
Abstract
Major facilitator superfamily domain containing 12 (MFSD12) regulates lysosomal cysteine import and promotes the proliferation and survival of melanoma cells. However, the expression and function of MFSD12 in other cancers, particularly in lung cancer, remain unclear. The expression of MFSD12 across various types of cancers and corresponding control tissues was examined using TIMER. MFSD12 expression in lung adenocarcinoma (LUAD) and its correlation with distinct clinicopathological features of LUAD patients were analyzed with UALCAN. The correlation between MFSD12 expression and survival of LUAD patients was assessed using the R package, survival, and the relationship between MFSD12 expression and immune infiltration status in LUAD was investigated using CIBERSORT. In addition, MFSD12 expression was knocked down in PC9 LUAD cells and their proliferation, capacity for expansion, cell cycle, apoptosis, and migration/invasion were evaluated through CCK-8 assays, colony formation assays, 7-AAD staining, Annexin V/PI staining, and Transwell assays, respectively. The stemness of these PC9 cells was determined through Western blotting, flow cytometry, and tumor sphere formation assays. MFSD12 mRNA levels were significantly elevated in multiple types of cancers, including LUAD. MFSD12 expression was also positively correlated with cancer stage, nodal metastasis, and infiltration of various immune cells in LUAD, and high MFSD12 levels predicted poor survival among LUAD patients. Knockdown of MFSD12 in PC9 cells resulted in decreased proliferation, attenuated colony formation capacity, cell cycle arrest, elevated apoptosis, impaired migration/invasion, and reduced stemness in PC9 cells. MFSD12 is an oncogene in LUAD.
Introduction
Lung cancer, being the most prevalent kind of malignant neoplasm, has shown a progressive rise in both morbidity and fatality rates on an annual basis (The, 2019). Nonsmall cell lung cancer (NSCLC) makes up about 84% of all lung cancer cases (Ganti et al., 2021). The most prevalent type of NSCLC is lung adenocarcinoma (LUAD), which accounts for 40% of cases. Lung squamous cell carcinoma (LUSC) comes in second at 25% to 30% and large cell carcinoma comes in third at 5% to 10% (Alduais et al., 2023). Lung cancer patients are often diagnosed at an advanced stage because there are no obvious symptoms in the early stages (Luo et al., 2021), resulting in a poor prognosis.
Moreover, despite the development of various targeted drugs for treating lung cancer, the 5-year survival rate for individuals diagnosed with lung cancer continues to be comparatively poor as a result of the development of medication resistance, recurrence of tumors, and spread of cancer cells to other parts of the body (Ashrafi et al., 2022; Gong et al., 2021; Luo et al., 2021). Hence, it is imperative to promptly ascertain new druggable targets and enhance therapeutic approaches to improve the prognosis of lung cancers, including NSCLC.
Major facilitator superfamily domain containing 12 (MFSD12) is a transmembrane protein that regulates the entry of cysteine into melanosomes and lysosomes (Adelmann et al., 2020). Recently, it has been reported that MFSD12 is involved in glycosphingolipid metabolism by modulating lysosomal homeostasis (Hong et al., 2023). Additionally, MFSD12 has been found to be overexpressed in melanoma, where it promotes the proliferation of melanoma cells (Wei et al., 2019). However, the role of MFSD12 in other physiological and pathological processes, such as various cancers, and the potential mechanisms by which it fulfills its functions remain unexplored.
This work investigated the expression of MFSD12 in different types of cancers using bioinformatic methods. Specifically, our analysis focused on MFSD12 expression in LUAD and its correlation with the clinicopathological features, prognosis, and immune infiltration status of LUAD. Additionally, we conducted functional studies on MFSD12 in PC9 LUAD cells. Our data revealed that MFSD12 is overexpressed in LUAD and exhibits an oncogenic function in LUAD cells. These findings significantly contribute to our understanding of the role of MFSD12 in cancer development.
Methods
Database analysis
The TCGA-LUAD data, encompassing gene transcription data from LUAD patients and clinicopathological information, were obtained from UCSC Xena (http://xena.ucsc.edu/). The correlation between MFSD12 levels and patient prognosis was determined using Kaplan–Meier analysis with the R package, survival (v3.2–13) (Goldman et al., 2020). The expression of MFSD12 across various cancer types was assessed using TIMER (https://cistrome.shinyapps.io/timer/) (Li et al., 2017).
The association between MFSD12 and clinicopathological features in lung cancer was investigated using UALCAN (http://ualcan.path.uab.edu/analysis.html), a widely utilized tool for tumor subgroup gene expression and survival analysis (Chandrashekar et al., 2017). The immune infiltration status of LUAD samples was evaluated by CIBERSORT (Chen et al., 2018). This study did not have an Institutional Review Board (IRB) approval or exemption statement or clinical trial registration number because the data set used for the bioinformatics analysis was derived from TCGA-LUAD, and the study was not a clinical trial and did not use human tissue and animals.
Cell culture and siRNA transfection
All cell lines (A549, H1299, and PC9) were sourced from XIAMEN Anti-hela Biological Technology. A549 cells were cultured in DMEM with 10% fetal bovine serum (Gibco), while H1299 and PC9 cells were cultured in RPMI 1640 medium with 10% fetal bovine serum. These cells were maintained in a humidified incubator at 37°C with 5% CO2. For siRNA transfection in PC9 cells, we utilized both control (siCtrl) and MFSD12-targeting siRNAs (siMFSD12), and transfection was performed using the X-tremeGENE siRNA Transfection Reagent (Roche). The siRNA sequences are listed in Table 1.
siRNA Sequences of Major Facilitator Superfamily Domain Containing 12
Western blotting assay
The lung cancer cells were subjected to lysis in RIPA buffer supplemented with protease inhibitors, followed by complete homogenization. Following centrifugation, the supernatant was collected, and the overall protein concentration was quantified using a bicinchoninic acid (BCA) assay. In brief, firstly, according to the instructions of the BCA kit (Beyotime), the BSA standard was diluted gradient, the BCA working solution was prepared. The standard at each dilution concentration and the sample to be tested were added to the 96 well plate, then the BCA working solution was added respectively, mixed, and incubated at 37°C for 30 min. Finally, the absorbance value at the wavelength of 562 nm was measured using a microplate reader. Subsequently, the protein samples were denatured and separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis. The separated proteins were then transferred onto PVDF membranes, which were subsequently blocked using a 5% nonfat milk/TBST solution.
After blocking, the membranes were treated with primary antibody solutions for one night at 4°C. They were then washed three times with TBST. Next, the membranes were incubated with a secondary antibody solution. Finally, the membranes were washed with TBST three additional times and incubated with an ECL working solution. The LAS-3000 Luminescent Image Analyzer (Fujifilm) was used to visualize the signals.
The antibodies used for Western blotting are as follows: MFSD12 antibody (1:500, PA5-42677; Invitrogen), GAPDH antibody (1:5000, 10494-1-AP; Proteintech), OCT4 antibody (1:5000, 11263-1-AP; Proteintech), Nanog antibody (1:3000, 14295-1-AP; Proteintech), C-MYC antibody (1:5000, 10828-1-AP; Proteintech), STAT3 antibody (1:5000, 10253-2-AP; Proteintech), phospho-STAT3 (P-STAT3) antibody (1:1000, #9131; Cell Signaling Technology), JAK2 antibody (1:1000, #3230; Cell Signaling Technology), phospho-JAK2 (P-JAK2) antibody (1:1000, #4406; Cell Signaling Technology), β-actin (1:5000, 20536-1-AP; Proteintech), and HRP-conjugated goat anti-rabbit IgG (1:2000, SA00001-2; Proteintech).
qPCR assay
Following the protocol provided by the manufacturer, total RNA was isolated from lung cancer cells using the TRIzol reagent (Invitrogen). Then, cDNA was synthesized using the Reality Reverse Transcriptase cDNA Synthesis Kit (Molecular Innovations) with 1 ng of RNA as a template. For qPCR, the SYBR Green PCR Master Mix (Invitrogen) was employed, and the relative expression levels of MFSD12 were calculated using the 2−ΔΔCq method. The primers used for qPCR analysis are listed in Table 2.
Major Facilitator Superfamily Domain Containing 12 Gene Primers for Quantitative Polymerase Chain Reaction Analysis
Colony formation assay
Cells transfected with siCtrl or siMFSD12 were seeded at a density of 1000 cells per well in six-well plates and grown for 14 days. Then, the cells were washed with PBS and fixed for 30 min in 4% paraformaldehyde. After fixation, they were stained with 0.5% crystal violet for 15 min. After that the cells were washed, and an IX73 inverted microscope (OLYMPUS) was used to take pictures of the colonies, which were then quantified using ImageJ software (NIH).
Cell proliferation assay
Cell proliferation was measured using the CCK-8 assay. Briefly, PC9 cells were planted at an average density of 1000 cells per well in 96-well plates. After 24 h, either control or MFSD12-targeting siRNA was transfected into the cells. Following an additional 24-h incubation, CCK-8 solution (C0037; Beyotime) was added to the 96-well plate on the 1st, 2nd, 3rd, 4th, and 5th day to measure cell proliferation. OD450 values were determined using an Infinite F50 microplate reader (Tecan).
Cell cycle assay
To examine the cell cycle, PC9 cells were seeded in six-well plates at a density of 1 × 106 cells per well. After 24 h, cells were transfected with either control or MFSD12-targeting siRNA. Following an additional 24-h incubation, the cells were treated with prechilled ethanol overnight at −20°C. Subsequently, they were washed with PBS and incubated with a 7-AAD/PBS solution in the dark for 30 min at 4°C. Finally, a BD FACSCanto™ II machine was used for flow cytometry analysis, and the results were processed with FlowJo software (v10).
Apoptosis assay
Following the manufacturer's instructions, the FITC Annexin V Apoptosis Detection Kit with propidium iodide (PI) (#640914; BioLegend) was used to detect apoptotic cells. In brief, PC9 cells were seeded in six-well plates at a density of 1 × 106 cells per well and cultured for 24 h. Subsequently, cells were transfected with either siCtrl or siMFSD12. After 24 h, the cells were washed with PBS, digested with a 0.25% trypsin solution, pelleted, resuspended in Annexin V binding buffer, and treated with the Annexin V-FITC/PI working solution.
Finally, flow cytometry analysis was performed using a BD FACSCanto II machine, and the results were analyzed using FlowJo software (v10).
Transwell assay
Cell migration and invasion were studied using Transwell assays in 24-well plates equipped with 8-μm Transwell chambers (#3422; Corning). For the migration assay, cells transfected with either siCtrl or siMFSD12 were resuspended in serum-free culture medium and seeded into the top chamber at a density of 1 × 105 cells per well, while the bottom chamber was filled with 700 μL of complete culture medium. After 48 h, the cells were fixed with methanol for 30 min, followed by staining with 0.05% crystal violet (Solarbio) for 5 min.
Finally, an IX73 inverted microscope (OLYMPUS) was used to observe and image the cells, and images were analyzed using ImageJ (NIH). For the invasion assay, all experimental procedures were the same as those for the migration assay, except that the top chambers were precoated with matrix glue for 2 h at 37°C.
Sphere formation assay
PC9 cells were inoculated in 24-well plates at 1000 cells per well. Cells were grown in DMEM/F12 media with B27 (Gibco; 17504044), EGF (20 ng/mL; Novoprotein), bFGF (20 ng/mL; Novoprotein), and insulin (4 μg/mL; Gibco). After a culture period of 12–18 days, the resulting spheres were observed and recorded using an IX73 inverted microscope (OLYMPUS).
Flow cytometry assay
After being washed with PBS, the cells were digested, pelleted, and centrifuged at 300 g for 5 min. The cells were resuspended after centrifugation and treated with FITC-conjugated mouse monoclonal antibodies against human CD44 (397517; BioLegend), EpCAM (324203; BioLegend), or isotype control antibodies (401205; BioLegend). A BD FACScan flow cytometer was used to identify the signal. To set a threshold for background fluorescence, unstained cells were employed as a negative control.
Statistical analysis
The differential expression of MFSD12 was assessed using the Wilcoxon matched-pairs signed rank test. Overall survival (OS) was analyzed using the Kaplan–Meier method and log-rank test. Correlation analysis was conducted using Pearson's correlation coefficient. Unpaired Student's t-tests and one-way ANOVA were employed for comparisons between two groups and multiple groups, respectively, utilizing GraphPad Prism (v8).
All experimental results are presented as the mean ± SEM, with statistical significance set at p < 0.05. All experiments in this study were conducted in triplicate for the purpose of quantification.
Results
MFSD12 expression is elevated in various cancers and positively associated with cancer stage and nodal metastasis in LUAD
We used the TIMER database to study the expression of MFSD12 across different cancers. The results showed that MFSD12 expression was significantly increased in a variety of cancers, including two types of lung cancer: LUAD and LUSC (Fig. 1A). Subsequently, we evaluated the association between MFSD12 expression and the clinicopathological features of LUAD samples using the UALCAN online database.
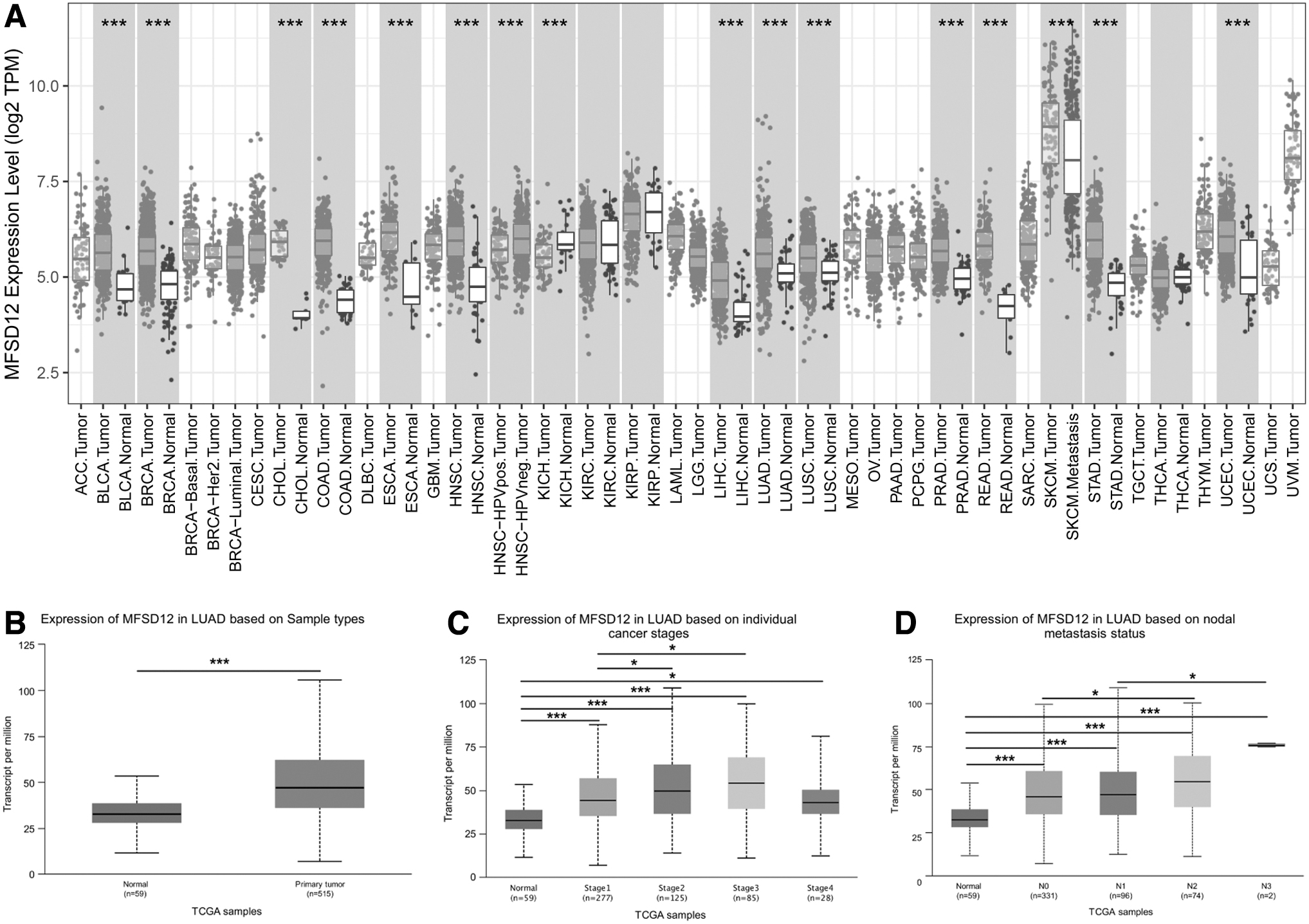
Expression of MFSD12 across various types of cancers and its association with clinicopathological features of LUAD.
The findings confirmed elevated MFSD12 expression in LUAD samples (Fig. 1B) and revealed a significant positive correlation with both cancer stage and nodal metastasis in LUAD (Fig. 1C, D). Based on the results presented, it can be observed that MFSD12 potentially plays a role in facilitating LUAD progression.
High MFSD12 expression predicts poor OS in LUAD patients and correlates with increased immune cell infiltration in LUAD tissues
To assess the prognostic value of MFSD12 expression in LUAD, we analyzed the association between MFSD12 expression and OS status using data from 510 LUAD patients, with relevant clinical information obtained from UCSC Xena. According to the median expression of MFSD12, patients were divided into the high expression group and low expression group. Employing the Kaplan–Meier method, we observed a significant correlation between high MFSD12 levels and a worse prognosis in LUAD (Fig. 2A).
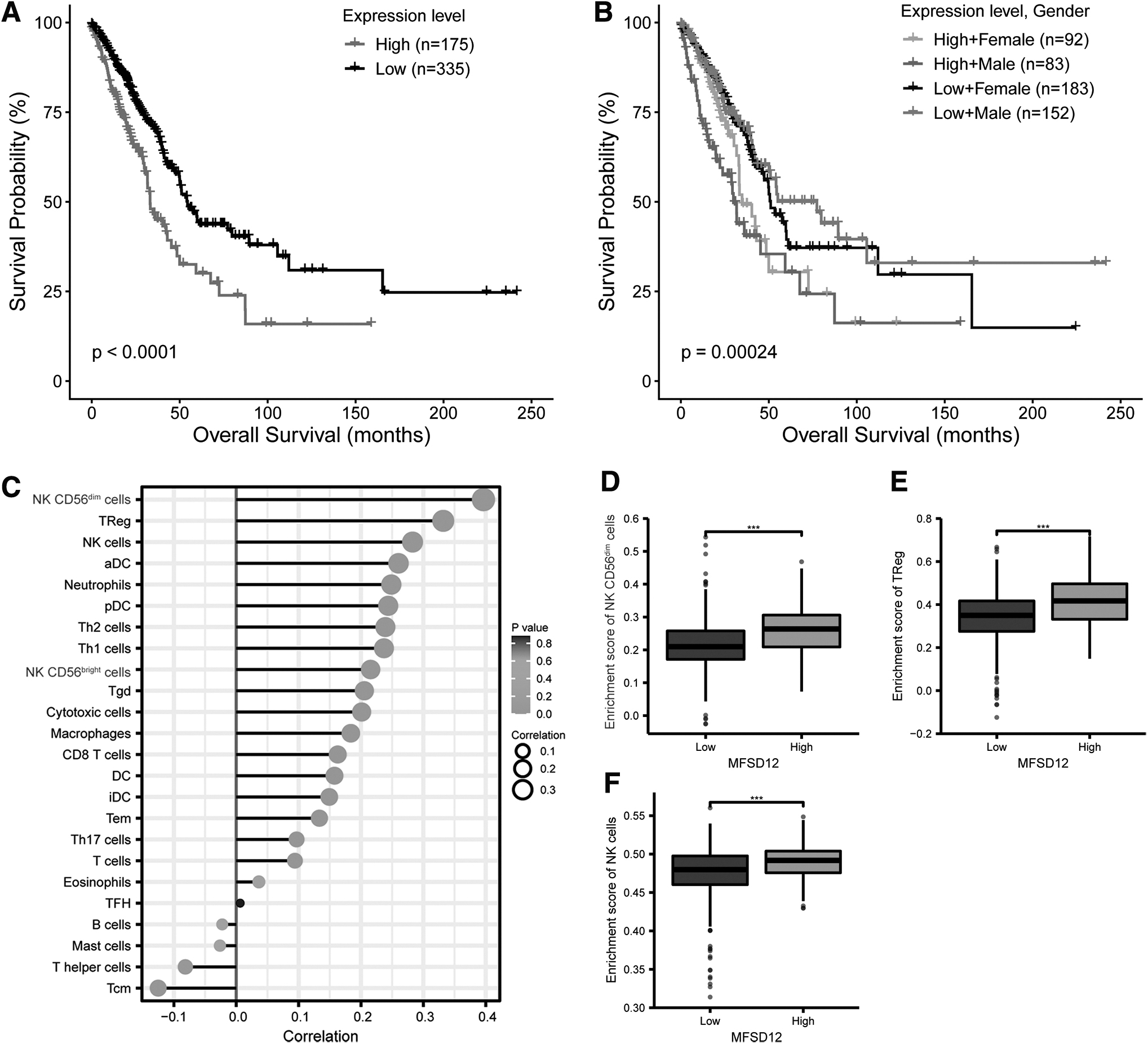
Correlation between MFSD12 expression and survival of LUAD patients and infiltration of immune cells in LUAD.
Furthermore, within the group of LUAD patients expressing high or low levels of MFSD12, male patients exhibited better OS than female patients (Fig. 2B). Additionally, an examination was conducted to explore the correlation between the expression of MFSD12 and infiltration of immune cells in LUAD using CIBERSORT. The results unveiled a significant positive correlation between MFSD12 expression and multiple types of immune cells (Fig. 2C).
Notably, high MFSD12 expression exhibited the most significant associations with NK CD56dim cells, regulatory T (Treg) cells, and NK cells (Fig. 2D–F). These findings further support the notion of MFSD12's oncogenic potential in LUAD.
Knockdown of MFSD12 reduced proliferation, attenuated colony formation capacity, impaired the cell cycle, elevated apoptosis, and mitigated the migration and invasion of PC9 LUAD cells
To investigate the function of MFSD12 in LUAD, we initially depleted MFSD12 in PC9 cells, a commonly used LUAD cell line with an exon 19 deletion in the EGFR gene (Ono et al., 2004). qPCR and Western blotting confirmed that all three siRNAs targeting MFSD12 efficiently reduced MFSD12 expression in PC9 cells, with siMFSD12–2 showing the highest knockdown efficiency (Fig. 3A–C). Therefore, we selected siMFSD12–2 (hereafter referred to as siMFSD12) for subsequent experiments.
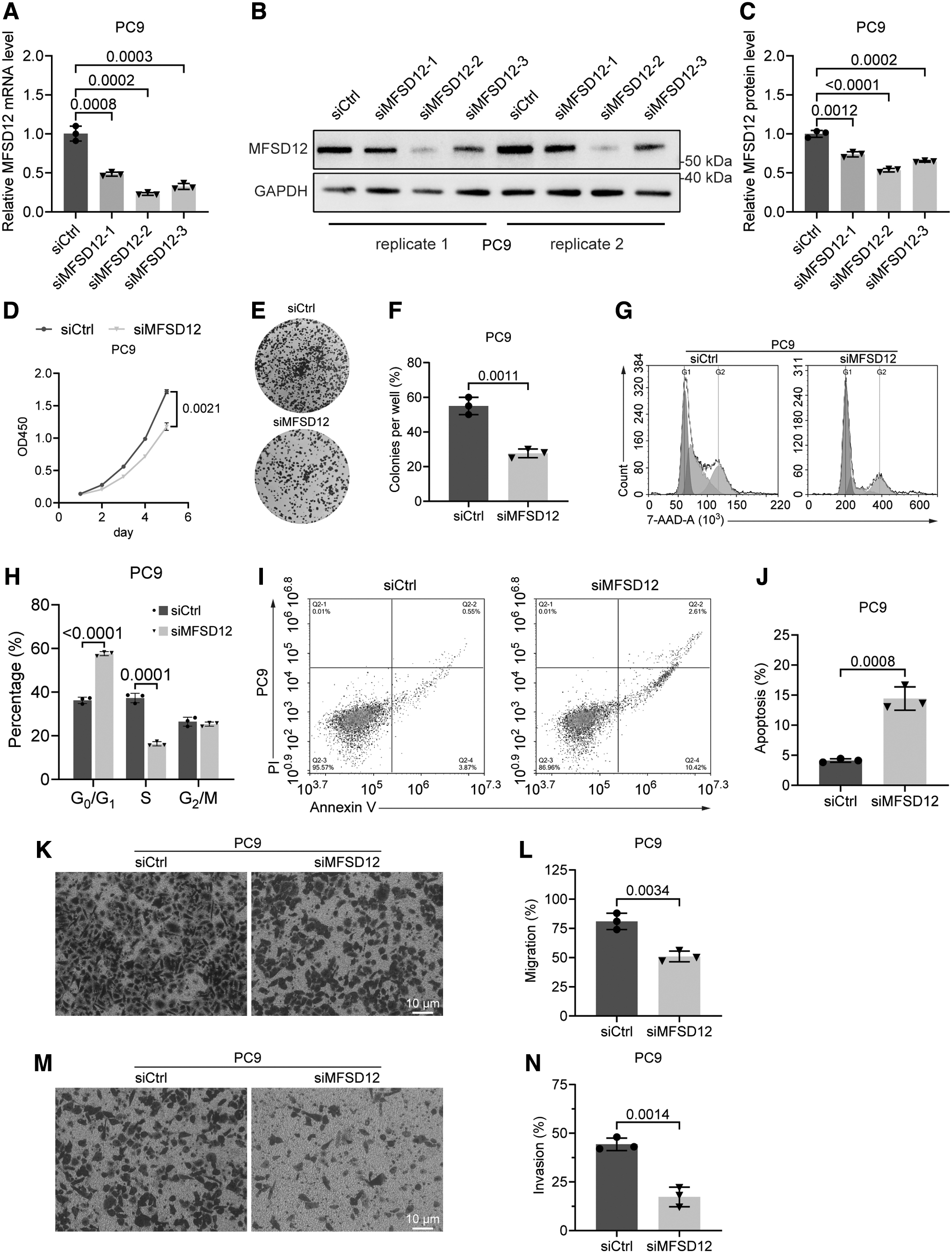
The effect of MFSD12 deficiency on PC9 cells.
A CCK8 test was performed to evaluate the effect of MFSD12 knockdown on proliferation of PC9 cells, which revealed a significant decrease in proliferation in MFSD12-deficient PC9 cells compared with control cells (Fig. 3D). The data obtained from the colony formation assay provided additional evidence that the absence of MFSD12 significantly compromised the ability of PC9 cells to generate colonies (Fig. 3E, F). Consistent with these findings, MFSD12 depletion led to cell cycle arrest and an increased level of apoptosis in PC9 cells, as indicated by 7-AAD and Annexin-V/PI staining results (Fig. 3G–J).
Furthermore, results from the Transwell experiment demonstrated a significant decrease in the migratory and invasive capabilities of PC9 cells following the knockdown of MFSD12 (Fig. 3K–N). Collectively, these observations provide strong evidence of the oncogenic role of MFSD12 in LUAD.
MFSD12 deficiency impaired the stemness of PC9 cells
We then assessed the impact of MFSD12 deficiency on the stemness of PC9 cells. Western blotting results revealed a significant downregulation of OCT4, Nanog, and C-MYC expression in PC-9 cells upon MFSD12 depletion (Fig. 4A, B). Additionally, phosphorylation of STAT3 and JAK2, known to maintain the undifferentiated state of mouse embryonic stem cells and frequently elevated in cancers (Matsuda et al., 1999; Mengie Ayele et al., 2022; Wang et al., 2022), was remarkably reduced in MFSD12-deficient PC9 cells (Fig. 4A, B).
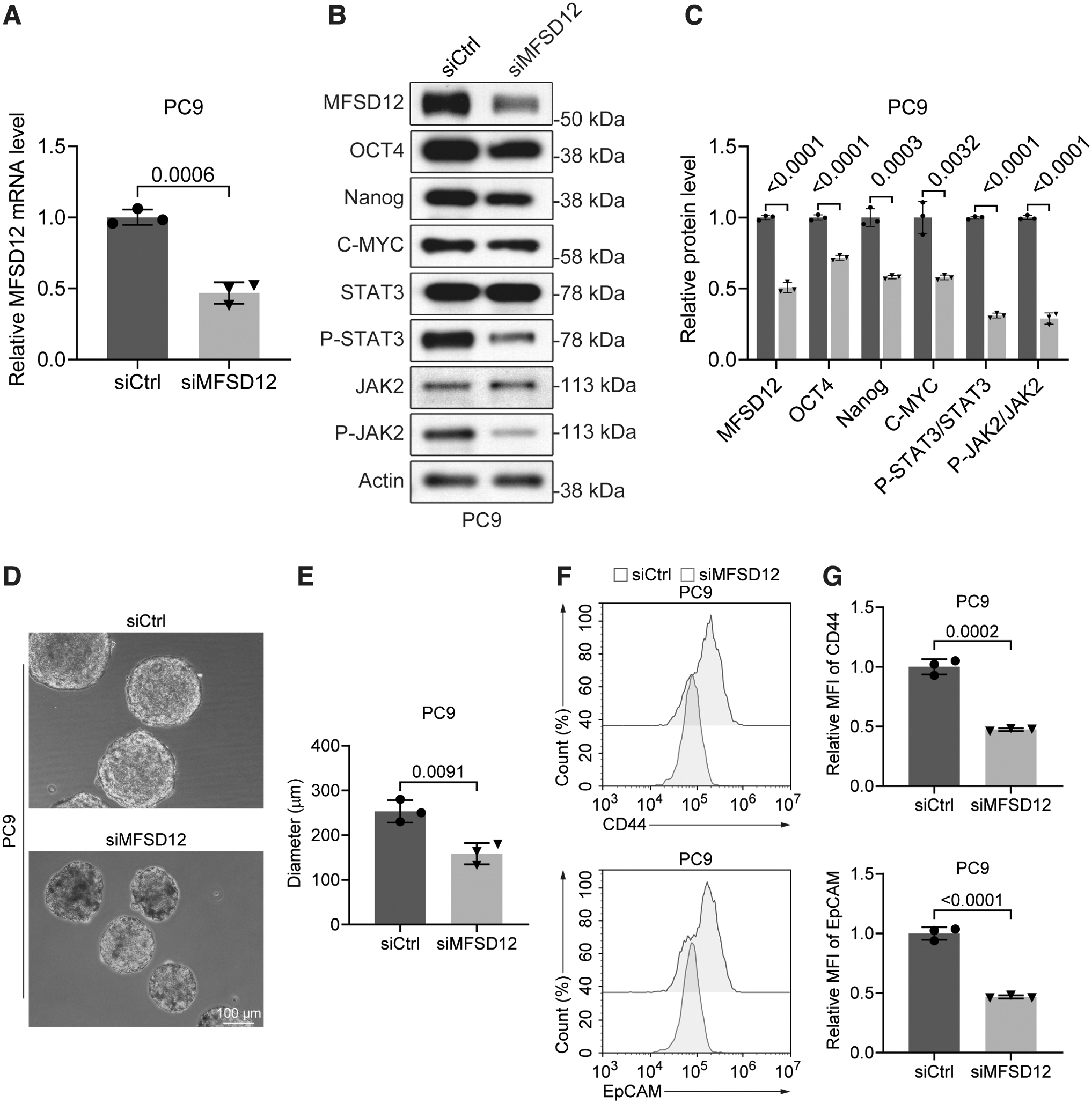
Impact of MFSD12 deficiency on the stemness of PC9 cells.
Consistent with these findings, flow cytometry analysis revealed a reduction in the expression of the stem cell markers, CD44 and EpCAM, upon MFSD12 deficiency (Fig. 4C, D). Furthermore, results from the sphere formation assay, which measures the self-renewal capacity and multipotency of cancer cells (Bahmad et al., 2018; Johnson et al., 2013), indicated that MFSD12 deficiency significantly mitigated the stemness of PC9 cells. These findings demonstrate that MFSD12 may promote LUAD by maintaining the stemness of cancer cells.
Discussion
The major facilitator superfamily (MFS) is recognized as the most extensive superfamily of secondary active transporters, playing a crucial role in the ATP-independent transportation of small molecules across cellular membranes (Drew et al., 2021). These transporters are ubiquitously expressed and play crucial roles in controlling growth, metabolism, and homeostasis in all organisms (Drew et al., 2021; Pao et al., 1998). Among the MFS members, MFSDs are orphan transporters and their functions are not yet well characterized (Drew et al., 2021).
However, accumulating evidence points to the involvement of MFSDs in cancer development. For instance, MFSD2A, a member of the MFSD family, is significantly downregulated in lung cancer cells and LUAD, functioning as a tumor suppressor gene by regulating cell cycle and matrix attachment (Spinola et al., 2010). MFSD2A may also inhibit gastric cancer progression by suppressing angiogenesis (Shi et al., 2018). MFSD4, another MFSD family member, exhibits reduced expression in gastric cancer with hepatic metastasis and serves as a predictor of hepatic recurrence and metastasis risk in gastric cancer (Kanda et al., 2016).
Furthermore, MFSD4A expression is decreased in nasopharyngeal carcinoma and inhibits its malignant progression by degrading EPHA2 (Yang et al., 2022). Conversely, MFSD12, as previously mentioned, promotes melanoma cell proliferation (Wei et al., 2019). Consistent with this, our data showing MFSD12 overexpression in multiple cancers and its oncogenic role in LUAD further support the notion that MFSD12 acts as an oncogene. This apparent discrepancy in the functions of different MFSDs in cancer may be attributed to their distinct cellular distributions and the substrates they transport.
The precise mechanism by which MFSD12 facilitates cancer progression remains unclear. However, considering that MFSD12 is essential for cysteine import into lysosomes in nonpigmented cells (Adelmann et al., 2020), it is plausible that MFSD12 influences cancer development by modulating cysteine balance within the lysosome. This is important because disrupted cysteine accumulation in the lysosome can affect cellular ROS levels (Sumayao et al., 2016), which in turn may potentially play a role in progression of cancer (Liou and Storz, 2010).
Moreover, considering the diverse roles of cysteine in promoting cancer cell proliferation (Combs and DeNicola, 2019), further investigation is needed to elucidate the exact role of MFSD12-mediated cysteine import into lysosomes in cancer progression.
In addition to regulating the proliferation of LUAD cells, our findings also shed light on the impact of MFSD12 on the stemness of these cells. Since MFSD12 regulates cysteine levels, which are known to promote stem cell proliferation and function by modulating cellular redox levels (Kong et al., 2018; Wang et al., 2013), we hypothesize that MFSD12 may maintain the stemness of LUAD and other cancer cells through similar mechanisms.
Moreover, Bhardwaj et al. (2023) reported that mice inoculated with DC661-treated cells developed adaptive immunity and tumor rejection in “immune hot” tumors, but not in “immune cold” tumors. While DC661-induced LLP was critical for LMP-mediated immunogenic cell death, this process was reversed by the lysosomal transporter, MFSD12, which imported cysteine into lysosomes (Bhardwaj et al., 2023). We all know that the catabolic and degrading capabilities of endolysosomal systems are well utilized in mammalian immune responses (Saftig and Puertollano, 2021).
Therefore, in our opinion, the relationship between MFSD12 and infiltration of NK CD56dim and NK cells in LUAD may rely on the process that MFSD12 may change the lysosomal systems in cancer cells, thereby mediating the infiltration of NK CD56dim and NK cells and finally influencing tumor progression. However, direct evidence is required to verify this hypothesis, and further investigation is warranted to explore additional potential mechanisms by which MFSD12 influences cancer cell stemness.
Conclusions
In conclusion, our findings highlight the overexpression and oncogenic role of MFSD12 in LUAD, underscoring MFSD12 as a potential druggable target for the treatment of this lethal disease.
Footnotes
Authors' Contributions
W.J.Z. contributed to the conception and design of the research, performed the research and data analysis, and drafted the manuscript. X.L.H. contributed to data analysis. Z.X.C. performed the research. X.J.L. contributed to the design of the study. W.J.Z. contributed to the design of the study and revised the manuscript. All authors read and approved the final manuscript.
Ethics Approval and Consent to Participate
Data collection and processes were performed according to policies of the GEO and TCGA project.
Disclosure Statement
No competing financial interests exist.
Funding Information
There was no funding support for this study.