
Abstract
Ewing sarcoma family tumors (ESFTs) are a group of aggressive tumors mainly affecting children and young people. A compound derived from Curcuma wenyujin plant or lemon grass, β-elemene, has exhibited antitumor effects to ESFT cells, the mechanism of which remains to be clarified further. Autophagy is involved in the antitumor effects of various drugs, whereas the role of autophagy in the antitumor effects of β-elemene persists controversial. Herein we found that β-elemene treatment inhibited the viability of ESFT cells in a dose-dependent manner. The increase of LC3-II level and the decrease of p62 level were observed in β-elemene-treated cells, as well as the increase of autolysosomes, which indicated the promotion of autophagic flux. Sequentially the autophagy inhibition using 3-MA treatment or ATG5 depletion significantly reversed the viability repression and apoptosis induction by β-elemene treatment. In addition, autophagy was found to be important in the toxic effects induced by the combination treatment of β-elemene and IGF1R inhibition in ESFT cells. Our data suggested an essential role of autophagy in β-elemene-induced apoptosis in ESFT cells, which is anticipated to provide novel insights to the development of ESFT treatments.
Introduction
Ewing sarcoma family tumors (ESFTs) are a group of highly progressive and poorly prognostic malignant tumors affecting children and young adults (Riggi et al., 2021). Although the 5-year relative survival rate for all cancers combined in USA was improved to 85% during 2011 through 2017 in children and 86% in adolescents, the 5-year relative survival of ESFTs is only 70–76% in children and 50–59% in adolescents (Siegel et al., 2022). Currently the treatments for ESFTs comprise chemotherapy (McTiernan et al., 2006), radiotherapy (Foulon et al., 2016), and surgery plus radiotherapy (Ahmed et al., 2017), the effects of which are very limited.
Recently, various immunotherapies and molecular targeted therapies (Guerzoni et al., 2015; Heitzeneder et al., 2019; Iniguez et al., 2018; Italiano et al., 2020; Juergens et al., 2011; Kailayangiri et al., 2017; Kelleher and Thomas, 2012; Naing et al., 2012; Sun et al., 2016) of ESFTs including IGF1R inhibition have been studied, whereas the clinical application remains to be clarified. In addition, the possibility of drug resistance and disease recurrence is concerned. Therefore, novel safe and effective approaches are needed urgently.
β-elemene (1-methyl-1-vinyl-2,4-diisopropenyl-cyclohexane) originates from Curcuma wenyujin Y. H. Chen & C. Ling, a traditional Chinese medicinal herb. C. wenyujin has been applied to many clinical events, including treating cancer (Fu, 1984; Li et al., 2021; Yin et al., 2012). The traditional use of β-elemene is to target against a broad spectrum of tumors (Fu, 1984; Jiang et al., 2017; Li et al., 2021; Ma et al., 2016; Wang et al., 1996), and is often used to treat tumors clinically in China (Tan et al., 2000). Lots of studies show in many kinds of cancer cells that β-elemene treatment induces apoptosis and cell cycle arrest, reverses drug resistance, enhances drug effects, and so on (Chen et al., 2012; Dai et al., 2013; Li et al., 2005; Zhao et al., 2015; Zhou et al., 2016; Zhu et al., 2011). We previously reported that β-elemene treatment was able to induce apoptosis of ESFT cells through targeting the phosphorylation of insulin receptor (Wu et al., 2019). However, the mechanisms in detail need to be further elucidated.
Autophagy is a highly conserved process from yeast to mammals, transporting damaged organelles and denatured proteins to lysosomes for enzymatic hydrolysis. Autophagy process includes the following stages: formation of phagophore, autophagosome, autolysosome, and degradation (Klionsky et al., 2021; Lamb et al., 2013). Although autophagy can promote cell growth and maintain life, extensive or persistent autophagy will also induce cell death (Cuervo, 2004; Denton and Kumar, 2019; Doherty and Baehrecke, 2018).
Thus, autophagy is a double-edged sword facilitating cell death or survival, and the disruption of autophagy leads to the pathological processes of various diseases, such as cancer and inflammatory diseases (Dikic and Elazar, 2018; Levine and Kroemer, 2019). Although inhibition of autophagy is able to enhance the antitumor effects of several chemotherapeutic drugs (Liu et al., 2020; Zhang et al., 2020; Zhang et al., 2014), autophagy has been reported to mediate the cell death induced by some antitumor medicine (Puissant et al., 2010).
The effects of β-elemene on autophagy remain controversial. β-elemene treatment inhibited autophagic flux to reverse gefitinib resistance in NSCLC cells, whereas induced autophagy to reverse the resistance of colorectal cancer cells or to protect from apoptosis in various cancer cells (Guan et al., 2014; Lin et al., 2014; Liu et al., 2020; Liu et al., 2012; Liu et al., 2011; Zhang et al., 2020). In this study, we report that β-elemene treatment induces autophagy-mediated apoptosis in EFST cells. Moreover, β-elemene treatment shows toxic effects in combination with the IGF1R inhibitor through facilitating autophagy. These data have provided a novel insight to develop novel therapeutic approaches for treatment of ESFTs.
Materials and Methods
Cell culture
Human ESFT cell line A673 was obtained from ATCC, and MHH-ES-1 was obtained from Deutsche Sammlung von Mikroorganismen und Zellkulturen (DSMZ) and cultured in DMEM supplemented with 10% fetal bovine serum (FBS). They were kept in a 5% humidified CO2 incubator at 37°C.
Reagents
β-elemene (95%) was obtained from Yuanda Pharmaceuticals (Dalian, China). PPP (S7668) was purchased from Sellek. Z-VAD (HY-16658) and 3-MA (HY-19312) were purchased from MedChem Express (Monmouth Junction, NJ). CQ (C6628) was purchased from Sigma-Aldrich.
Plasmids and transfection
The human pCDH-CMV-mRFP1-EGFP-LC3B-EF1-Puro plasmid (P4838) was purchased from Miaolingbio. The psPAX2 (Addgene plasmid#12260) and pMD2.G (Addgene plasmid#12259) were gifts from Didier Trono. pLKO.1 was a gift from David Root (Addgene plasmid#10879) (Moffat et al., 2006). shATG5 sequence 5′-GGATGCAATTGAAGCTCAT-3′ was cloned into pLKO.1. mRFP1-EGFP-LC3B or shATG5, psPAX2, together with pMD2.G plasmids were transfected into 293T cells with transfect regents polyethyleneimine (24765; Polysciences). Twenty-four hours later, virus was collected to infect the cells at the presence of 8 μg/mL Polybrene (H9268; Sigma). Seventy-two hours later the cells were incubated with 2 μg/mL puromycin (P8833; Sigma) to establish stable cell lines.
Cell viability analysis
Cells were seeded at 1 × 104 cells per well into 96-well plates and treated with different concentration of β-elemene combined with or without inhibitors for 24 or 48 h. Cell viability was evaluated with CCK-8 (CK04; Dojindo) according to the manufacturer′s protocol.
Transmission electron microscopy
Cell pellets were fixed in 2.5% glutaraldehyde and 1% osmium tetroxide (OsO4) sequentially. Sections (80 nm) of cell pellets were obtained after embedded in reductive resin. The sections were stained with uranyl acetate and lead nitrate, and observed under TEM (JEM-2000EX).
Apoptosis analysis
Cells were seeded at 1 × 106 cells into 21 cm2 dishes and treated with different concentration of β-elemene combined with or without 3-MA for 24 h. Apoptosis was measured with Annexin V-FITC Apoptosis Detection Kit (AD10; Dojindo). The cells were collected and washed with cold PBS, and stained simultaneously with Annexin V-FITC and PI. The cells were analyzed using Accuri C6 (BD).
Antibodies and Western blot analysis
The relevant antibodies were purchased as follows: β-actin (20536-1-AP; Proteintech); PARP (9532; CST); LC3 (12741; CST); p62 (ab56416; Abcam). Western blot analysis was performed according to the protocol (Huang et al., 2014), and the protein level was determined using Chemistar™ High-sig ECL Western Blotting Substrate (180-5001; Tanon). Images were captured with an Image Analyzer ChemiDoc XRS+ (BioRad).
Monitoring autophagy flux
mRFP-EGFP-LC3 transfected cells were treated with β-elemene, CQ or β-elemene together with CQ for 24 h. Images were obtained by a confocal microscope (Leica TCS SP5 × ). RFP+EGFP+ LC3 puncta (yellow dots) represented autophagosomes, whereas RFP+EGFP− LC3 puncta (red dots) represented autolysosome (Hariharan et al., 2011; Klionsky et al., 2021).
Statistical analysis
The data were shown as the mean ± SD with at least three independent experiments. Differences between groups were evaluated with Student's t-test. The statistical significance in the figures was set at *p < 0.05, **p < 0.01, and ***p < 0.001. GraphPad Prism 5 software was used for statistical analyses.
Results
β-elemene treatment induces apoptosis and influences autophagy in ESFT cells
We first evaluated the effects of β-elemene treatment on ESFT cell lines (Fig. 1A–E). The results showed that the viability of both A673 and MHH-ES-1 cells was suppressed by the 24-h treatment dose-dependently (Fig. 1B, C). Simultaneously apoptotic bodies were observed under TEM (Fig. 1D). The Western blot results exhibited that the level of an apoptosis marker, cleaved-PARP, was increased remarkably by β-elemene treatment (Fig. 1E). The data suggested that β-elemene induced apoptosis in ESFT cells.
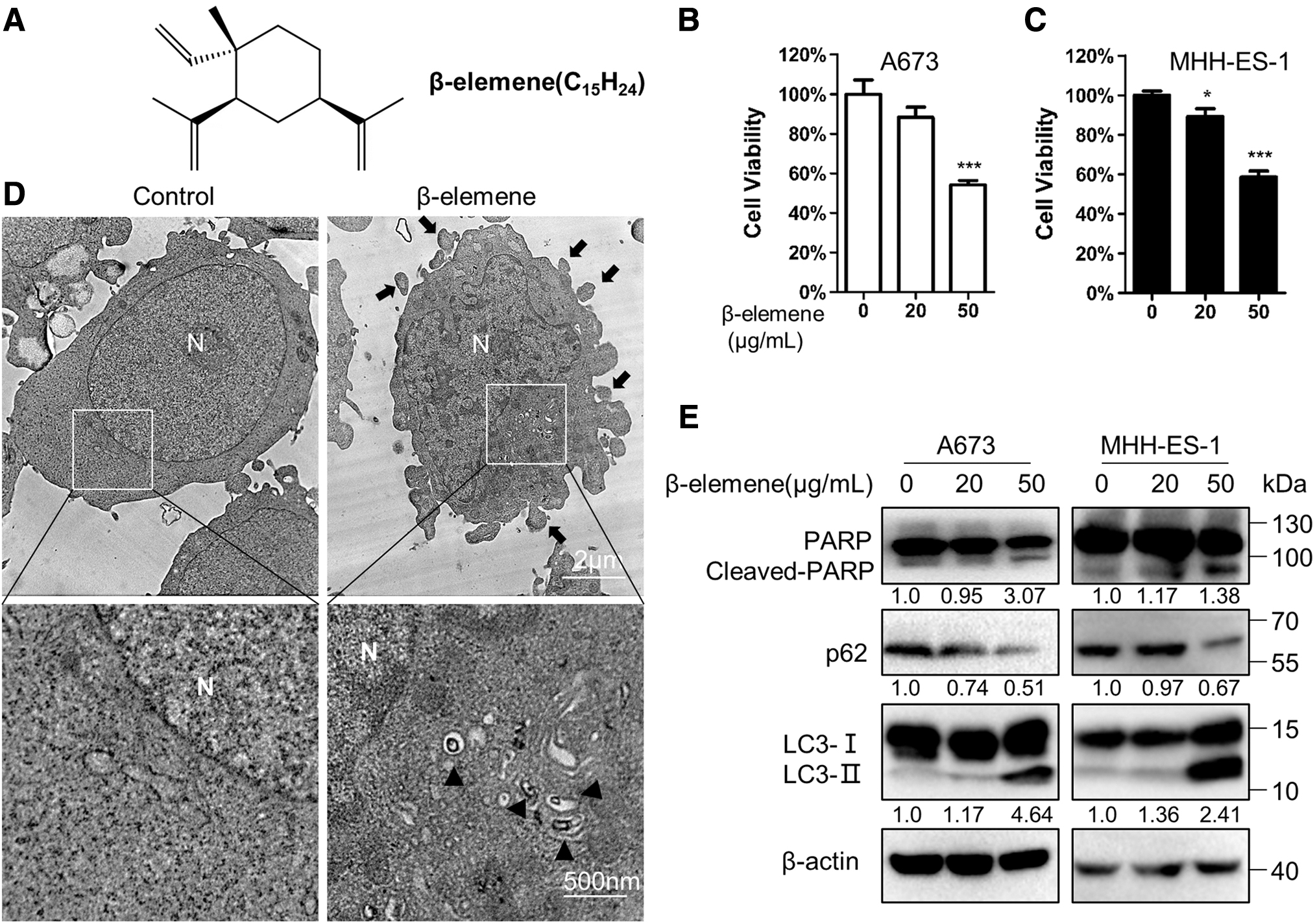
Effects of β-elemene treatment on A673 and MHH-ES-1 cells.
In addition, we found that autophagic bodies were increased significantly after β-elemene treatment (Fig. 1D). The autophagy marker LC3-II was greatly increased along with cleaved-PARP, whereas the level of a selective autophagic substrate, p62, was downregulated significantly in β-elemene-treated cells (Fig. 1E). These data indicated that β-elemene treatment influenced autophagy in ESFT cells.
β-elemene treatment facilitates autophagic flux
The level of LC3-II could be increased by both induction of autophagosome formation and inhibition of autophagosome degradation. To clarify the cause of LC3-II accumulation further, chloroquine (CQ) was used to inhibit the fusion of autophagosome with lysosome. As the results, the decline in p62 level caused by β-elemene treatment was impaired by the combination with CQ treatment, whereas the LC3-II accumulation induced by β-elemene treatment was further enhanced by CQ treatment (Fig. 2A). These data suggested that β-elemene facilitated autophagosome formation and autophagic flux.
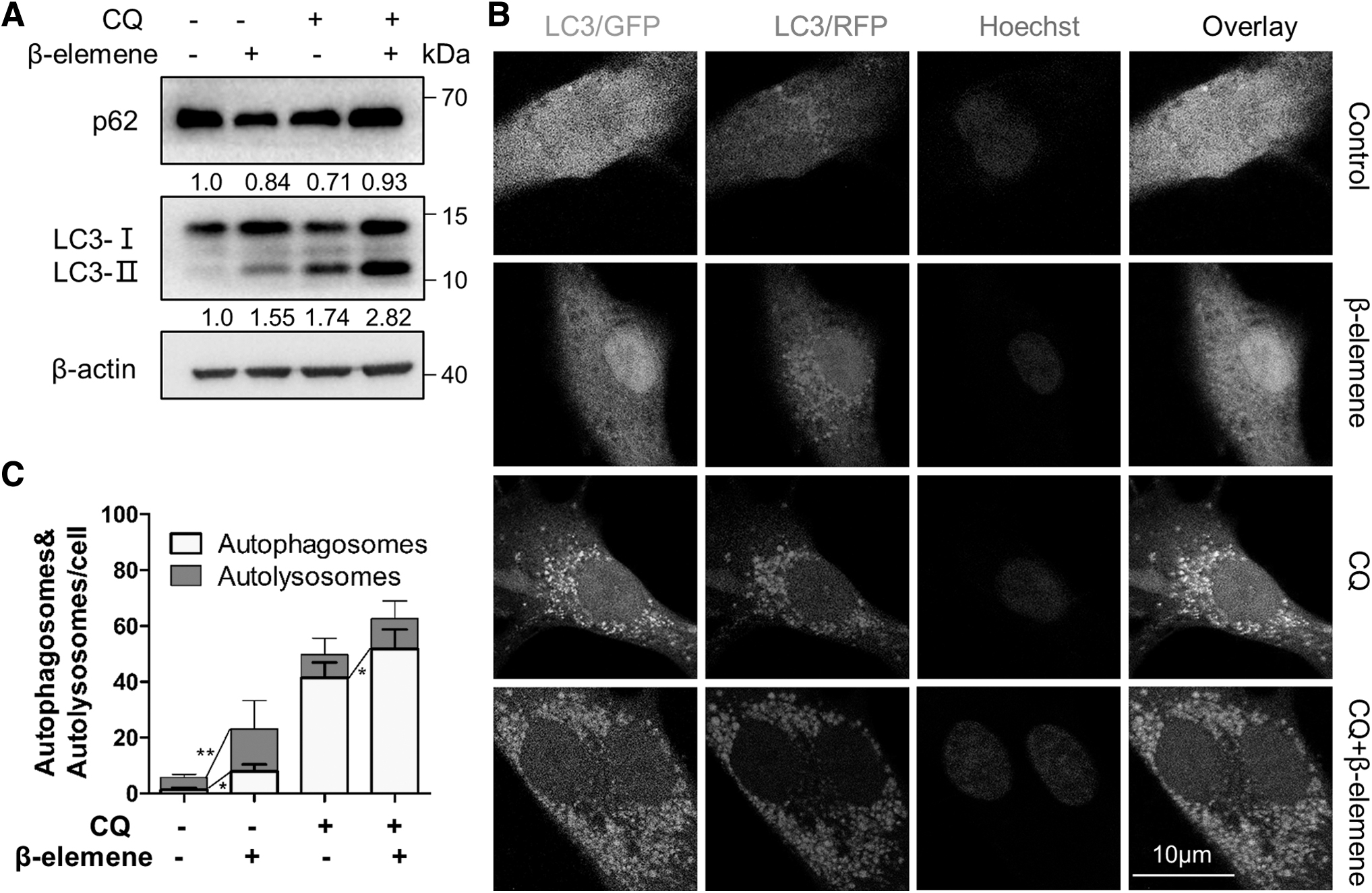
Effects of β-elemene on autophagy. A673 cells were exposed to the indicated treatments for 24 h.
We next introduced the mRFP-EGFP-LC3 reporter into A673 cells and analyzed autophagic flux further through observing LC3 punta under confocal microscopy. In these cells, autophagosomes (mRFP+EGFP+LC3 puncta, yellow dots) show both mRFP and EGFP signals, whereas autolysosomes (mRFP+EGFP− LC3 puncta, red dots) only exhibit mRFP signal due to the quenching of EGFP signal in the acidic lysosomal environment (Klionsky et al., 2021). The results showed that β-elemene treatment increased the number of autophagosome from 1.4 to 7.2 per cell and autolysosomes from 4.4 to 15.2 per cell compared with untreated (control) cells (Fig. 2B, C). In addition, CQ treatment resulted in more autophagosomes in β-elemene-treated cells, indicating that β-elemene treatment mainly promotes autophagosome formation instead of impairing degradation. The autophagic flux was significantly stimulated by β-elemene treatment.
β-elemene treatment triggers autophagy-mediated apoptosis
To explore the relationship of the autophagy and the apoptosis induced by β-elemene treatment, we treated A673 cells using an autophagy inhibitor 3-Methyladenine (3-MA), which represses autophagosome formation together with β-elemene for 24 or 48 h. Cell viability assays were performed, and the results showed that 3-MA treatment significantly reversed β-elemene-repressed cell viability (Fig. 3A). Moreover, apoptosis assays showed similar results. The ratio of PI-/Annexin V+cells representing early apoptosis was increased to 23.9% by β-elemene treatment from 1.32%, which was restored to 9.10% by the combination with 3-MA; whereas the ratio of PI+/Annexin V+cells representing late apoptosis was increased to 9.29% by β-elemene treatment from 3.09%, which was restored to 3.90% by the combination with 3-MA (Fig. 3B). Therefore, autophagy is necessary for the apoptosis induced by β-elemene treatment.

Effects of autophagy on the apoptosis induced by β-elemene treatment.
We next depleted ATG5 (shATG5) to inhibit autophagosome formation. Accordingly, the difference in the cellular viability between β-elemene-treated and control cells was significantly diminished (Fig. 3C). The viability was repressed by β-elemene treatment in control cells; meanwhile, the ATG5-depleted cells treated with β-elemene showed higher viability than the control cells. Moreover, as shown in Figure 3D, the levels of LC3-II, p62, and cleaved PARP altered in β-elemene-treated cells were reversed significantly after ATG5 depletion. These data suggest that β-elemene treatment induces apoptosis through autophagy stimulation. Thus, β-elemene treatment facilitated autophagy-mediated apoptosis.
Autophagy is important for the toxic effects of β-elemene/PPP combination
IGF-1R inhibitor picropodophyllin (PPP) is effective in ESFT preclinical models and involved in phase I and II clinical trials for ESFTs (Wu et al., 2021). In this study, we treated the cells using a low concentration of PPP (0.2 μM) together with β-elemene (30 μg/mL), and the combination treatment resulted in a more significant reduction in cell viability than the single treatment with PPP or β-elemene only. 3-MA markedly retrieved the cell viability of PPP/β-elemene-treated cells (Fig. 4A–D). These results indicate that PPP/β-elemene-triggered autophagy ultimately results in autophagic cell death in ESFT cells. And a greater upregulation in the levels of LC3-II as well as cleaved-PARP was observed in Western blotting results, compared with those in the cells treated with PPP or β-elemene only (Fig. 4E). Therefore, autophagy should be important for the toxicity of β-elemene/PPP combination.
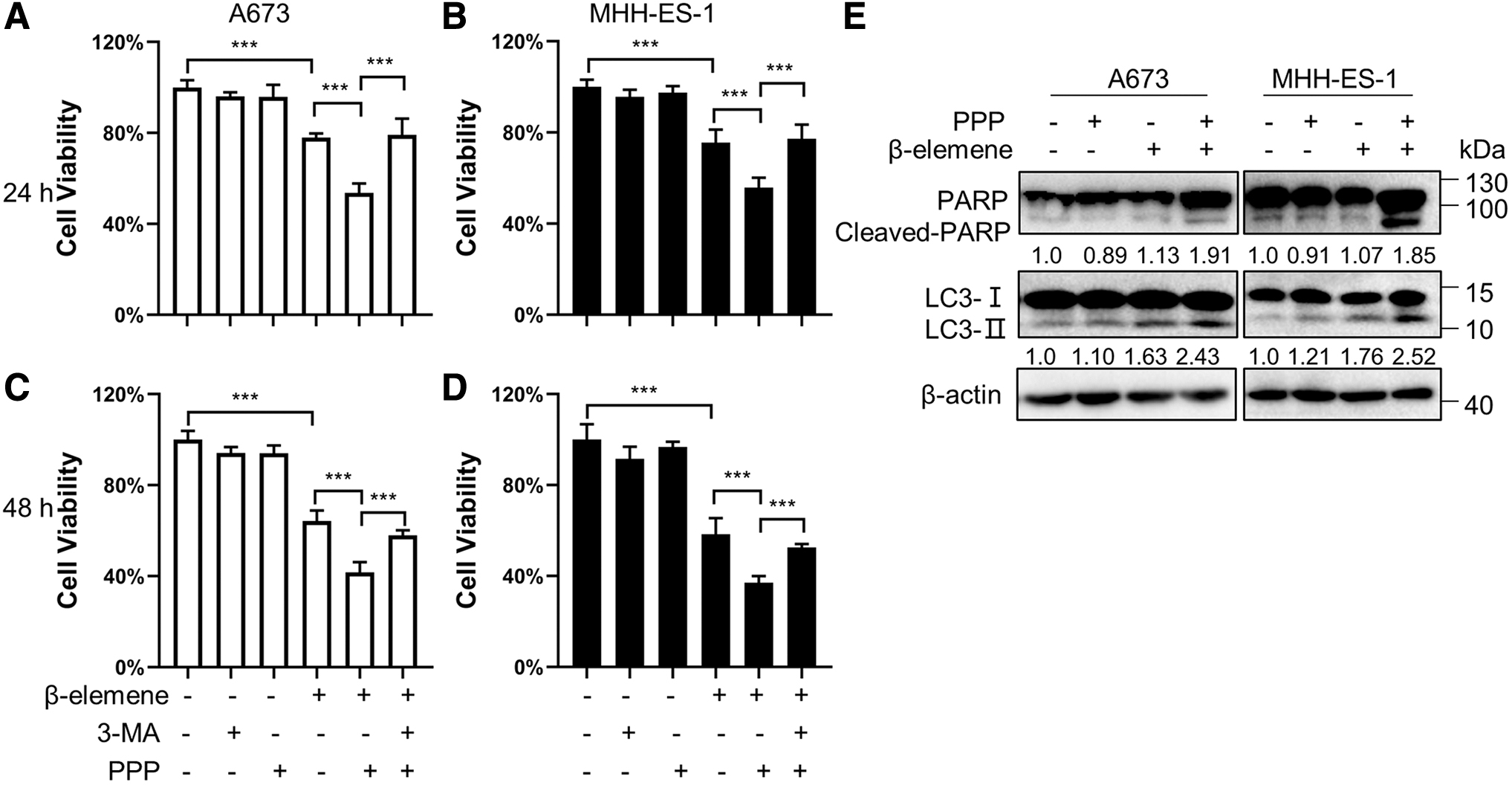
Effects of the combination treatment of β-elemene and PPP on A673 and MHH-ES-1 cells. Cells were exposed to the indicated treatments for 24 or 48 h. Cell viability analyses
Discussion
ESFTs are highly ranked in the pediatric tumors with unfavorable prognosis, lacking efficient therapies (Riggi et al., 2021). β-elemene showed toxicity to various tumor cells, including ESFT cells, whereas the role of autophagy in the effects of β-elemene treatment remains debatable. In this study, we investigated the relationship of autophagy and β-elemene-induced cell death, and found that β-elemene facilitated autophagic flux, which was necessary for the apoptosis caused by β-elemene treatment in ESFT cells (Figs. 1–3 and 5). In addition, combination of β-elemene and IGF1R inhibitor showed toxic effects in ESFT cells through autophagy promotion (Fig. 4).
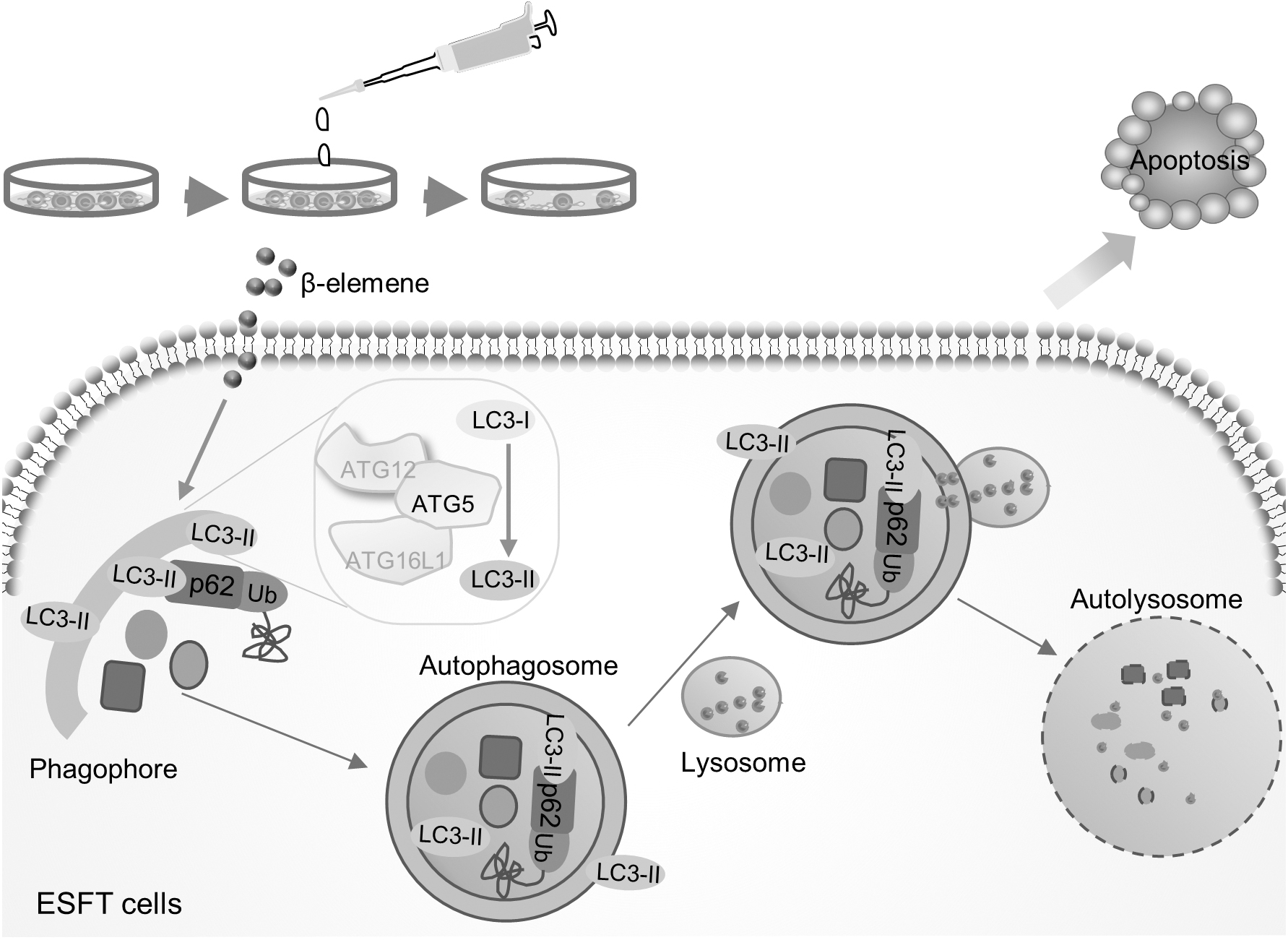
Schematic graph representing the working model of β-elemene in ESFT cells.
LC3-I is conjugated with phosphatidylethanolamine (PE) on the autophagosomal membrane to form LC3-II, which would be degraded by lysosomal proteases along with the autophagosomal cargo (Kabeya et al., 2000). Therefore, accumulation of LC3-II not only reflects the increasing number of autophagosomes caused by enhancement of autophagic activity, but also is caused by the obstruction of the fusion of autophagosomes and lysosomes or the inhibition of the degradation of autolysosomes. Herein we found that the level of LC3-II was significantly upregulated by β-elemene treatment in ESFT cells (Figs. 1 and 2).
CQ was next used to block the fusion of autophagosomes and lysosomes, and LC3-II level was increased more by β-elemene treatment (Fig. 2A). Simultaneously, β-elemene treatment downregulated the level of autophagy substrate p62, which was inhibited by combined with CQ (Fig. 2A). Meanwhile, we transfected the mRFP-EGFP-LC3 reporter into A673 cells, and β-elemene treatment was found visibly to be able to increase the number of autolysosomes (Fig. 2B, C). These data suggest that both autophagosome formation and autophagic flux were activated by β-elemene treatment.
Autophagy often promotes cell survival, and sometimes is required for cell death execution. Either autophagy activation or autophagy inhibition is involved in the effects of anticancer drugs (Kim et al., 2021). Our results showed that autophagy inhibitor 3-MA weakened β-elemene-induced apoptosis and restored cell viability. Furthermore, ATG5 knockdown that impaired autophagosome formation reversed the cell viability and the levels of LC3-II and p62 altered by β-elemene treatment (Fig. 3). Taken together, these data indicate that autophagy is necessary for the cell death induced by β-elemene treatment in ESFT cells, suggesting that β-elemene treatment triggered autophagy-mediated cell death.
Among the novel treatments of ESFTs, IGF1R blockade is considered effective according to the results of clinical trials (Anderson et al., 2016; Kurzrock et al., 2010; Olmos et al., 2010). However, drug resistance and disease recurrence remained to be unresolved problems. Our data have shown that the combination with low concentration of β-elemene (30 μg/mL) and IGF1R inhibitor PPP (0.2 μM) resulted in significant toxic effects in ESFT cells, which might provide a new way to prevent the drug resistance or disease recurrence of IGF1R blockade. In addition, the increased LC3-II level indicated the important role of autophagy in the combination treatment (Fig. 4), which indicated the possibility of combining autophagy activation with IGF1R blockade.
Conclusion
β-elemene treatment increases autophagy-mediated apoptosis in ESFT cells. Autophagy is important for the toxic effects of β-elemene/PPP combination. Our data provided a novel insight for the development of new ESFT treatments.
Footnotes
Acknowledgments
We thank all the Huang laboratory members for the kind support.
Authors' Contribution
Data curation, investigation, validation, and writing—original draft by T.Z. Investigation and validation by T.Z., C.G., S.J., R.Y., and H.T. Conceptualization, validation, and supervision by C.L. Conceptualization, funding acquisition, methodology, project administration, resources, supervision, and writing—review and editing by L.H.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Educational Department of Liaoning Province (LJKFZ20220250) to L Huang.