
Abstract
Cornus iridoid glycosides (CIGs), including loganin and morroniside, are the main active components of Cornus officinalis. As one of the key enzymes in the biosynthesis of CIGs, geranyl pyrophosphate synthase (GPPS) catalyzes the formation of geranyl pyrophosphate, which is the direct precursor of CIGs. In this study, the C. officinalis geranyl pyrophosphate synthase (CoGPPS) sequence was cloned from C. officinalis and analyzed. The cDNA sequence of the CoGPPS gene was 915 bp (GenBank No. OR725699). Phylogenetic analysis showed that CoGPPS was closely related to the GPPS sequence of Actinidia chinensis and Camellia sinensis, but relatively distantly related to Paeonia lactiflora and Tripterygium wilfordii. Results from the quantitative real-time PCR showed the spatiotemporal expression pattern of CoGPPS; that is, CoGPPS was specifically expressed in the fruits. Subcellular localization assay proved that CoGPPS was specifically found in chloroplasts. Loganin and morroniside contents in the tissues were detected by high-performance liquid chromatography, and both compounds were found to be at higher levels in the fruits than in leaves. Thus, this study laid the foundation for further studies on the synthetic pathway of CIGs.
Introduction
C
C. officinalis is mainly distributed in Henan, Shaanxi, and Gansu Provinces and other regions in China (Bai et al., 2014). The yield and quality of C. officinalis are subject to regional environmental and processing conditions (Jeung et al., 2019; Yu et al., 2010). However, the contents of active components of C. officinalis cannot meet the increasing demand for medical application (Yang et al., 2019). Extensive exploitation of C. officinalis for its commercially and ecologically valued metabolites needs genetic engineering strategies to increase the levels of active substances in this species (Alagna et al., 2016; Zhou et al., 2016).
CIGs, which are monoterpenes, are some of the most important secondary metabolites in C. officinalis. Terpenoids are derived from two 5-carbon (C5) units named dimethylallyl diphosphate (DMAPP) and isopentenyl diphosphate (IPP), which is an isomer of DMAPP (Christianson, 2007). Plants have two pathways to synthesize the C5 units, namely the mevalonate pathway located in the cytoplasm and the 2-C-methyl-D-erythritol 4-phosphate pathway (MEP) in plastids (Liao et al., 2016; Vranová et al., 2013).
Studies have demonstrated that most iridoid glycosides are formed by condensation of the C5 structural units of the MEP pathway (Oudin et al., 2007). The synthesis of iridoid glycosides can be divided into three steps. The first step is synthesis of DMAPP and IPP. The second step is synthesis of the iridoid skeleton. The final step is synthesis of iridoid glycosides and various modification reactions (Singh et al., 2021).
Trans-pentenyl transferase catalyzes DMAPP and IPP to synthesize the carbon skeleton of iridoids. This enzyme can be divided into three types according to the carbon chain lengths of its products, namely geranyl pyrophosphate synthase (GPPS), farnesyl diphosphate synthase (FPPS), and geranylgeranyl diphosphate synthase (GGPPS) (Lackus et al., 2019; Maheshwari et al., 2020).
GPPS catalyzes the head–tail condensation of DMAPP and IPP into C10-geranyl diphosphate (GPP), which is the direct precursor of iridoid glycosides (Tholl et al., 2004). FPPS catalyzes GPP and IPP to synthesize C15-farnesyl pyrophosphate (FPP), and GGPPS catalyzes FPP or GPP and IPP to synthesize C20-geranylgeranyl pyrophosphate (GGPP) (Alcaíno et al., 2014).
GPP is an important node in the biosynthesis of terpenoids, from which monoterpenes, diterpenes, triterpenes, and various terpenoids are synthesized in the plastids (Coman et al., 2014; Rubat et al., 2017). Given that GPPS is a key enzyme in the iridoid pathway, it plays an important role in the growth and development of plants.
To date, GPPS genes have been isolated from more than 20 plants, such as Mentha spicata, Antirrhinum majus, Mentha piperita, Clarkia breweri, and Camptotheca acuminata (Chang et al., 2010; Reddy et al., 2017; Yang et al., 2017). The plant GPPS genes generally have two structural forms, namely homodimers and heterodimers. The heterodimers comprise large subunits (LSUs) and small subunits (SSUs) (Schmidt and Gershenzon, 2008).
Studies have shown that LSUs and SSUs are similar to GGPPS. The similarity between LSUs and GGPPS is as high as 70%, and GGPPS can replace LSUs and combine with SSUs to form the heterodimer GPPS, which can catalyze the synthesis of iridoids (Tholl et al., 2004). Comparison of amino acid sequences revealed that the LSU contains aspartate-rich regions (DDxxD and DDxxxxD), which are essential for binding substrates and catalyzing reactions, while the SSU contains two CxxxC conserved motifs, which are used for the interaction between two subunits (Kamran et al., 2020).
The SSU has been reported to be responsible for regulating the activity of LSU, and LSU is responsible for catalyzing the synthesis of GPP (Reddy et al., 2017). Studies have shown that the combination of LSU and SSU can catalyze the production of GPP and GGPP in vitro, but only GPP in vitro. The combination of GGPPS and SSU can form the heterodimer GPPS that synthesizes GPP.
The results indicated that the heterodimer GPPS gene can catalyze the production of GPP under the regulatory activity of the SSU, which determines the synthesis of metabolites such as monoterpenes (Rai et al., 2013). Studies have indicated that the LSU has catalytic activity alone, but exceptions are found in the case of Humulus lupulus (Wang and Dixon, 2009).
In this study, the CoGPPS gene was cloned from the leaf and fruit tissues of C. officinalis. We have analyzed the CoGPPS gene expression in C. officinalis. We performed subcellular localization, prokaryotic expression analysis, and high-performance liquid chromatography (HPLC) analysis.
These studies will lay a foundation for further analysis of the function of the CoGPPS gene in C. officinalis.
Materials and Methods
Plant material and sampling
The leaves and fruits were taken from the Sui Tang Dynasty City Ruins Botanical Garden (Luoyang, China) in the middle of May, June, July, August, September, and October 2020 (Fig. 1). All tissues were frozen in liquid nitrogen and stored at −80°C.

The fruits of Cornus officinalis at different months.
Wild-type tobacco (Nicotiana benthamiana) was used in this study. The seeds were soaked in a Petri dish with water. Two-day cold-treated seeds were sown on the surface of the soil (nutrient soil:vermiculite = 1:2) to germinate. The plants were placed in a 14-h light–10-h dark condition at 23°C ± 2°C. The leaves from 6-week-old plants were used to perform the subcellular localization study.
RNA extraction and cDNA synthesis
The total RNA from the C. officinalis leaves was isolated using the TaKaRa MiniBEST Plant RNA Extraction Kit (TaKaRa, Dalian, China) according to the manufacturer's protocol. The integrity of total RNA was detected through 1.5% agarose gel electrophoresis, and purity and concentration of RNA were tested using the NanoDrop™ 2000 ultra-micro spectrophotometer (NanoDrop Technologies). Single-strand cDNA was synthesized using the TRUEscript First Strand cDNA Synthesis Kit (Aidlab, Beijing, China).
CoGPPS gene cloning and sequencing
Gene-specific primers, namely CoGPPS-F and CoGPPS-R (Table 1), were designed based on the c70900_g1 sequence of CoGPPS from the transcriptome database of C. officinalis (Hou et al., 2018). The cDNA of CoGPPS was amplified using the first strand of cDNA as a template. The conditions for amplifying the CoGPPS gene were as follows: 94°C for 5 min; followed by 35 cycles at 95°C for 30 s, 55°C for 30 s, and extension at 72°C for 60 s; with a final extension at 72°C for 5 min. The PCR-amplified products were purified.
Designed Primer Sequences
The restriction site is underlined.
CoGPPS, Cornus officinalis geranyl pyrophosphate synthase.
A tail was added to the purified blunt-end PCR products and ligated into the pTOPO-T vector (Aidlab). Thus, the ligands were transformed into competent cell Escherichia coli DH5α and sequenced by Sangon Biotech (Shanghai).
Bioinformatic analysis
The open reading frame (ORF) of CoGPPS was predicted using ORFfinder from the National Center for Biotechnology Information (NCBI). The molecular weight and hydrophobicity of CoGPPS were analyzed using the ExPASy tool (Séverine et al., 2021). Subcellular localization and the conserved domain of CoGPPS were determined by BaCelLo (Andrea et al., 2006) and CD-Search, respectively.
A homologous search for the amino acid sequences of CoGPPS was conducted using an online BLASTp tool. The phylogenetic tree of CoGPPS was constructed using MEGA, version 7.0 (Sudhir et al., 2016), through the neighbor-joining method.
Quantitative real-time PCR analysis
The transcript level of the CoGPPS gene was detected by quantitative real-time PCR (qRT-PCR). RNA was extracted from the leaves and fruits of C. officinalis by using the TaKaRa MiniBEST Plant RNA Extraction Kit (TaKaRa) and reverse transcribed using the TRUEscript First Strand cDNA Synthesis Kit (Aidlab).
The primers (qCoGPPS-F and qCoGPPS-R) were designed using Integrated DNA Technologies (IDT), and the reference primers were qCoGAPDH-F and qCoGAPDH-R (Table 1). The Roche LightCycler®96 System and SYBR Green Master Mix Kit (Yeasen, Shanghai, China) were used for qPCR analysis. Each reaction was repeated three times, and values were calculated using the 2−ΔΔCt method (Livak and Schmittgen, 2001).
Subcellular localization
For protein localization, we constructed the CoGPPS-GFP fusion protein with the Kpn Ι and Spe Ι cloning sites of pCAMBIA1300-GFP. Gene-specific primers (Table 1) were used to amplify the ORF sequence of CoGPPS without a stop codon. The amplification product was fused with the C-terminal of GFP to generate pCAMBIA1300-CoGPPS-GFP. The resulting recombinant of CoGPPS-GFP was sequenced to verify the correct protein coding.
Then, pCAMBIA1300-CoGPPS-GFP was transferred into Agrobacterium tumefaciens (GV3101) competent strains. Transformed recombinant Agrobacterium cells were propagated in an Luria-Bertani liquid medium until the OD600 value reached 0.6, and the bacterial cells were collected. The suspension solution was then injected into the lower surface of 6-week-old tobacco leaves using a 1-mL sterile syringe.
After 36–48 h of infiltration, the fluorescence of the GFP was observed at 488 nm using a laser confocal microscope (FV3000; Olympus, Japan).
HPLC analysis
To analyze the loganin and morroniside contents of the different samples of C. officinalis, 0.1 g of ground powder from the frozen leaves and fruits was soaked in 10 mL of 80% (v/v) methanol, followed by ultrasonic extraction for 40 min at 60°C. All samples were filtered through a 0.22-μm filter membrane and then analyzed using HPLC.
Sample separation was performed on the Agilent 1260 Infinity HPLC System using a C18 (250 × 4.6 mm, 5 μm) reverse-phase column with MeOH (solvent A) and ultrapure water (solvent B) mobile phases, A:B ratios of 23:77, and a flow rate of 1.0 mL/min. The sample components were determined by a UV detector at a wavelength of 240 nm. A quantitative analysis was performed using the standards for loganin and morroniside.
Results
Characterization and analysis of the CoGPPS gene
From the cDNA library of C. officinalis, CoGPPS was cloned and found to have a full-length ORF. The CoGPPS gene included a 915-bp ORF and encoded a 304-amino acid protein. The molecular weight and theoretical isoelectric point of the CoGPPS predicted protein were 33.2 kDa and 6.2, respectively. The predicted protein belonged to the isoprenoid synthase superfamily, which uses IPP and DMAPP as substrates, and it is the key enzyme in the MEP pathway.
Phylogenetic analysis of CoGPPS
The BLASTp search revealed that the CoGPPS amino acid sequence shares 60–70% identity with the GPPS sequences of Castanea mollissima, Camellia sinensis, Paeonia lactiflora, Carpinus fangiana, Actinidia chinensis, Coffea eugenioides, and Tripterygium wilfordii. A phylogenetic tree was built to analyze the evolutionary relationship. CoGPPS and GPPS from A. chinensis and C. sinensis were clustered into the same branch on the tree (Fig. 2).
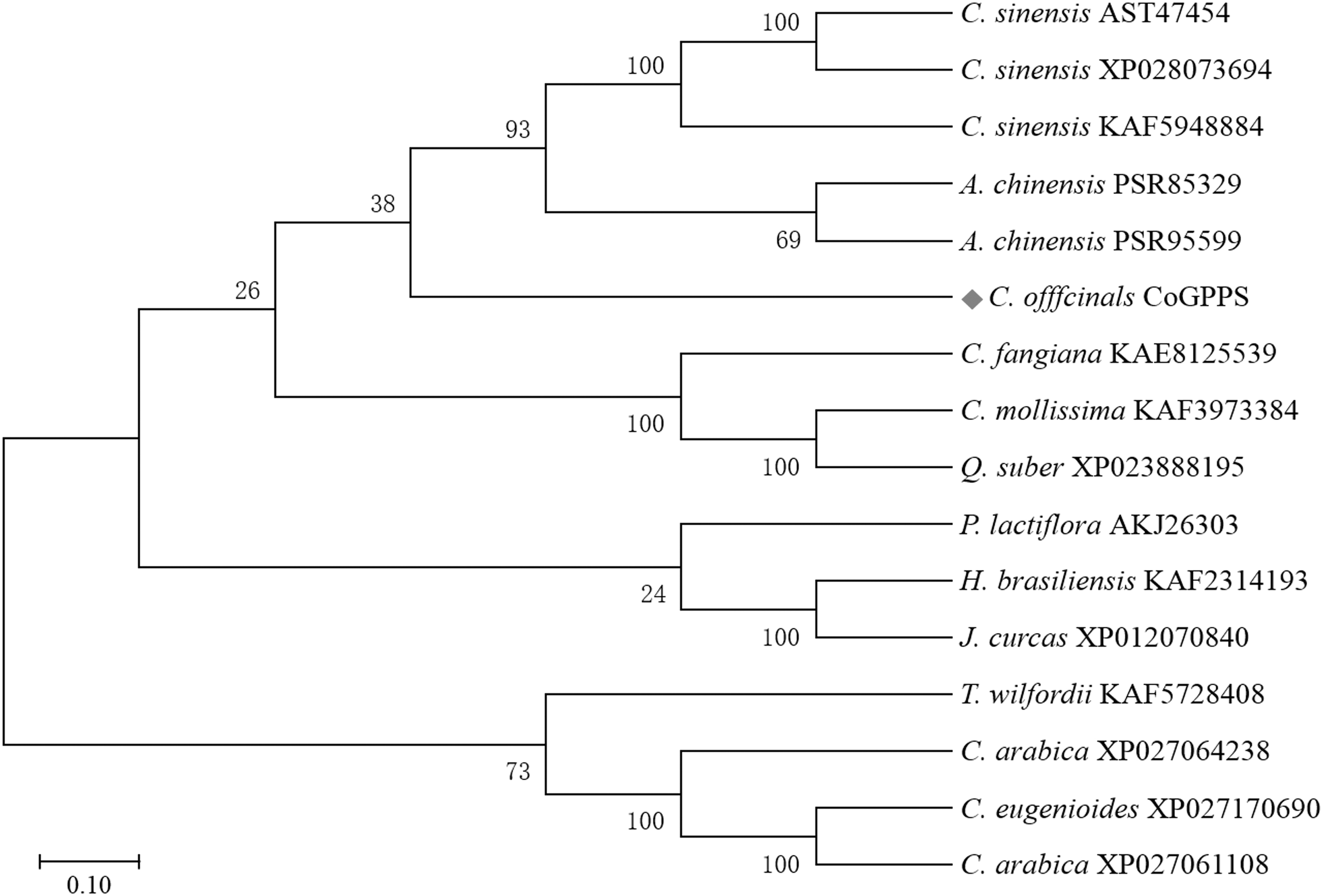
The phylogenetic tree of CoGPPS sequences of C. officinalis and its closely related species constructed with the GPPS sequences using the neighbor-joining method. CoGPPS, C. officinalis geranyl pyrophosphate synthase.
This result displayed that the CoGPPS was closely related to the GPPS sequences of A. chinensis and C. sinensis, but was relatively distantly related to those of P. lactiflora and T. wilfordii.
Spatiotemporal expression pattern of the CoGPPS gene
The expression levels of the CoGPPS gene in different tissues at various developmental stages were measured using qRT-PCR. The transcript levels of all samples were compared with the internal control of the relative transcript level of CoGPPS in the leaves in May. In detail, the CoGPPS gene was highly expressed in the fruits, but showed low expression in the leaves (Fig. 3).
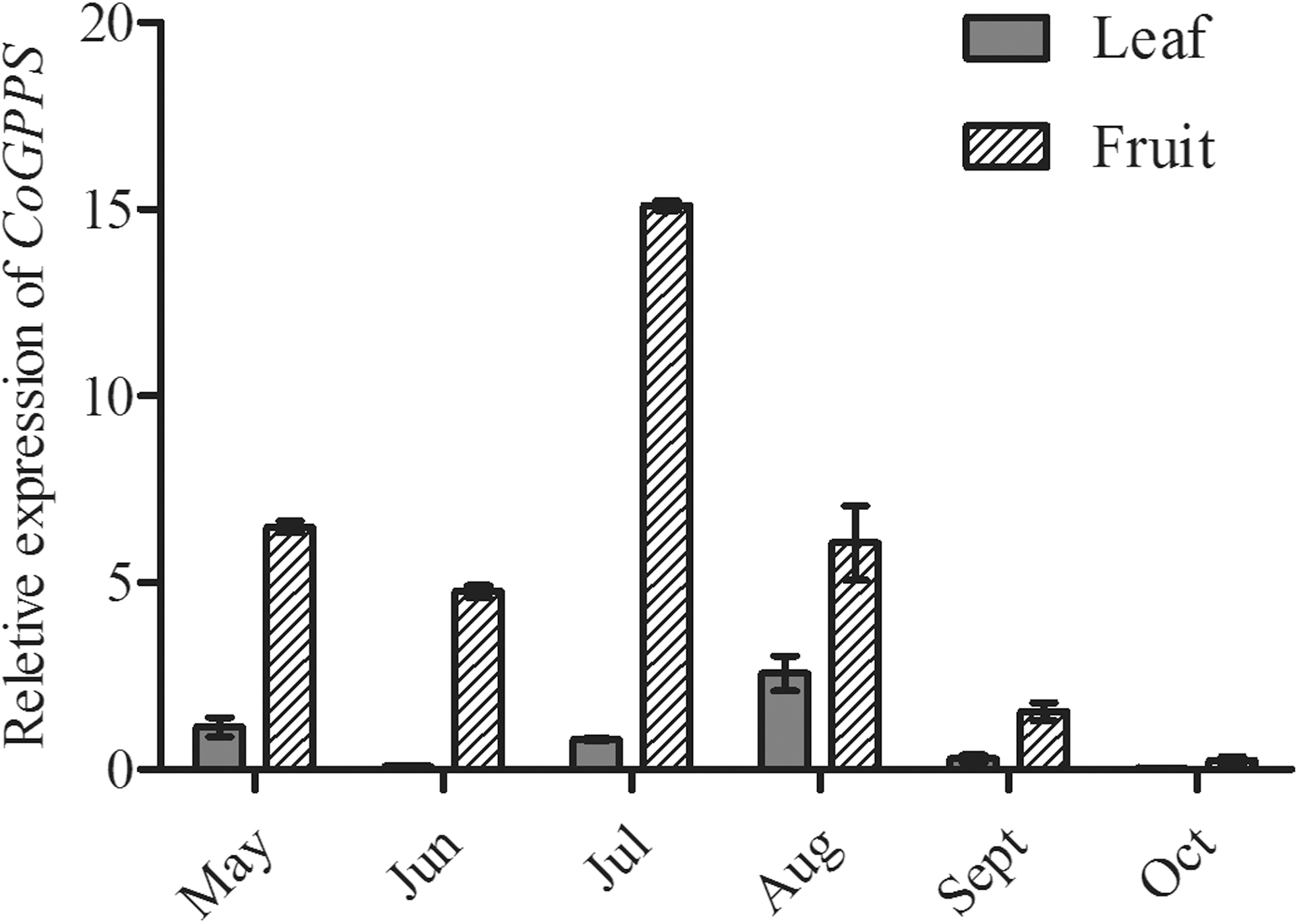
Analysis of the spatiotemporal expression pattern of CoGPPS in C. officinalis leaves and fruits from May to October.
Temporal analysis of expression of the CoGPPS gene in the fruits showed that the gene expression level gradually increased from May and reached its peak in July, while it decreased significantly in August.
Subcellular localization of CoGPPS
In silico analysis showed that the predicted CoGPPS protein was localized in the chloroplast. To validate this result, the protein sequence of CoGPPS was fused to the pCAMBIA1300-GFP vector, and the recombinant plasmid was transiently expressed in the N. benthamiana leaves. As observed using a laser confocal microscope, green fluorescence of the recombinant vector coincided with autofluorescence of the chloroplast, indicating that green fluorescence of CoGPPS was detected in the chloroplast (Fig. 4).

Subcellular localization of the CoGPPS-GFP protein in tobacco leaves.
Changes in loganin and morroniside contents in leaves and fruits of C. officinalis
The loganin and morroniside contents in the various tissues of C. officinalis varied remarkably. As shown in Figures 5 and 6, these compounds were more abundant in the fruits than in the leaves. This result indicated that loganin and morroniside were mainly synthesized in the fruits. The loganin content was higher when the fruit was immature, declined slightly during the ripening period, and finally stabilized, but it gradually decreased in the leaves. The morroniside content in the fruits changed significantly; that is, it first increased and then decreased, but its content in the leaves was extremely low.

Analysis of the loganin content in the leaf and fruit of C. officinalis by HPLC. Different lowercase letters indicate significant differences between treatments (p < 0.05). HPLC, high-performance liquid chromatography.
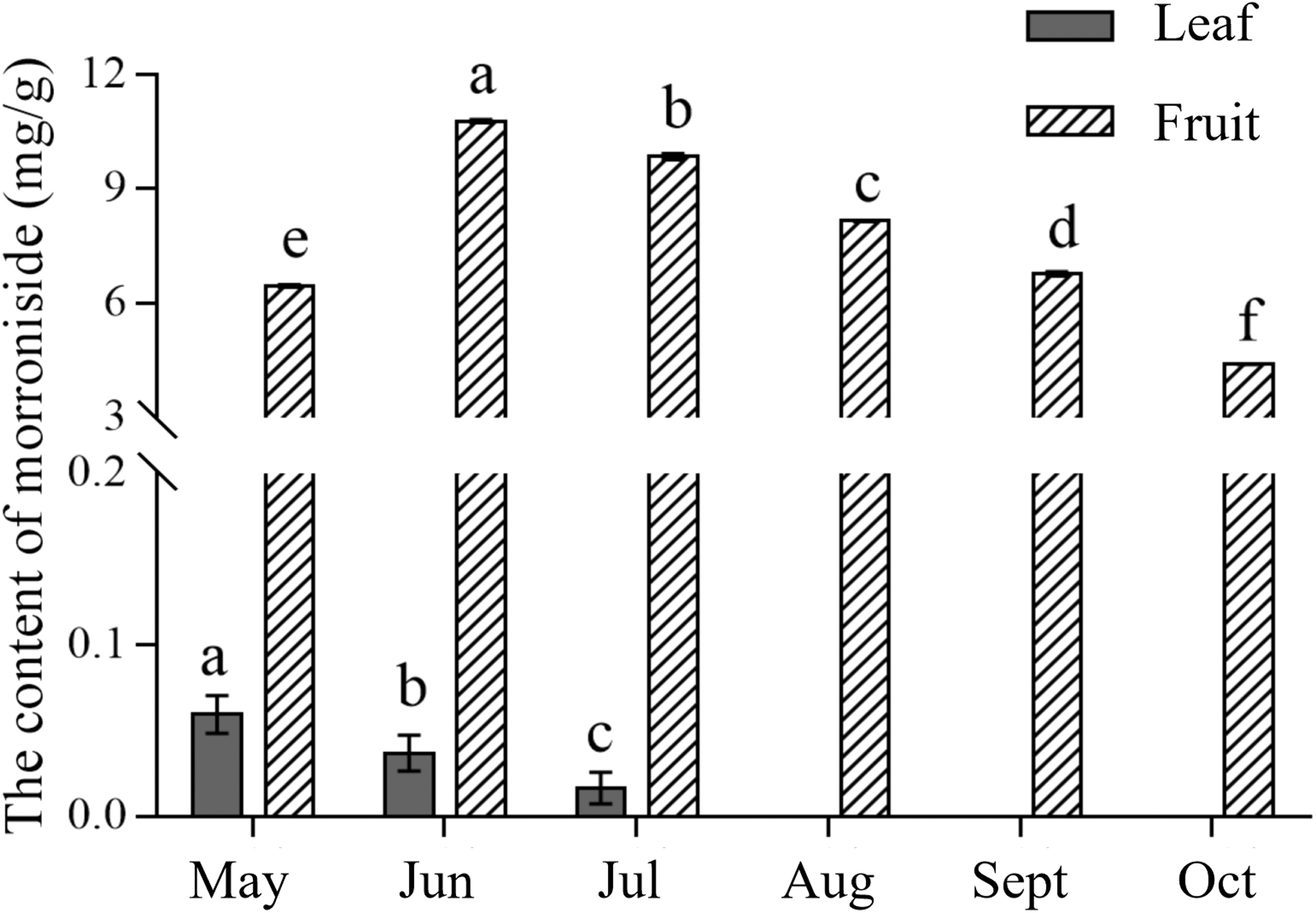
Analysis of the morroniside content in the leaf and fruit of C. officinalis by HPLC. Different lowercase letters indicate significant differences between treatments (p < 0.05).
Discussion
CIGs represent one of the most abundant groups of active compounds in C. officinalis, among which loganin and morroniside are the primary active compounds. CIGs have anti-inflammatory, hypoglycemic, antibacterial, and other activities (Chen et al., 2016). The key enzymes in the biosynthesis of CIGs have gradually become the focus of scientific research due to their importance in regulating the synthesis of CIGs.
Short-chain prenyl diphosphate synthases use IPP and DMAPP as substrates to synthesize direct precursors of CIGs. With reference to the transcriptome of C. officinalis (Hou et al., 2018), we cloned the CoGPPS gene and conducted a sequence analysis. Subsequent analysis revealed that the CoGPPS gene belonged to the isoprenoid synthase superfamily.
Most GPPS genes have tissue-specific expression and may be involved in regulating the synthesis of monoterpenoids. For example, the expression level of the CpGPPS.SSU1 gene in Chimonanthus praecox was higher in the petal and flower, but lower in the pistil, leaf, and stamen (Kamran et al., 2020). The expression pattern of TwGPPS.SSU ΙΙ was highest in the root, followed by those in the leaf, stem, and flower (Su et al., 2019). In this study, expression levels of CoGPPS in the leaf and fruit were analyzed by qRT-PCR.
The results indicated that the CoGPPS gene expression was higher in the fruit, but lower in the leaf. In addition, the transcript level of CoGPPS gradually increased during the growth stage of the fruit and slowly decreased as the fruit matured. These phenomena indicated that CoGPPS may be involved in regulating the development of fruits and accumulation of iridoid glycosides in C. officinalis.
Most of the enzymes of the MEP pathway have transit peptides that locate in plastids, such as LcGPPS.SSU1 and CpGPPS/GGPPS (Kamran et al., 2020; Zhao et al., 2020). In previous studies on GPPS protein localization, transient expression analysis has been commonly used to confirm the localization of GPPS. Studies have shown that LeGPPS is located in the cytoplasm, and this gene may be an acquired mutation of LeFPPS in the cytoplasm, which changed the enzymatic activity of LeGPPS (Ueoka et al., 2020).
In the present study, analysis of the transient expression revealed that the CoGPPS protein had strong GFP fluorescence in the chloroplast, indicating that the CoGPPS gene may be a subunit of a homodimer or heterodimer. The location of CoGPPS in chloroplasts further implied that the function of the CoGPPS enzyme was involved in the biosynthesis of monoterpenes.
In plants, synthesis of CIGs is a spatially and developmentally regulated phenomenon (Xue et al., 2019). Maturity is one of the main factors affecting the composition and content of active ingredients (Xu et al., 2013). The fruit development of C. officinalis is divided into three stages by change of color (Ahn et al., 2017). Loganin levels in C. officinalis also change during fruit ripening and decrease as the fruit matures (Ahn et al., 2017).
In the current study, loganin and morroniside were more abundant in the fruit than in the leaf. In addition, these compounds showed an initial increase and then decrease in the fruits. Studying the content of effective compounds at different developmental stages is helpful to determine the quality of C. officinalis.
Footnotes
Acknowledgment
The authors extend their special thanks to Han Guo for her contribution to this study and for collecting experimental materials for this study.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the National Natural Science Foundation of China (U1404829), a key project at the central government level: The Ability Establishment of Sustainable Use for Valuable Chinese Medicine Resources (2060302), and the Natural Science Foundation of Henan Province (202300410151). This research was also supported by a special fund for construction of technical systems of the traditional Chinese medicine industry in Henan Province (2023–24) and the Henan Province Chinese medicine industry science and technology commissioner service group.