
Abstract
Oridonin (ORI), derived from Chinese herbs Rabdosia rubescens, has anti-inflammatory, proapoptotic, anticancer effects. Previous studies have found that ORI induces apoptosis in rheumatoid arthritis fibroblast synovial cells (RA-FLSs), but this mechanism is not clear. We will investigate the apoptosis mechanism of ORI on RA-FLSs. RA-FLSs were treated with various concentrations of ORI (0, 5, 10, 15, 20, 25, and 30 μM) for 24 h. CCK8, LDH, and hochest/PI assay determined the viability, cytotoxicity, and death of ORI on RA-FLSs. The endoplasmic reticulum probe was used to observe structural changes of endoplasmic reticulum in RA-FLSs. RNA expression was detected with RNA sequencing analysis and quantitative real-time PCR. The PERK/eIF2α/CHOP pathway protein of the endoplasmic reticulum was verified with Western Blot. Our results show that ORI induced the apoptosis of RA-FLSs from CCK8, LDH, and Hochest/PI. The endoplasmic reticulum distribution was altered in RA-FLSs after being treated with ORI. Bioinformatics analysis of RNA sequencing data found that 1453 genes were elevated. The PERK/eIF2α/CHOP pathway of the endoplasmic reticulum was regulated from the Gene ontology and KEGG analysis. The results of quantitative real-time PCR and Western blot analysis verified the regulation of PERK/eIF2α/CHOP pathway in RA-FLSs. Our data imply that the endoplasmic reticulum's PERK/eIF2α/CHOP signaling pathway is certainly implicated in the induction of RA-FLS apoptosis by ORI. This study has important implications for the pharmacological effects of ORI and the treatment of RA.
Introduction
Rheumatoid arthritis (RA) is a chronic, diverse inflammatory, and immunological disease. The global prevalence of RA is estimated to be 0.5–1% (Gravallese and Firestein, 2023). Joint swelling, cartilage degradation, and joint abnormalities are the most prevalent RA symptoms. The tumor-like growth of intra-articular fibroblast synovial cells (FLSs) is the key pathogenic characteristic of RA (Alivernini et al., 2022). The tumor-like growth of rheumatoid arthritis fibroblast synovial cells (RA-FLSs) relates to apoptotic resistance (Fava et al., 1994). The antiapoptotic mechanism of RA-FLSs is mediated through the Fas receptor, TNF receptor, P53, and mitochondrial pathways (Pope, 2002; Zhai et al., 2018a). Protein kinase R-like endoplasmic reticulum kinase (PERK), an endoplasmic reticulum stress-related kinase, has been associated to cell death (Bonsignore et al., 2023; Liu and Ju, 2023). However, it is uncertain whether RA-FLS antiapoptosis is linked to the endoplasmic reticulum stress-related kinase PERK.
Oridonin (ORI) is a naturally occurring terpenoid derived from the Chinese herb Rabdosia rubescens. ORI has been shown to have anti-inflammatory, antitumor, antimicrobial, and immunomodulatory properties (Li et al., 2021). It has been reported to be effective against colon, ovarian, and gastric cancers (He et al., 2017; Ma et al., 2016; Yang et al., 2017). Research found that ORI regulated the cell cycle, autophagy, and apoptosis to promote cell death (Liu et al., 2012). Our study found that ORI enhanced apoptosis in RA-FLSs by regulating autophagy (He et al., 2020), but the mechanism is unclear. Therefore we will show the mechanism of ORI that influences the endoplasmic reticulum stress pathway to trigger apoptosis in RA-FLSs to provide new ideas and strategies for the prevention and treatment of RA.
Materials and Methods
Cell culture
RA-FLSs were obtained from patients admitted to the Huazhong University of Science and Technology Union Shenzhen Hospital. The study was approved by the Institutional Research Ethics Committee of Huazhong University of Science and Technology Union Shenzhen Hospital (No. M202200122). RA-FLSs were cultured as described in detail previously (He et al., 2020). Tissue samples were cut into 4 × 4 × 2 mm fragments and maintained in a low-glucose DMEM culture medium containing 10% FBS, and 1% penicillin-streptomycin at 37°C. We gently added 1–2 mL complete DMEM to the bottom of the flask without agitation, incubated the flask at 37°C in a 5% carbon dioxide incubator with the daily medium change, and monitored the culture until the cells were confluent. For subculturing purposes, cells were detached using 0.05% trypsin–ethylenediaminetetraacetic acid treatment at 37°C.
Cell viability assay
CCK8 assay was used to determine the viability of RA-FLSs upon treatment with ORI, as described in detail previously. In brief, RA-FLSs were planted in the 96-well plates at the density of 2 × 104 cells per well and incubated overnight. Afterward, ORI with various concentrations (0, 5, 10, 15, 20, 25, and 30 μM; Aladdin, Shanghai, China) was administered for 24 h of incubation. Ten microliters of CCK8 (Beyotime, Shanghai, China) were added, and incubated for another 4 h at 37°C. Then the absorbance was measured at 450 nm on a microplate spectrophotometer. The cell viability was presented as the decrease over the control group: [ODsample−ODblank]/[ODcontrol−ODblank].
LDH assay
LDH assay was used to determine the cytotoxicity of ORI to RA-FLSs. RA-FLSs were planted in 96-well plates at the density of 2 × 104 cells per well and cultured overnight. Afterward, ORI with various concentrations (0, 5, 10, 15, 20, 25, and 30 μM) was administered for 24 h of incubations. Cell supernatant was collected and centrifuged at 700 g for 10 min at room temperature, 60 μL cell-free supernatant was aspirated from the cell supernatant and incubated with 30 μL LDH (Beyotime) substrate solution for 30 min, and then the absorbance at 490 nm was recorded on a microplate spectrophotometer.
Hochest/PI assay
Hochest/PI assay was used to determine the survival and death of ORI to RA-FLSs. RA-FLSs were seeded in 96-well tissue culture plates at the density of 2 × 104 cells per well and cultured overnight. According to CCK8 results, ORI various concentrations 15, 20, 25 μM was added for 24 h, then cell were stained with DAPI (Beyotime) and PI (Beyotime) mixed solution for 10 min. The nucleus was monitored by a fluorescence microscope excited by blue light and ultraviolet light.
Endoplasmic reticulum stress assay
Endoplasmic reticulum probe was used to determine the effect of ORI on RA-FLSs, RA-FLSs were seeded in 48-well tissue culture plates at the density of 4 × 104 cells per well and cultured overnight. After the ORI solution was added at final concentrations of 15 and 20 μM for 24 h, the endoplasmic reticulum probe (Beyotime) was added for 30 min after the PBS was washed twice, and then nuclei were stained with DAPI solution for 10 min. The intracellular localization of RA-FLSs was detected by fluorescence microscope.
RNA sequencing and analysis
RNA samples from the control group and 20 μM ORI group were submitted to Beijing Genomics Institution Genome for RNA sequencing and analysis. RA-FLSs were seeded in six-well tissue culture plates at the density of 5 × 105 cells per well and cultured overnight. After ORI solution was added at a final concentration of 20 μM for 12 h, a total of six samples were available, including three samples of ORI groups and three samples of control groups. Total RNA from cultured cells was extracted with Trizol RNA isolation reagents (YESEN, Shanghai, China). RNA samples in triplicates for each group were submitted to Beijing Genomics Institution (Wuhai, China) for RNA sequencing and analysis.
The results were filtered and the cutoff point was set to twice the difference. Differential expression analysis of two groups was performed using the edgeR, DESeq software. Cluster analysis and the heat map use the multi-experiment viewer software. Differential gene ontology (GO) enrichment analysis uses Goatools (
Quantitative real-time PCR analysis
The candidate genes were validated using qPCR. RNA samples were prepared the same way as preparing RNA-seq samples, as described above. The concentration and purity of total RNA were measured by NanoDrop 2000 Spectrophotometer (Thermo Fisher Scientific, USA). Two micrograms RNA were converted to cDNA by a Rever-tAidTM First Strand cDNA Synthesis Kit with gDNA Eraser and amplified using PowerUPTMSYBR Green Master mix kit (all from Thermo Fisher Scientific) in an ABI 7500/7500Fas Detection System (ABI, USA). The thermal profile for qPCR was 60 s of preincubation at 93°C for one cycle, followed by 40 cycles of 93°C for 30 s, 55°C for 30 s, and 70°C for 30 s. The full primer list is shown in Table 1.
The Primers Used for Real-Time Polymerase Chain Reaction Analysis
Western blot assays
The protein expression assay was used to explore the pathway effect of ORI on RA-FLSs. RA-FLSs were seeded in six-well tissue culture plates at the density of 5 × 105 cells per well and cultured overnight. After ORI solution was added at final concentrations of 15, 20, and 25 μM for 24 h, the cells were washed twice with cold PBS and lysed once for 30 min in the cell lysis buffer (Beyotime). Pyrolysis liquid was centrifuged at 13,000 g for 15 min, the supernatant was collected, and the protein produced from each sample was quantified using the Pierce Bicinchoninic acid Protein Assay Kit.
The protein was separated using 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis and electro-imprinted to polyvinylidene fluoride membrane. Following this, the transferred membranes were incubated with blocking in phosphate buffered saline with Tween 20 containing 5% skimmed milk powder, and then the membrane was submerged under the first antibody p-PERK (1:5000; Catalog No. 102853; Invitrogen), p-eIF2α (1:5000; Catalog No. 3398; CST), ATF4 (1:5000; Catalog No. 11815; CST), CHOP (1:5000; Catalog No. 2895; CST), and Bax (1:5000; Catalog No. 2772; CST) overnight at 4°C, followed by the peroxidase coupled second antibody anti-GAPDH (1:5000; Catalog No. 3700; CST) for 2 h at 37°C. The immunoreactive bands were visualized using enhanced chemiluminescence reagents (Beyotime) and imaged with X-film in a dark room.
Data analysis
All experiments were performed with at least three replicates per group. Statistical analyses were performed using GraphPad Prism 9.0. Experiments were presented as the mean ± standard deviation. Significance was determined using Student's t-test, one-way analysis of variance, as appropriate. A value of p < 0.05 was considered statistically significant.
Results
Effect of ORI on viability and cytotoxicity of RA-FLSs
To evaluate the effects of ORI on RA-FLSs, we determined the cell viability using the CCK8 assay. The CCK8 experiment indicated that ORI decreased RA-FLSs viability (Fig. 1A). The inhibitory half maximal inhibitory concentration of ORI on RA-FLSs was 20.21 μM after 24 h. As the report is consistent with our earlier study (He et al., 2020), this demonstrates that ORI has strong antiproliferation action on RA-FLSs. To investigate the cytotoxicity of ORI on RA-FLSs, we assessed the level of LDH release. LDH assay revealed that when ORI (20, 25, 30 M) treated RA-FLSs for 24 h, the level of LDH release in the supernatant increased significantly (Fig. 1B). Therefore, we chose ORI concentrations of 15, 20, and 25 μM for the following study.
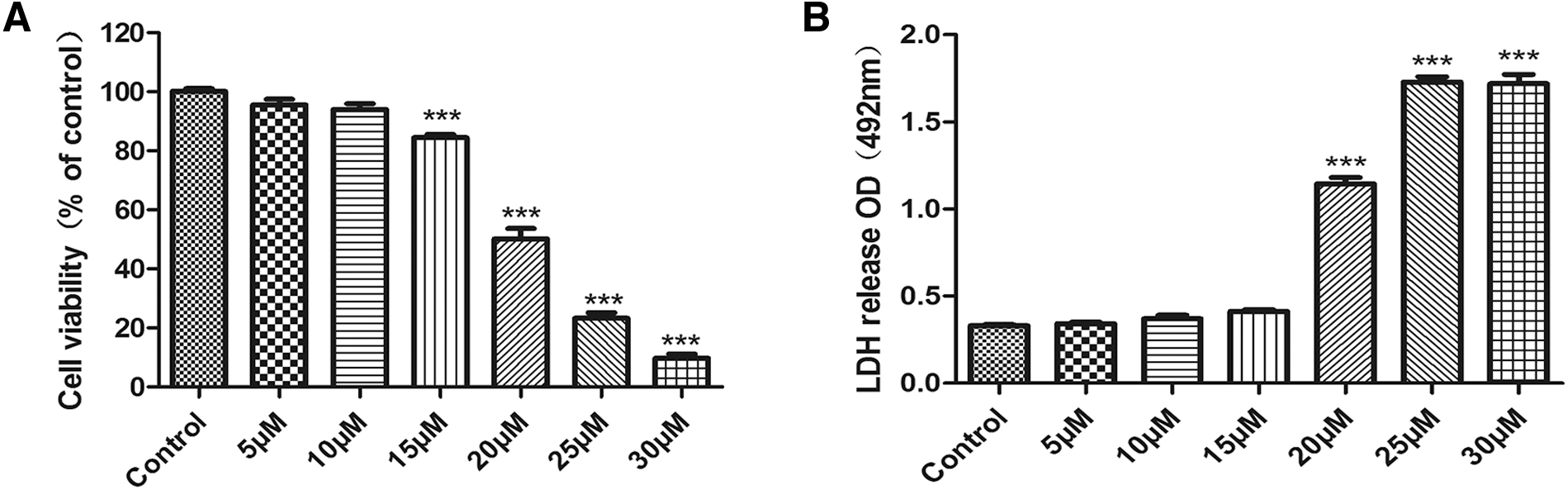
Effect of ORI on viability and cytotoxicity for RA-FLSs.
Effect of ORI on death in RA-FLSs
To investigate whether ORI induced death on RA-FLSs by Hochest/PI assay. Hoechst stain could easily penetrate the cell membrane and finally bound to DNA, the blue DAPI stain shows the nuclei. The death was measured using a PI fluorescent red probe. The results showed that ORI (15, 20, and 25 μM) treated RA-FLSs, the death percentage was significantly increased (Fig. 2A, B).
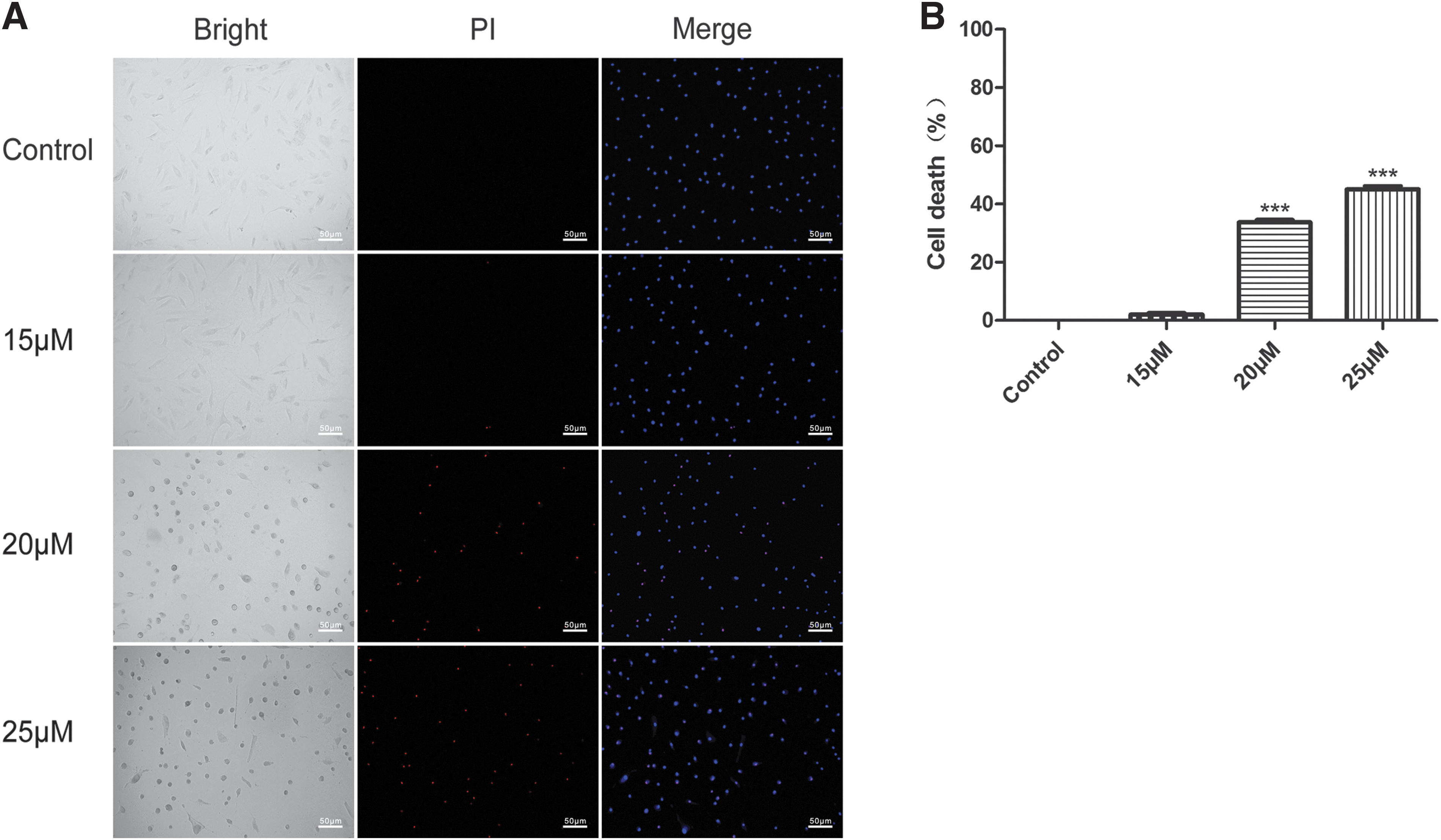
Effect of ORI on cell death for RA-FLSs.
Effect of ORI on the endoplasmic reticulum assay in the RA-FLSs
The endoplasmic reticulum is directly associated with cell death. We investigated the endoplasmic reticulum after ORI intervention in RA-FLSs. The results showed that following ORI intervention in the endoplasmic reticulum, the endoplasmic reticulum distribution changed dramatically and the endoplasmic reticulum structure condensed from the long spindle to round, distributed around the nucleus (Fig. 3).
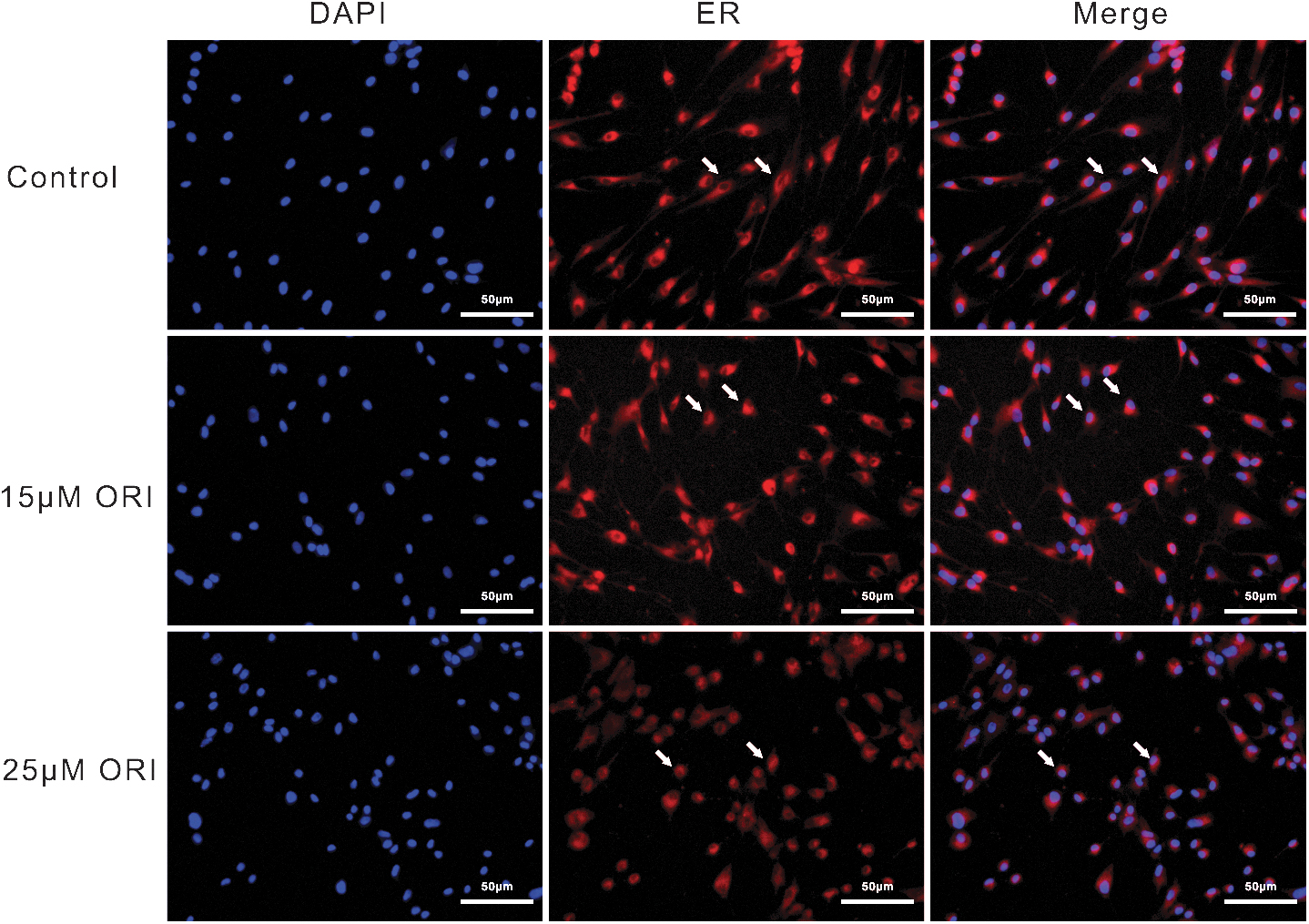
Effect of ORI on endoplasmic reticulum for RA-FLSs. The effect of ORI (15 and 25 μM) on endoplasmic reticulum of RA-FLSs for 24 h. The blue fluorescence of DAPI staining viability nuclei, The ER-Tracker Red fluorescence staining endoplasmic reticulum.
RNA sequencing and analysis
The endoplasmic reticulum distribution significantly altered after ORI treatment of RA-FLSs. Therefore, we explored the molecular mechanisms of the endoplasmic reticulum stress pathway using bioinformatics tools. Bioinformatics analyses of RNA sequencing data revealed 2493 genes that were differentially expressed (fold change ≥2, false discovery rate <0.05). One thousand four hundred fifty-three genes were upregulated, and 1042 genes were downregulated in ORI groups.
The distribution of differentially expressed genes between control and ORI groups is shown in the volcano plot. The green dots represented downregulated genes, while the purple ones represented upregulated genes in the volcano plot (Fig. 4A). GO analysis revealed that the 1453 differentially expressed genes were significantly enriched in 20 GO terms related to the biological process, of which the endoplasmic reticulum unfolded protein response was considerably enriched in GO biological process (Fig. 4B). Gene set enrichment analysis shows the high expression of endoplasmic reticulum-to-cytosol transport (Fig. 4C). In the network analysis of “protein processing in endoplasmic reticulum” protein expression, the protein processing in endoplasmic reticulum was significantly upregulated by ORI; the predicted protein of eIF2α, ATF4, and CHOP was upregulated expression in the PERK pathway of endoplasmic reticulum (Fig. 4D).
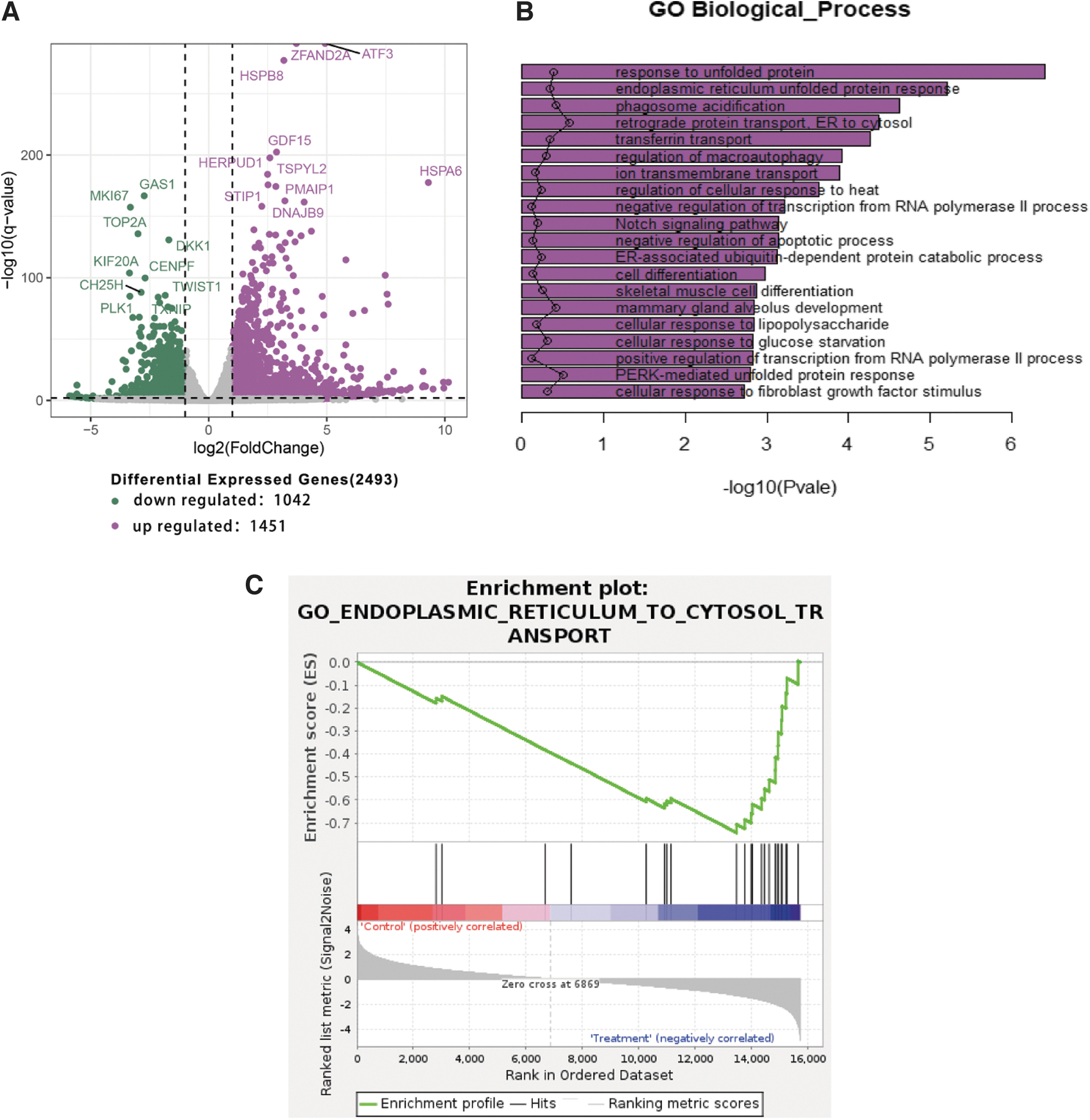
Genome-wide analysis of ORI on endoplasmic reticulum in the RA-FLSs.

Quantitative real-time PCR analysis
Real-time PCR was performed for gene quantitative analysis to confirm the changes in RNA sequencing results. Four genes relevant to the endoplasmic reticulum signaling pathway were selected (PERK, eIF2α, ATF4, and CHOP). As shown in Figure 5, similarly, the levels of eIF2α, ATF4, and CHOP mRNA were significantly increased in RA-FLSs. The PERK level also increased in RT-qPCR, but not in RNA-seq data. This could be due to different sensitivities of these two methods.
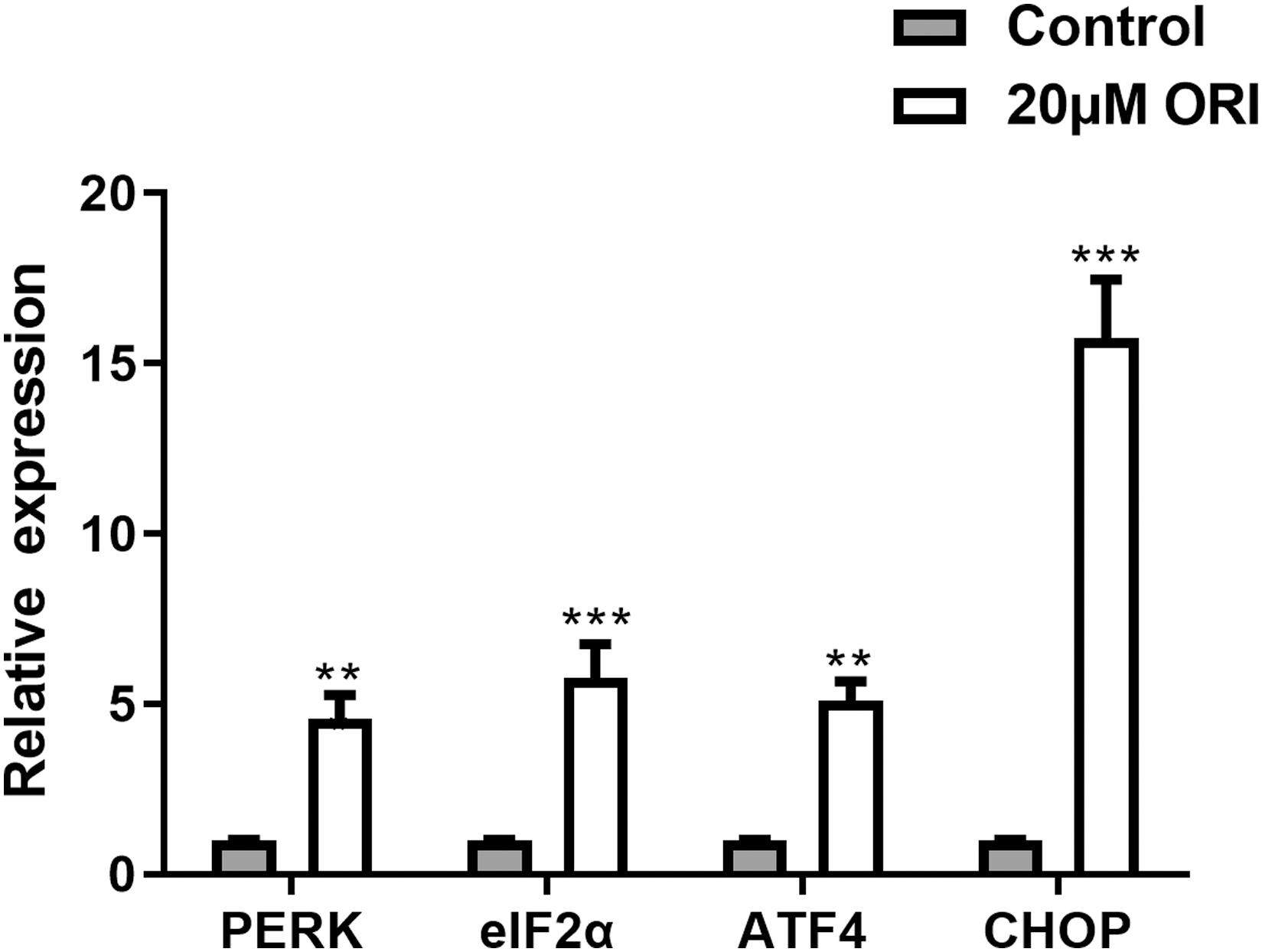
The gene mRNA expression of PERK, eIF2α, ATF4, and CHOP in RA-FLSs treated with 20 μM ORI or control for 12 h. Significance of differences between means: **p < 0.01, ***p < 0.001 versus control group.
Western blot analysis
To further verify whether the PERK/eIF2α/CHOP of endoplasmic reticulum stress pathway was activated, the expression of five proteins (p-PERK, p-eIF2α, ATF4, CHOP, and Bax) was examined using a Western blot. As shown in Figure 6, compared with the control group, the expression levels of p-PER, p-eIF2α, ATF4, CHOP, and Bax protein were significantly increased when treated with 25 μM ORI for 24 h. The results were consistent with the RNA sequencing and real-time PCR.

The protein expression of p-PERK, p-eIF2α, ATF4, CHOP, and Bax in RA-FLSs treated with ORI (15, 20, and 25 μM) for 24 h. Data were presented as mean ± SD (n = 3). Significance of differences between means: *p < 0.05, **p < 0.01, ***p < 0.001 versus control group.
Discussion
The pathogenic mechanism of rheumatoid arthritis is the tumor-like development of fibroblast synovial cells (Alivernini et al., 2022; Zhai et al., 2019). Fibroblastic synovial cell tumor-like growth results in the production of matrix metalloproteinases and the release of inflammatory agents such as IL-1β, IL-6, and TNF-α, as well as pannus growth and articular cartilage and bone degradation (McInnes and Schett, 2017; Zhai et al., 2019).
An association has been discovered between tumor-like growth of RA-FLSs and apoptotic resistance (Fava et al., 1994). Drug-induced apoptosis of synovial cells is a potential therapy strategy for RA. Our findings show that ORI has a substantial antiproliferation effect against RA-FLSs in our study. We also discovered an increase in the apoptotic protein Bax. This result is consistent with earlier research (He et al., 2020; Shang et al., 2016). Because apoptosis increases cell membrane permeability and cell death (Kroemer et al., 1998), we discovered that ORI increased LDH and cell death using the LDH release and Hochest/PI tests. Therefore, we hypothesized that ORI activates the apoptotic pathway in RA-FLSs.
ORI is the main active ingredient isolated from the Chinese herb R. rubescens. ORI has anti-inflammatory, proapoptotic, and anticancer effects (Li et al., 2021). ORI triggered RA-FLSs apoptosis in our study. From the RNA sequencing results, we discovered that the endoplasmic reticulum stress signaling system was strongly expressed, with enhanced RNA gene expression of the PERK/eIF2α/CHOP pathway. According to research, the endoplasmic reticulum stress-related kinase PERK is linked to apoptosis (Liu and Ju, 2023). Endoplasmic reticulum stress is related to the inflammation of RA (Tang et al., 2022).
The endoplasmic reticulum stress pathway refers to the activation of unfolded protein response, endoplasmic reticulum overload response, and caspase-12-mediated apoptosis pathway in response to misfolded and unfolded protein aggregation in the endoplasmic reticulum lumen and disturbed calcium ion homeostasis (Ji et al., 2014). GRP78, a glycoregulatory protein found in the endoplasmic reticulum lumen, binds to three transmembrane proteins on the ER surface: inositol requiring enzyme 1 (IRE1), activating transcription factor 6 (ATF6), and PERK, rendering signal transduction factors inactive (Wang et al., 2018).
When unfolded or misfolded substrates accumulate in the ER lumen, PERK, IRE1, and ATF6 are released from the GRP78/BIP binding domain, phosphorylated IRE and PERK induce protein CHOP synthesis. The CHOP is the most important hallmark protein in endoplasmic reticulum stress induced apoptosis and involved in the regulation of apoptosis in response to numerous cellular stressors (Maurel et al., 2014). Overexpression of CHOP increases the expression of Bax, a proapoptotic Bcl-2 family component (Yu et al., 2014). Clinical medicines have been shown to cause rheumatoid arthritis cell death through persistent, excessive endoplasmic reticulum stress-induced CHOP expression or activation of caspase-12-dependent pathways. Heme oil induced apoptosis in rheumatoid arthritis fibroblastic synovial cell line MH7A cells by increasing CHOP expression (Jeong et al., 2014). Resveratrol elevated the expression of Caspase-12 and CHOP and caused the formation of RA-FLSs apoptosis (Lu et al., 2019).
In addition, some studies report ORI induced apoptosis in tumor cells by activating the protein of endoplasmic reticulum stress pathway. ORI increases phosphorylated PERK and CHOP expression in HepG2 cells and promotes apoptosis in hepatoblastoma HuH-6 cells by activating the endoplasmic reticulum stress system through the IRE1/ASK1/JNK pathway (Cai et al., 2013; Wang et al., 2011). Our results showed that the PERK, eIF2α, and CHOP mRNA and protein expression of PERK/eIF2α/CHOP pathway were increased. In conjunction with our earlier findings, we hypothesize that ORI promotes apoptosis in RA-FLSs by activating the PERK/eIF2/CHOP of endoplasmic reticulum stress signaling pathway.
Although our work suggested that ORI might be able to activate the PERK/eIF2α/CHOP endoplasmic reticulum stress signaling pathway to trigger apoptosis in RA-FLSs, this experiment is still not quite sufficient. First, we solely looked at phenotypic changes rather than designing intervention trials with the endoplasmic reticulum stress signaling pathway. Second, Shang reported that ORI may induce IL-1β-exposed RA-FLSs by reducing the phosphorylation of ERK1/2 and JNK expression (Shang et al., 2016); we did not evaluate the ERK1/2 and JNK pathways in our study.
Oxidative stress is related to the inflammation of RA (Tang et al., 2022). The Nrf2-HO-1-ROS pathway could improve RA symptoms by inhibiting oxidative stress (Zhai et al., 2018b). Interestingly, PERK can activate the Nrf2-HO-1 pathway (Dong et al., 2023); possibly, ORI can suppress inflammation in RA-FLSs by activating the PERK/Nrf2-HO-1 pathway. Thus, we need to further investigate whether endoplasmic reticulum stress regulation is related to the ERK1/2, JNK, and Nrf2-HO-1pathways in the treatment of ORI on RA.
Conclusion
In summary, ORI decreases RA-FLSs activity, while also increasing LDH release and inducing RA-FLSs death. ORI affected the endoplasmic reticulum structure of RA-FLSs. RNA sequencing data, quantitative real-time PCR, and Western blot tests revealed that the endoplasmic reticulum stress pathways PERK, eIF2α, and CHOP were considerably altered. These findings imply that the endoplasmic reticulum's PERK/eIF2α/CHOP signaling pathway is certainly implicated in the induction of RA-FLSs apoptosis by ORI (Fig. 7). This study has important implications for the pharmacological effects of ORI and the treatment of RA.

The PERK/eIF2α/CHOP signaling pathway of endoplasmic reticulum in the apoptosis of RA-FLSs by ORI.
Footnotes
Authors' Contributions
Participated in research and study design: S.H., N.T., and J.Z. Conducted experiments: S.H. Performed data analysis: C.H.. Wrote or contributed to the writing of the article: S.H.
Data Availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was funded by Shenzhen Nanshan District Scientific Research Project (Grant No. 2020088), Huazhong University of Science and Technology Union Shenzhen Hospital research project (Grant No. 2020006), and National Natural Science Foundation of China (Grant No. 82174290).