
Abstract
The full name of the FTO gene is fat mass and obesity-associated gene. In recent years, it has also been found that FTO is involved in m6A demethylation and regulates the progression of multiple cancers, including gastric cancer. The cancer stem cell theory argues that cancer stem cells are key factors in cancer metastasis, and inhibiting the expression of stemness genes is a good method to inhibit metastasis of gastric cancer. Currently, the role of the FTO gene in regulating stemness of gastric cancer cells is still unclear. By analyzing public databases, it was discovered that FTO gene expression was increased in gastric cancer, and high expression of FTO was associated with poor prognosis of patients with gastric cancer. After gastric cancer stem cells were isolated, it was found that FTO protein expression was increased in gastric cancer stem cells; stemness of gastric cancer cells was reduced after the FTO gene knockdown; subcutaneous tumors of nude mice were smaller than those of the control group after FTO knockdown; and stemness of gastric cancer cells was enhanced after FTO was overexpressed by plasmid. By reviewing additional literature and experimental validation, we found that SOX2 may be the factor by which FTO promotes the stemness of gastric cancer cells. Therefore, it was concluded that FTO could promote the stemness of gastric cancer cells, and targeting FTO may be a potential therapeutic approach for patients with metastatic gastric cancer.
CTR number: TOP-IACUC-2021-0123
Introduction
The full name of the FTO gene is fat mass and obesity-associated gene, which is an obesity-related allele, also known as the obesity gene. In recent years, it has also been found that FTO is involved in m6A demethylation (Abdollahi et al., 2022; Wang et al., 2022). The m6A modification is the most common type of RNA chemical modification, which refers to the methylation modification formed by the 6th nitrogen atom of adenine (A) catalyzed by methyltransferase (Boccaletto et al., 2018; Desrosiers et al., 1974).
m6A is a dynamic and reversible modification that occurs mainly through three types of proteins: the m6A methyltransferases, also known as writers (Cui et al., 2017); the m6A demethylase base transferases, also known as erasers, which include FTO and ALKBH15 (Huang et al., 2015); and the m6A reader genes (Meyer and Jaffrey, 2017). FTO regulates various biological activities in disease progression, such as cancer development, immunity, and tumor progression (Su et al., 2018; Wei et al., 2022; Yang et al., 2019). Some studies have also found that FTO regulates the progression of gastric cancer (Zhang et al., 2022a; Zhou et al., 2022).
Gastric cancer is a common cancer with the fifth highest incidence and fourth highest mortality in the world (Sung et al., 2021). The 5-year survival rate of patients with metastatic gastric cancer is less than 10%, and metastasis of gastric cancer is the main cause of death in patients with gastric cancer (Bouriez et al., 2018; Dassen et al., 2014). Cancer stem cells are a small subset of cells in cancer with stem cell properties, self-renewal capacity, and multicellular differentiation capabilities (Boumahdi et al., 2014; Fu et al., 2019) and they are considered a key factor in cancer recurrence, metastasis, and drug resistance (Chen et al., 2022; Kim et al., 2022; Sher et al., 2022).
Therefore, targeting gastric cancer stem cells is an effective way to inhibit gastric cancer metastasis (Yang et al., 2022). Gastric cancer stem cells are a small fraction of gastric cancer cells with the ability to self-renew and their surface markers are CD44 and CD133 (Chen et al., 2019; Nam et al., 2017; Xu et al., 2019). Su et al. (2020) found that FTO is critical in the self-renewal of leukemia stem cells and they reported two, potent, small-molecule FTO inhibitors with anticancer effects.
SOX2 is thought to be an important transcription factor controlling cancer stem cells (Mirzaei et al., 2022; Rodriguez-Pinilla et al., 2007). Likewise, SOX2 plays an important role in promoting the stemness of gastric cancer cells, for example, Lu et al. (2021) found that SOX2 promotes the stemness and metastasis of gastric cancer cells and Pádua et al. (2020) found that SOX2-enriched gastric cancer cells exhibit cancer stem cell characteristics.
The role of FTO in gastric cancer stem cells has not been reported so far. We determined that FTO can promote the stemness of gastric cancer cells by in vivo and in vitro experiments and provide some information for the treatment of metastatic gastric cancer.
Materials and Methods
Clinical specimens
The prognosis data of patients with gastric cancer were obtained from the Kaplan-Meier Plotter website and the R2 Genomics Analysis and Visualization Platform website from the Amsterdam University Medical Centers, University of Amsterdam and the FTO expression data of patients with gastric cancer were obtained from The Cancer Genome Atlas (TCGA), Gene Expression Profiling Interactive Analysis (GEPIA) website and Gene Expression Omnibus (GEO) databases (GSE54129 and GSE63089).
Plasmids and reagents
The siRNA of FTO was provided by RiboBio (Guangzhou, China), and the targeting sequence is GGATGACTCTCATCTCGAA. Primers for FTO were obtained from RiboBio, sequence (5–3), F: CTGTGCCATTGTGTATGTCTGC, and R: CAC-TTCATCTTGTCCGTTGTAGG. pcDNA3.1 for FTO was purchased from IGE Biotechnology (Guangzhou, China). FTO, SOX2, CD44, and CD133 antibodies were provided by ABclonal (Wuhan, China).
Cell lines and cell culture
SGC7901 and MGC803 were obtained from Cell Type Culture Collection of the Chinese Academy of Sciences (Shanghai, China). DMEM and fetal bovine serum (FBS; Gibco, Logan, UT, USA) were used to culture cells. Cells were mainly cultured in complete medium consisting of 10% FBS and 90% DMEM at 37°C and 5% carbon dioxide concentration. The medium was changed every 2 days.
Cell transfection
The cells were first cultured in six-well plates for 48 h, then the medium was aspirated, and Opti-MEM (Gibco) was added to starve them for 6 h. siRNA (50 nmol/L), pcDNA3.1(1 μg/mL), and Lipofectamine 2000 (Invitrogen, USA) were mixed for 10 min and then added to the cells and incubated for 6 h in the medium. Then, the complete medium was added after the final aspiration, and protein and RNA were extracted after 48 h.
Real-time quantitative PCR
We first added 1 mL of TRIzol, then added 200 μL of chloroform, mixed it well, and centrifuged it. After that we aspirated the supernatant night and added an equal volume of isopropanol, added 75% alcohol to wash, and then dissolved with diethylpyrocarbonate water to obtain RNA. cDNA synthesis was performed with the PrimeScript RT reagent kit (TaKaRa, Dalian, China), followed by amplification with the SYBR Green PCR Master Mix kit (TaKaRa). Actin was used as an internal reference, and the primer sequences are listed in the Plasmids and reagents section.
Western blot analysis
Radio Immunoprecipitation Assay (RIPA) Lysis Buffer (Beyotime Biotechnology, Shanghai, China) was first added to each six-well plate, then the liquid was sucked into an Eppendorf tube and exposed to an ultrasonic wave to disrupt the cells, and the protein supernatant was obtained after centrifugation. After that the protein concentration was measured by the bicinchoninic acid assay method, and the protein concentration of each group was corrected.
After adding the loading buffer (Beyotime Biotechnology), the protein was heated for 10 min and the western blot experiment was performed, the stacking gel and separating gel were prepared, and then protein samples of each group were added. Electrophoresis was performed for 30 min, the voltage was changed to 120 V, the membrane transfer solution was configured, and then the membrane was transferred with the polyvinylidene fluoride membrane.
The current used was 300 mA and the time was 90 min. Afterward, the primary antibody was blocked and incubated overnight and the secondary antibody was incubated the next day and photographs were taken.
Two dimensional culture
A total of 1 × 103 cells were placed in a six-well, low-adherence cell culture plate (Corning; Cat. No. CLS3471). The human epidermal growth factor (EGF; 20 ng/mL; PeproTech, USA), human basic fibroblast growth factor (bFGF; 10 ng/mL; PeproTech), and 4 μg/mL insulin were added to RPMI 1640 (Sigma, USA) to prepare the medium. Cells were grown in the medium for 14 days, after which they were photographed under a microscope, and TRIzol and RIPA were added directly to the six-well plate to obtain the required RNA and protein.
Three dimensional culture
Cells were suspended in 75% of basement membrane extract matrix (1 mg/mL; prepared according to the manufacturer's instructions) and cultured in a six-well, low-adherence cell culture plate (Corning) to prevent them from attaching to the plate. The serum-free 1640 medium was supplemented with 20 ng/mL EGF, 10 ng/mL FGF, and 4 μg/mL insulin.
To extract cells from 3D culture, we disrupted the gels mechanically in cold phosphate-buffered saline, then the spheroids were isolated by centrifugation and incubated in TrypLE Express (Cat. No. 12604-021; Gibco) for 20 to 30 min to generate a single-cell solution. For other specific steps, please refer to our previously published articles (Li et al., 2021).
Animal studies
A total of 1 × 105 MGC803 cells (transfected with siRNA-FTO) were injected subcutaneously into nude mice (4-week-old, female, BALA/c nude mice) that were purchased from the Guangdong Animal Medical Center. Each group comprised four mice. The subcutaneous tumors of nude mice were measured 40 days later, and volume was calculated as follows: V = 1/2 (width2 × length).
Animal care and handling procedures were in accordance with the guidelines and approved by the ethics committee of Shenzhen TopBiotech Co., Ltd., the approval number is TOP-IACUC-2021-0123. Our animal experiments were repeated three times.
Statistical analysis
SPSS 19.0 (IBM, Chicago, IL, USA) and GraphPad Prism 7 (La Jolla, CA, USA) were mainly used to analyze data. Results were compared using Student's t-test, and prognosis was analyzed using the Kaplan–Meier method.
Results
High expression of FTO is associated with poor prognosis in gastric cancer patients, and FTO expression is elevated in gastric cancer patients
The relationship between the expression of FTO and the prognosis of patients with gastric cancer was analyzed using TCGA database through GEPIA, Gene Expression Profiling Interactive Analysis website. It was revealed that high expression of FTO was related to the poor prognosis of patients with gastric cancer (Fig. 1A). The same conclusion was obtained through the R2 database,
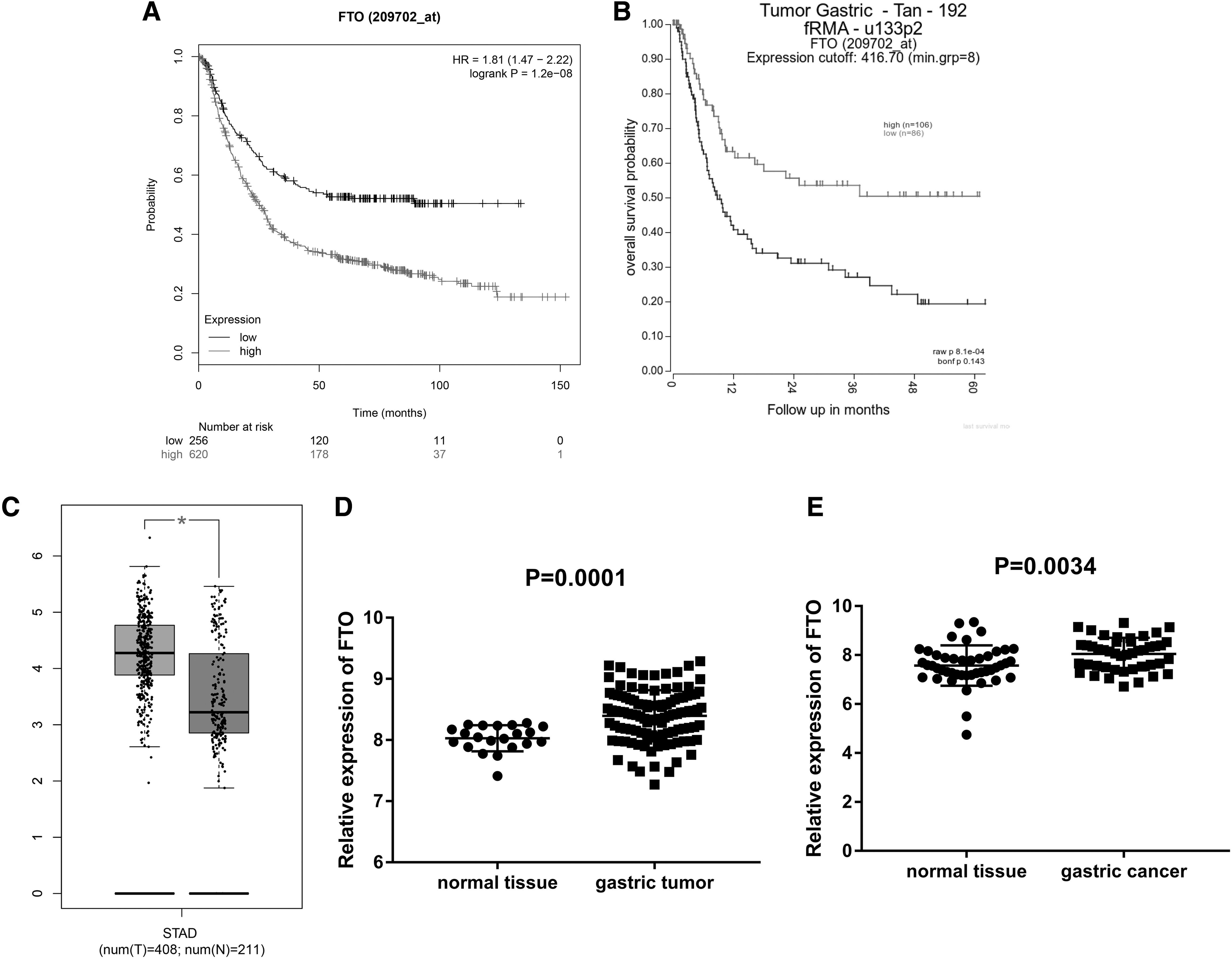
Expression and prognosis of FTO in gastric cancer.
In addition, it was illustrated that the expression level of FTO was increased in patients with gastric cancer in our TCGA database (Fig. 1C). At the same time, it was also found that the expression level of FTO in patients with gastric cancer was increased using GSE54129 and GSE63089 in the GEO database (Fig. 1D, E).
Protein expression of FTO was increased in gastric cancer stem cells
Gastric stem cell-like cancer cells were isolated by 2D and 3D culture, and expression of the FTO protein was detected by western blot experiment. It was found that expression of the FTO protein in 2D isolated cells of the SGC7901 cell line was higher than that of SGC7901 bulk cells and expression of the FTO protein in the 3D isolated cells of the SGC7901 cell line was higher than that of SGC7901 bulk cells (Fig. 2A, B).
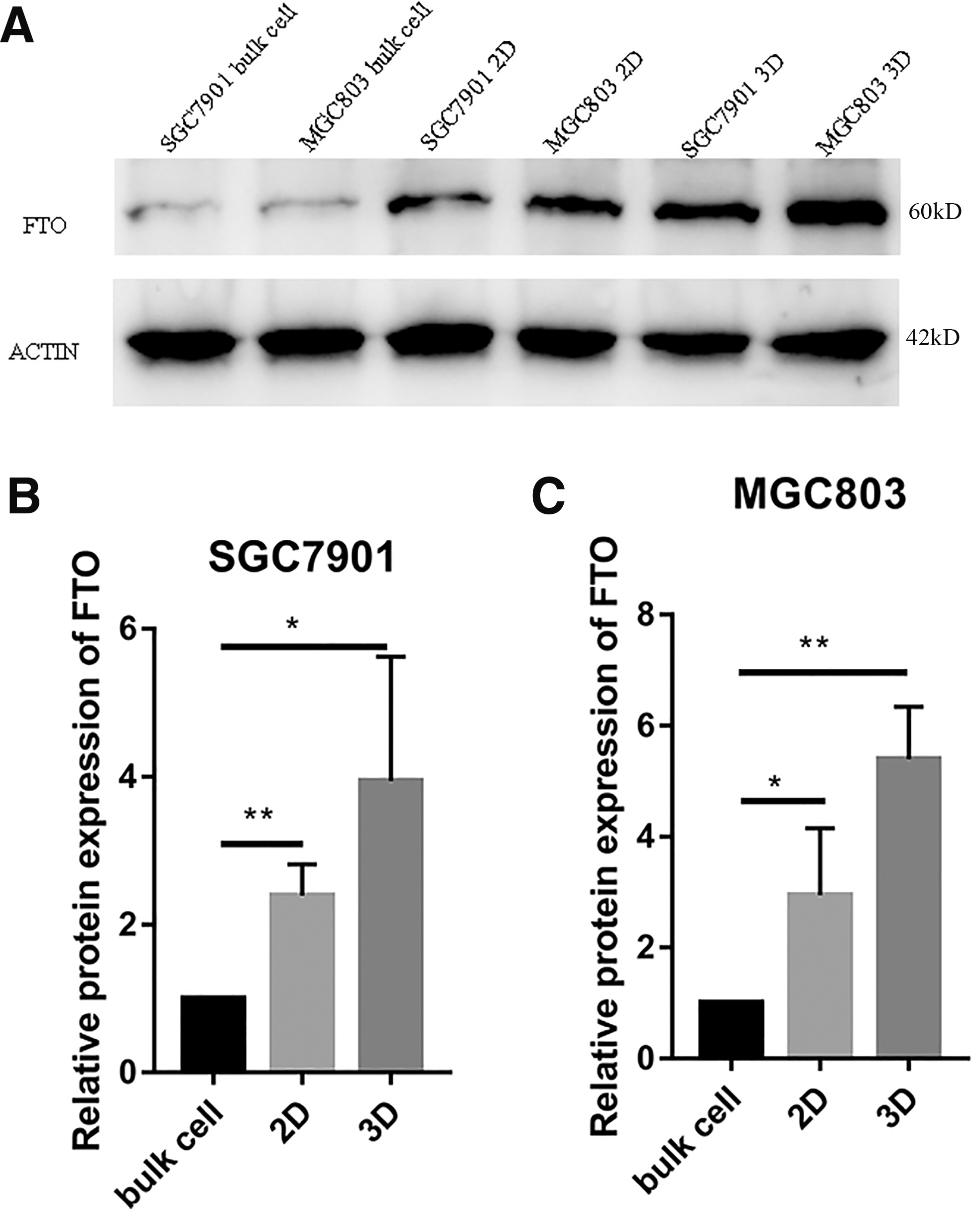
Expression of FTO protein in gastric cancer stem cells.
In addition, it was discovered that expression of the FTO protein in 2D isolated cells of the MGC803 cell line was 2.94 ± 0.6982 times that of MGC803 bulk cells and expression of the FTO protein in the 3D isolated cells of the MGC803 cell line was 5.39 ± 0.5465 times that of MGC803 bulk cells (Fig. 2A, C).
Knockdown of FTO inhibits the stemness of gastric cancer cells
FTO in SGC7901 and MGC803 cell lines was knocked down by siRNA, and it was found that mRNA expression of FTO in the SGC7901 cell knockdown group was lower compared with the control group and mRNA expression of FTO in the MGC803 cell knockdown group was lower compared with the control group (Fig. 3A); protein expression of FTO also decreased after FTO knockdown (Fig. 3B).

Knockdown of FTO inhibits the stemness of gastric cancer cells.
It was found that the number of spheroids in 2D and 3D cell cultures of SGC7901 was reduced after knockdown of FTO expression (Fig. 3C), which was the same in MGC803 cells (Fig. 3D). After knockdown of FTO, the expression level of the CD44 protein in SGC7901 was 0.5667 ± 0.06692 times that in the control group, the protein expression level of CD44 in MGC803 was 0.5267 ± 0.0318 times that in the control group, the protein expression level of CD133 in SGC7901 was 0.58 ± 0.03786 times that in the control group, and the CD133 protein expression of MGC803 was 0.5567 ± 0.07881 times that in the control group (Fig. 3E).
Knockdown of FTO inhibits tumor growth in vivo
A total of 1 × 105 MGC803 cells of the FTO knockdown group and control group were injected subcutaneously into nude mice for 40 days, then the tumor sizes in the FTO knockdown group and control group were measured. The tumor size was 862.4 ± 129.7 mm3 in the control group and 214.6 ± 40.59 mm3 in the FTO knockdown group (Fig. 4A, B).
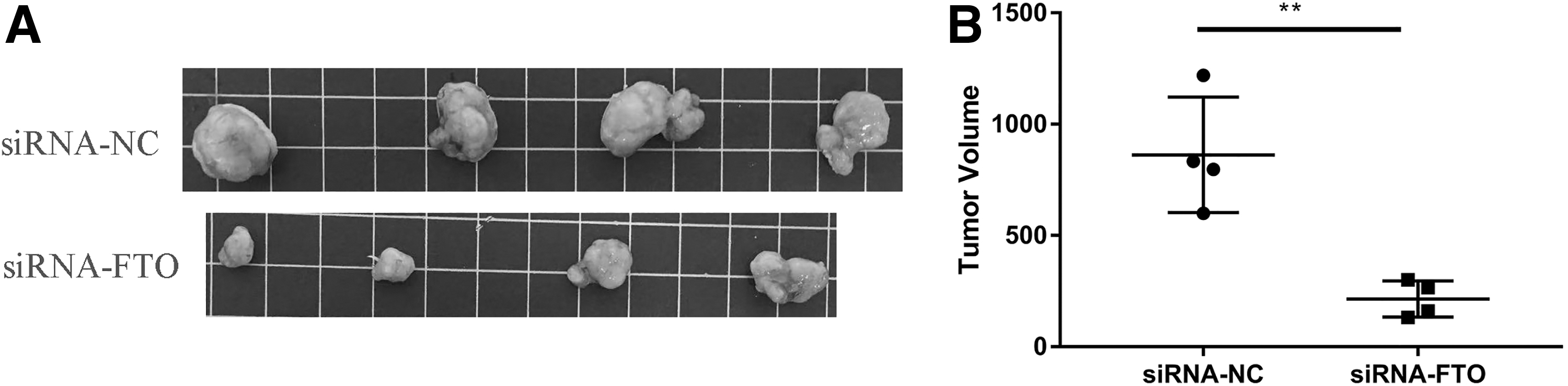
Knockdown of FTO inhibits tumor growth in vivo.
Overexpression of FTO promotes the stemness of gastric cancer cells
By adding the pcDNA3.1 of FTO and the control plasmid into SGC7901 and MGC803 cells, it was found that the mRNA expression of FTO in the overexpression group was higher compared with the control group for SGC7901 cells, and the mRNA expression of FTO in the overexpression group was higher compared with the control group for MGC803 cells (Fig. 5A); protein expression of FTO was also increased after overexpression of FTO (Fig. 5B).
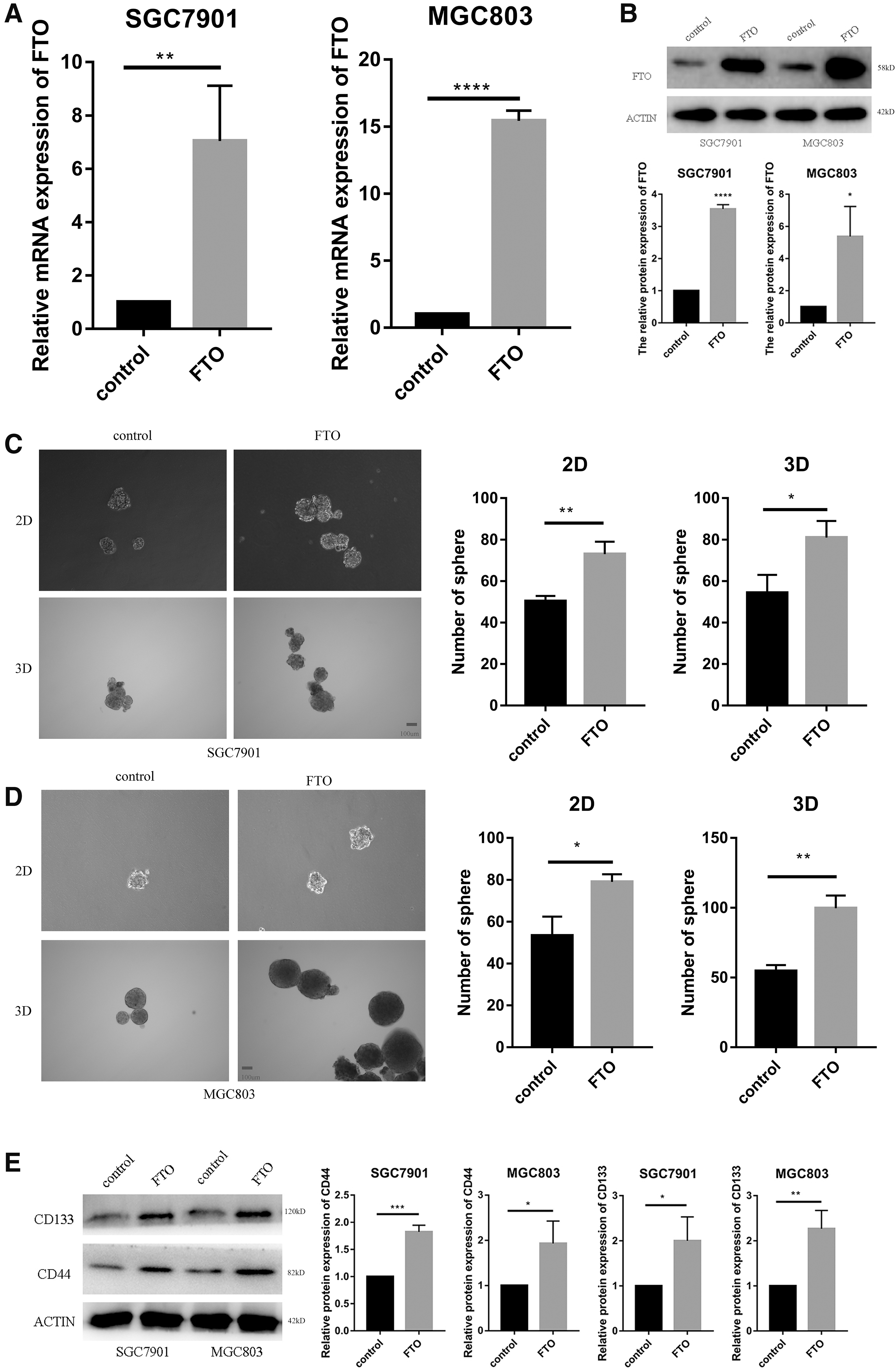
Overexpression of FTO promotes the stemness of gastric cancer cells.
The number of 2D and 3D spheroid cells of SGC7901 and MGC803 cells was higher relative to the control group after overexpression of FTO (Fig. 5C, D). After overexpression of FTO, the expression of the CD44 protein of SGC7901 was 1.823 ± 0.06984 times that in the control group; the expression of the CD44 protein of MGC803 was 1.933 ± 0.2849 times that in the control group; the expression of the CD133 protein of SGC7901 was 2 ± 0.3055 times that in the control group; and the CD133 protein expression of MGC803 was 2.267 ± 0.2333 times that in the control group (Fig. 5E).
FTO regulates the expression of SOX2 in gastric cancer cells
In the study by Li et al. (2017), the authors performed FTO overexpression in leukemia cells, followed by sequencing, and found changes in the expression of some genes. We conjectured that there might be a similar regulatory mechanism of FTO in gastric cancer. We performed PCR of several genes after overexpression of FTO and found changes in SOX2 expression and then using Western Blot (WB) experiments, we further verified that FTO could regulate SOX2 expression changes in gastric cancer cells.
We can see from the figure below that the expression of SOX2 mRNA was greatly reduced after knockdown of FTO (Fig. 6A), while the opposite is true after overexpression of FTO (Fig. 6B). Similarly, the protein expression of SOX2 decreased after FTO knockdown and increased after FTO overexpression (Fig. 6C, D).
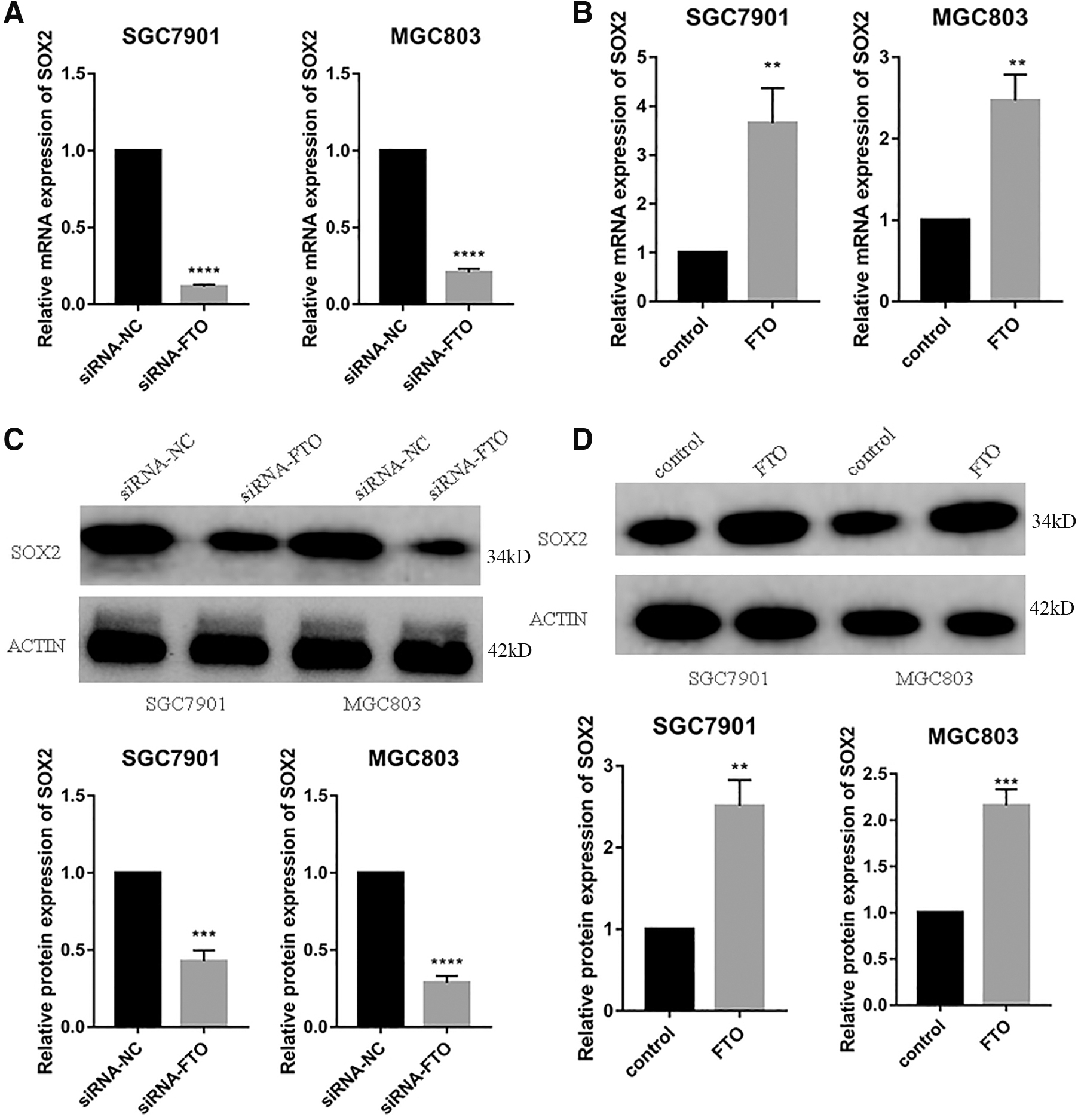
FTO promotes the expression of SOX2.
Discussion
Gastric cancer is a very common cancer and metastatic gastric cancer is the main cause of death in gastric cancer patients. The 5-year survival rate of gastric cancer patients with metastases is less than 10%, therefore inhibiting gastric cancer metastasis is an important method to improve the prognosis of gastric cancer patients (Kanda et al., 2020; Shibamoto et al., 2022). Gastric cancer stem cells are a key factor in gastric cancer metastasis, and studies have found that inhibiting gastric cancer stemness is the way to inhibit gastric cancer metastasis (Becerril-Rico et al., 2021; Ni et al., 2021).
For example, Pajuelo-Lozano et al. (2020) found that targeting MAD2 inhibits gastric cancer metastasis by inhibiting gastric cancer cell stemness. The FTO gene is the fat mass and obesity-associated gene; in recent years, it has been found that FTO participates in m6A demethylation and it also participates in cancer stemness and regulates the progression of gastric cancer (Jing et al., 2021; Shimura et al., 2022; Zhang et al., 2022b). However, the role of FTO in gastric cancer stem cells is currently unclear.
In general, the signaling pathways involved in the progression of cancer stem cells are the WNT signaling pathway, NOTCH signaling pathway, and Hedgehog signaling pathway, and it is the same in gastric cancer stem cells (Bhal and Kundu, 2023; Köseer et al., 2023; Ma et al., 2023; Yang et al., 2022). Inhibitors targeting these signaling pathways, such as vantictumab, MK0752, and vismodegib, may treat cancer by inhibiting cancer cell stemness (Rao et al., 2022).
Through TCGA and R2 databases, we found that the high expression of FTO is related to the poor prognosis of gastric cancer patients, and through TCGA and GEO databases, we found that the expression of FTO was increased in gastric cancer patients. We isolated gastric cancer stem cells by 2D and 3D cultures. In addition, using the WB experiment, we found that the protein expression of FTO was increased in gastric cancer stem cells (Li et al., 2021).
After knockdown of FTO, the stemness of gastric cancer cells was reduced; experimental results showed that the number of spheroid cells in 2D and 3D cultures was reduced; and the protein expression levels of CD44 and CD133, the markers of gastric cancer stem cells, were reduced (Sentani et al., 2021). Besides, it was also discovered that after knockdown of FTO, the tumor volume was reduced in nude mice.
We found several potential FTO downstream genes by reading other literature (Pajuelo-Lozano et al., 2020), and PCR screening showed that there were differential changes in SOX2. We later verified the protein expression of SOX2 and got the same results. Finally, it was found that overexpression of FTO promotes the stemness of gastric cancer cells and the mechanism may be that FTO affects the expression of SOX2.
Collectively, we found that FTO promotes the stemness of gastric cancer cells. Cui et al. (2017) also found that FTO promotes the progression of cancer stem cells. Huang et al. (2019) designed two small-molecule inhibitors to inhibit acute myeloid leukemia by targeting FTO, confirming that targeting FTO to inhibit cancer is feasible. Su et al. (2020) designed two small-molecule inhibitors targeting FTO to inhibit cancer stem cell maintenance. Bian et al. (2021) found that FTO promotes the self-renewal of liver cancer stem cells.
These findings provide a research basis for targeting FTO for cancer therapy, and our findings also provide new information for the treatment of metastatic gastric cancer. Of course, some studies have found that FTO inhibits the stemness of cancer cells. Huang et al. (2020) found that FTO inhibited the self-renewal of ovarian cancer stem cells. Relier et al. (2021) found that FTO inhibits the stem cell properties of colorectal cancer. The occurrence of these phenomena may be due to the different roles of FTO in different cancers.
Most importantly, our research is still insufficient, and the specific downstream molecular mechanism has not been further studied. The best way to detect whether FTO affects the m6A level of SOX2 is by MeRIP-pcr, which provides the most direct evidence. In addition, we did not explore whether FTO promotes the stemness of gastric cancer by activating the WNT signaling pathway, NOTCH signaling pathway, or Hedgehog signaling pathway. In the future, we need to further study its molecular mechanism to provide more detailed information.
In conclusion, we found that FTO promotes the stemness of gastric cancer cells, providing new information for the treatment of metastatic gastric cancer.
Footnotes
Authors' Contributions
G.L.,
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Shenzhen Sanming Project (SZSM201612041) and Shenzhen Science, Technology and Innovation Commission Project (GJHZ20180754917, ZDSYS201909020 92855097).