with an annealing temperature 55°C; for type II collagen (370-bp product), 5′-GACCCCATGCAGTACATGCG-3′ (sense) and 5′-AGCCGCCATTGATGGTCTCC-3′ (antisense) with an annealing temperature of 55°C; for SOX-9 (386-bp product), 5′-GCGCGTGCAGCACAAGAAGGACCACCCGGATTACAAGTAC-3′ (sense) and 5′-CGAAGGTCTCGATGTTGGAGATGACGTCGCTGCTCAGCTC-3′ (antisense) with an annealing temperature of 60°C; for COX-2 (299-bp product), 5′-GACCCCATGCAGTACATGGG-3′ (sense) and 5′-AGCCGCCATTGATGGTCTCC-3′ (antisense) with an annealing temperature of 45°C; for GAPDH (299-bp product), 5′-TCACCATCTTCCAGGAGCGA-3′ (sense) and 5′-CACAATGCCGAAGTGGTCGT-3′ (antisense) with an annealing temperature of 50°C. The primers were obtained from Genotech (Daejeon, Republic of Korea). PCR products were resolved on a 1.0% agarose gel and visualized with transilluminator.
Immunofluorescence staining
Chondrocytes were plated on glass coverslips and transfected with COP1 in the absence or presence of inhibitors. Then, the cells were washed three times with phosphate-buffered saline (PBS), fixed with 3.5% (w/v) paraformaldehyde in PBS for 20 min at 25°C in permeabilized with 0.1% Triton X-100 in PBS. The fixed cells were washed three times with PBS and incubated with antibodies against type II collagen and COX-2 overnight at 4°C. The cells were washed three times with PBS and incubated with secondary antibodies for 2 h at 25°C, followed by washing with PBS. Nuclei were stained with 4,6-diamidino-2-phenylindole (DAPI; Invitrogen). The fluorescence images were recorded with a BX51 fluorescence microscope (Olympus, Tokyo, Japan).
Chondrocyte proteoglycan staining
The cells were fixed with 3.5% (w/v) paraformaldehyde in PBS for 20 min and stained with 0.1% Alcian blue in 0.1 M HCl overnight. Chondrocytes were washed three times with distilled water and 6 M guanidine HCl was added for 6 h. Sulfated proteoglycan production was measured at 595 nm using a microplate reader.
Data analysis and statistics
All experiments were repeated thrice, and results are presented as the mean ± standard deviation (SD). Data were assayed with Prism GraphPad 9, and the statistical method used for the analysis of these experiments was the one-way analysis of variance followed by Tukey's post hoc test (p < 0.05) were considered a statistically significant.
Results
COP1 expression impedes differentiation in primary chondrocytes
Primary chondrocytes were used to investigate the role of COP1. After transfection and incubation with COP1 cDNA and siRNA along with control, the cells were harvested and quantified to evaluate the COP1 expression level. Western blotting results showed that upon cDNA transfection, COP1 expression increased, whereas siRNA neutralized the COP1 expression. No expression was observed in the empty vector and negative vector (Fig. 1A). Immunofluorescence was performed and the data showed that COP1 transfection reduced the expression of type II collagen level that propelled us to evaluate the properties of this gene by considering the expression of chondrocyte differentiation and inflammation markers (type II collagen, SOX9, aggrecan, and COX-2).

COP1 expression by COP1 cDNA and siRNA transfection in rabbit articular chondrocytes. Protein expression was identified via western blot analysis and immunofluorescence.
When the cDNA of COP1 was transfected, COX-2 expression increased while type II collagen, SOX9, and aggrecan (chondrogenic markers) decreased. In the case of siRNA transfection restored the expression of chondrogenic genes (Fig. 2A). These results were further analyzed using RT-PCR to confirm and quantify the accurate expression of these genes as observed in our western blot analysis results. The expression of type II collagen, SOX9 was reduced with cDNA transfection while ameliorating COP1 and COX-2 expression. The COP1 siRNA revived the chondrogenic gene expression and suppressed COX-2 expression (Fig. 2B). In Figure 2C, the graph represents the expression of type II collagen and COX-2 as compared with the control. The total proteoglycan synthesis was also assessed using Alcian blue staining. cDNA transfection reduced the proteoglycan synthesis level, whereas siRNA transfection restored it
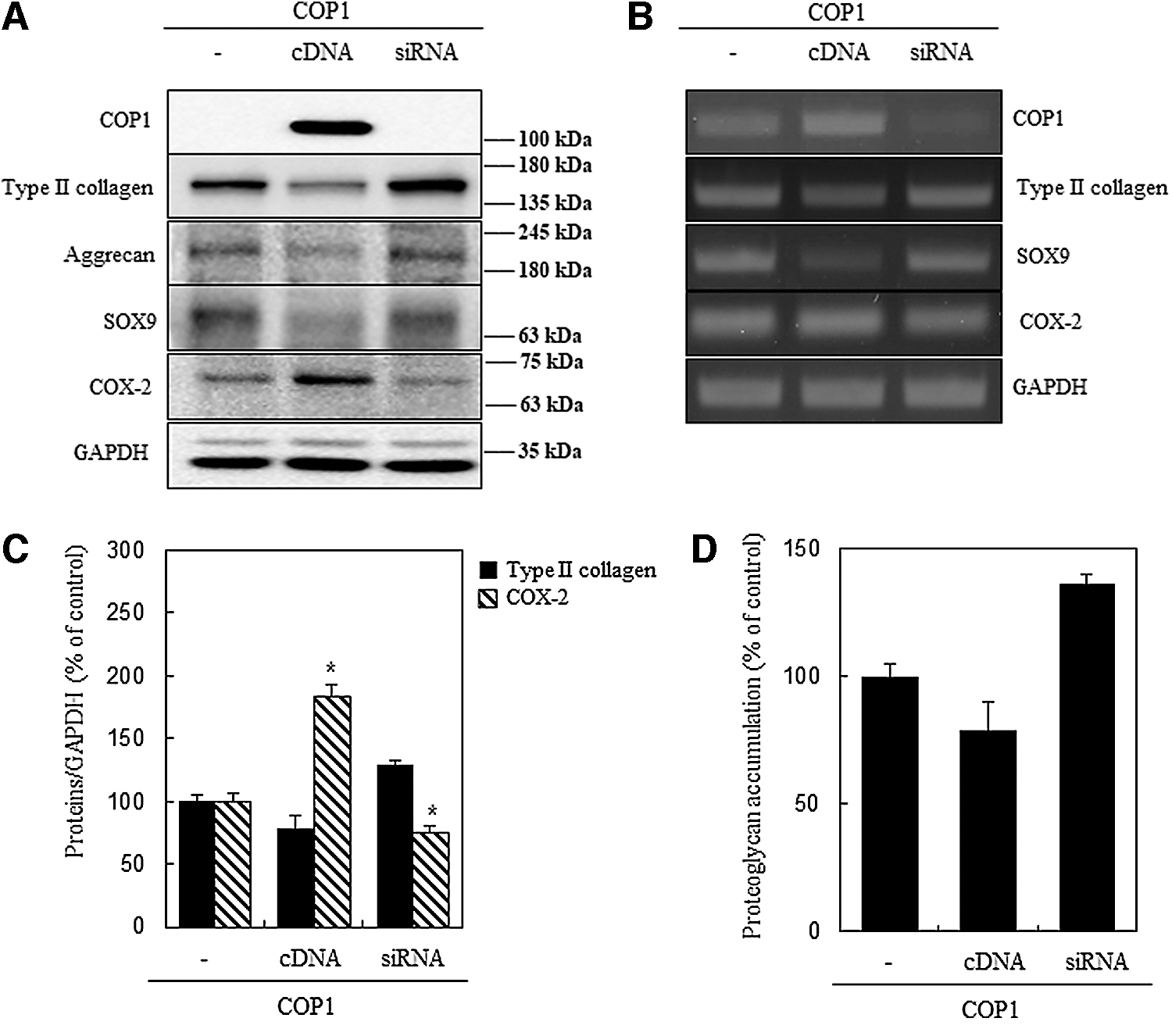
Effects of transfection of chondrocytes with COP1 cDNA and siRNA in rabbit articular chondrocytes.
Chondrocyte differentiation is highly affected by COP1 expression in a time- and dose-dependent manner
In this experiment, we evaluated COP1 cDNA and siRNA expression in a time- and dose-dependent manner. cDNA of COP1 concentration was maintained at 0.7 μg/mL, and its expression was checked at 12 and 24 h while siRNA concentration was 100 nM and its effect was recovered at 12 and 24 h, respectively. COP1 expression drastically increased from 12 to 24 h. The rise in COP1 expression downregulated type II collagen, SOX9, aggrecan, and increased COX-2 expression. siRNA administration sustained chondrogenic gene activity and regulated COX-2
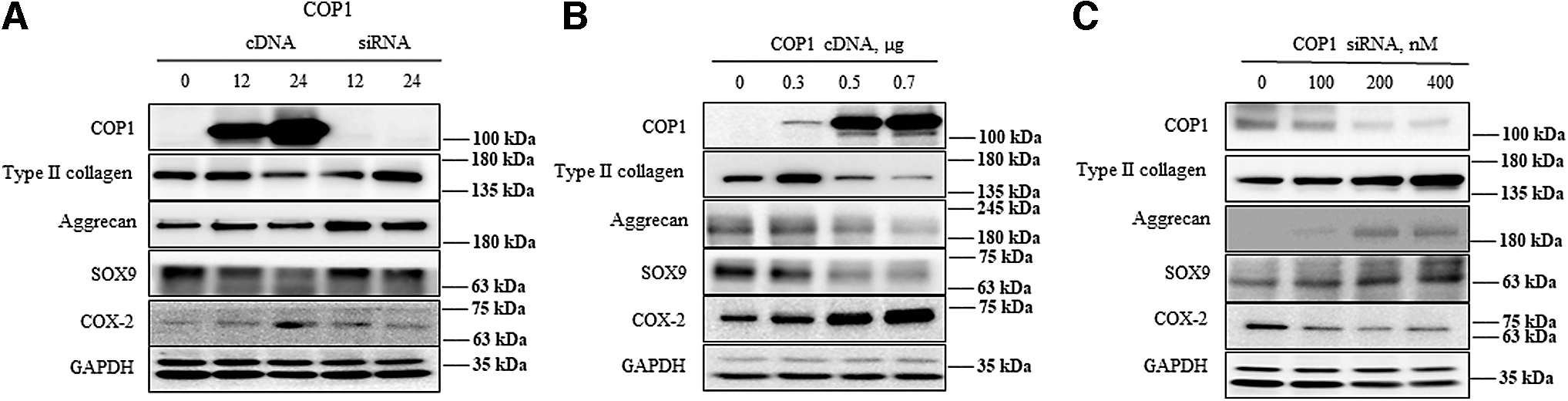
Effects of COP1 cDNA and siRNA transfection in a time- and dose-dependent manner in rabbit articular chondrocytes.
COP1 activity is ameliorated with dedifferentiating agents and negatively affected with differentiating agents
Studies have shown the influence of chondrocyte cellular proteins may intensify upon various drug treatments (Eo and Kim, 2020). Therefore, we noted the activity of COP1 cDNA and siRNA by using SNP and SVT, which are dedifferentiating and differentiating agents of chondrocytes, respectively. When the activity of SNP was evaluated in relation to COP1 cDNA, type II collagen expression substantially decreased along with SOX9 and aggrecan expression, whereas COX-2 activity increased twofold compared with that in the control. COP1 siRNA cotreated with SNP exhibited lowered COP1 expression and promoted type II collagen, SOX9, and aggrecan expression, whereas COX-2 activity remained unchanged compared with the control (Fig. 4A–C). In another experiment, SVT cotreated COP1 cDNA exhibited increased type II collagen, SOX9, and aggrecan expression, whereas siRNA transfection ameliorated the effect of SVT by upregulating type II collagen, SOX9, and aggrecan expression and reducing COX-2 expression (Fig. 4B–D). These experiments show that COP1 activity was antagonized in response to differentiating and dedifferentiating agents in rabbit articular chondrocytes.

COP1 regulates SNP- and SVT-induced chondrogenic genes and COX-2 in rabbit articular chondrocytes.
Differentiation and inflammation were regulated by COP1 through p38 kinase and ERK-1/-2 signaling pathway
The COP1 influence on different chondrocyte pathways was evaluated to determine their role in chondrocyte dedifferentiation. COP1 activity was evaluated in relation to p38 kinase and ERK-1/-2 signaling pathways. Upon transfection, COP1 cDNA induced the expression of phospho-p38 (p38) and pERK (Fig. 5A). These results were further explored by using SB203580 (p38 kinase inhibitor) and PD98059 (MEK inhibitor) along with COP1 cDNA and siRNA of chondrocytes. When cDNA was amalgamated with a p38 inhibitor, type II collagen activity decreases along with COX-2 activity. In contrast, siRNA cotreatment promoted type II collagen activity (Fig. 5B). When combined with an ERK inhibitor, type II collagen level increased with a slight increase in COX-2, whereas ERK expression decreased.
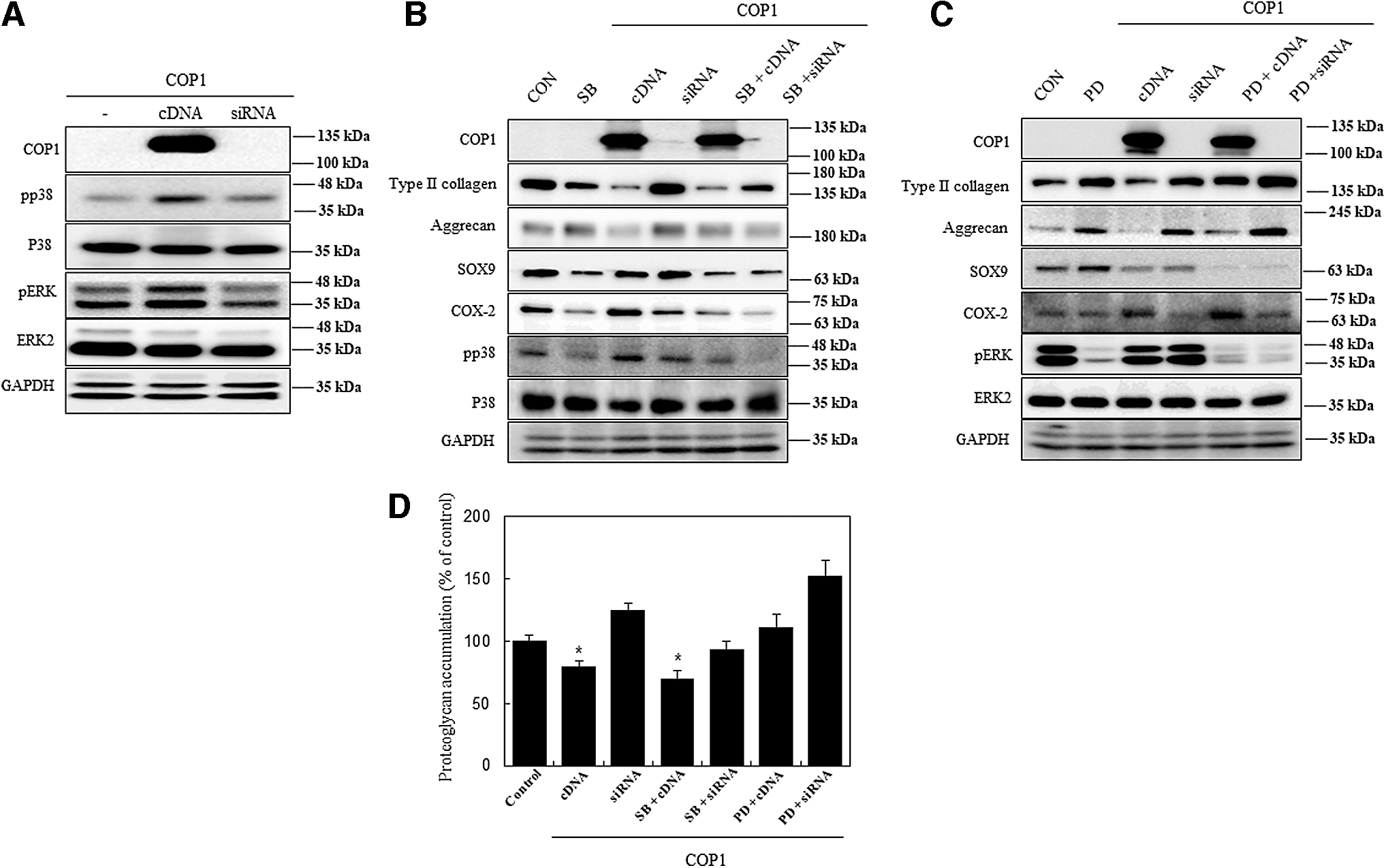
COP1 regulates differentiation and inflammation through p38 kinase and ERK-1/-2 signaling pathways in rabbit articular chondrocytes. Signaling pathways were identified after the transfection of chondrocytes with COP1 cDNA and siRNA.
However, when combined with siRNA of COP1 along with ERK inhibitor drastically upregulate the type II collagen expression while diminishing COX-2 and pERK levels (Fig. 5C). To verify the results, Alcian blue staining was used to quantify proteoglycan synthesis, and the results conformed with that of the western blot analysis (Fig. 5D). Our results show that COP1 causes dedifferentiation of chondrocytes by regulating the p38 kinase and ERK-1/-2 signaling pathway.
Discussion
E3 ubiquitin ligases are notorious protein agents implicated in the proliferation of cancer cells. This protein targets the primary effect of the cell cycle protein known as tumor suppressor protein p53. COP1 belongs to the family of E3 ubiquitin-protein ligase that inhibits the activity of p53 by activating ubiquitination (Ka et al., 2018; Zou et al., 2017). The ubiquitination of p53 causes proteasomal degradation by interacting with other protein targets such as c-Jun, E26 transformation-specific family, metastasis-associated protein-1, and forkhead box 01 (Choi et al., 2020). These protein interactions, along with the influence of COP1, cause the downregulation of p53 expression and protein. Such molecular phenomenon incites rapid and continuous cell proliferation, cell transformation into aggressive phenotype and tumor progression (Song et al., 2020). Moreover, this also aids in reducing cancer cell apoptosis, preventing cell cycle arrest, and promoting mutant cell formation, leading to drug and immunotherapy resistance (Liu et al., 2021).
However, the molecular mechanism of COP1 in relation to chondrocyte differentiation has not been explored. This study determined the role of COP1 and its molecular mechanism in primary chondrocytes obtained from the cartilage of New Zealand white rabbits with respect to differentiation. COP1 transfection in chondrocytes reduced differentiation capability by decreasing the expression of type II collagen, SOX9, and aggrecan but increased COX-2 expression and had not to effect on p53 (data not shown). The siRNA administration of COP1 revived the type II collagen, SOX9, and aggrecan while reducing COX-2 expression; these results were confirmed with RT-PCR, which supported our western blotting results. The Alcian blue staining determines the total proteoglycan levels, which particularly influences chondrocyte differentiation and cartilage health. COP1 overexpression caused a dramatic loss of proteoglycan level in primary chondrocytes.
The experiment suggested that COP1 negatively affects the concentration of proteoglycan level. The expression patterns of cDNA and siRNA of COP1 were studied in a dose- and time-dependent manner, revealing that when 0.7 μg/mL of COP1 is transfected, expression increased at 12 h and increased by twofold at 24 h. The increase in COP1 drastically increased the COX-2 level while reducing chondrogenic gene expression (type II collagen, aggrecan, and SOX9). However, when the concentration of cDNA of COP1 is increased, at 0.5–0.7 μg/mL it completely abrogated the expression of chondrogenic gene expression and increased COX-2 expression. For siRNA, all concentrations ranging from 100 to 400 nM of COP1 positively regulated chondrogenic genes and reduced COX-2 in a time- and dose-dependent manner.
The relationship between type II collagen and COX-2 was visualized with immunofluorescence by evaluating COP1 expression in chondrocytes. It was observed that the level of type II collagen decreases in the chondrocytes as the COX-2 level increases. It has been previously established that treating SNP causes dedifferentiation of primary chondrocytes owing to the formation of RNS (Kim et al., 2002), whereas SVT, a 3-hydroxy-3-methylglutaryl coenzyme A reductase inhibitor induces the chondrogenic gene expression by the WNT/β-catenin pathway (Yu et al., 2019).
When COP1 was cotreated with chondrocyte dedifferentiation reagents such as SNP and differentiation reagent SVT ameliorated the response of COP1 by radically downregulating the chondrogenic expression and increasing COX-2, whereas siRNA slightly revived the type II collagen as compared with the control. A synonymous type of result was observed when cotreated with SVT, as it increased the type II collagen, but twofolds increase was observed when cotreated with siRNA. These findings suggested that even in the presence of differentiation and an anti-inflammatory agent (SVT), COP1 maintains the chondrocyte dedifferentiated phenotype without affecting the inflammation, contrary to SNP, which exacerbates the inflammation. Blocking the activity of COP1 not only deters inflammation but also can rescue the chondrocyte phenotype. However, further studies are required to validate these findings.
The pathway utilized by COP1 was determined to identify the intermediatory agents involved in chondrocyte dedifferentiation. When COP1 was overexpressed, pp38 expression was downregulated and pERK expression was upregulated, whereas siRNA transfection reversed the expression of these proteins. The literature review suggested that both p38 kinase and ERK-1/-2 signaling pathway activation implicated in preventing the expression of chondrogenic genes and inducing chondrocytes dedifferentiation (Eo and Kim, 2020; Rosenzweig et al., 2013). Furthermore, when p38 kinase inhibitor (SB203580) and MEK inhibitor (PD98059) were introduced and cotreated with cDNA and siRNA of COP1, this treatment regulated the effect of dedifferentiation and the expression of chondrogenic genes. Likewise, pathway inhibition was evaluated with cotreatment of SB or PD regulated expression of COX-2 in transfected cells. These results were also supported with Alcian blue staining as they increased the proteoglycan levels upon treatment with p38 kinase and ERK-1/-2 signaling pathway inhibitors along with siRNA of COP1.
Conclusion
This study explored the overexpression role of COP1 protein in disrupting the chondrogenic expression and chondrocyte differentiation. Our results show that COP1 alters the expression of chondrogenic proteins and upregulates the expression of COX-2 and MAPK proteins. Thus, COP1 regulates differentiation and inflammation through the MAPK signaling pathway in rabbit articular chondrocytes. Furthermore, based on the results of our in vitro experiments, further studies need to be performed to establish the link of COP1 with the downstream targets such as ETS variant transcription factor, c-Jun, CCAAT enhancer binding protein (EBP-α and EBP-β) on chondrocyte differentiation to generate concrete evidence about the role of COP1 in OA.
Footnotes
Authors' Contributions
Y.S.E.: investigation and data analysis and curation, B.S.K.: investigation, F.H.S.: writing—original draft, S.J.K.: conceptualization, methodology, investigation, supervision, writing—review and editing.
Ethical Approval
This study is approved by the Chairman of the Institutional Animal Care and Use Committee, Kongju National University, Singwan Campus, Republic of Korea under the license number of KNU-2022_04.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Research Foundation of Korea (NRF) funded by the Korean Government (MEST) (2020R1I1A3B306969913).