
Abstract
Legg–Calve–Perthes disease (LCPD) is an idiopathic avascular necrosis of the pediatric femoral head. Bone remodeling and bone structural genes have the potential to contribute to the progression of LCPD when there is disequilibrium between bone resorption and bone formation. A case–control study was performed to search for associations of several common polymorphisms in the genes Receptor Activator for Nuclear Factor κappa B (RANK), Receptor Activator for Nuclear Factor κappa B Ligand (RANKL), osteoprotegerin (OPG), interleukin (IL)-6, and type 1 collagen (COL1A1) with LCPD susceptibility in Mexican children. A total of 23 children with LCPD and 46 healthy controls were genotyped for seven polymorphisms (rs3018362, rs12585014, rs2073618, rs1800795, rs1800796, rs1800012, and rs2586498) in the RANK, RANKL, OPG, IL-6, and COL1A1 genes by real-time polymerase chain reaction with TaqMan probes. The variant allele (C) of IL-6 rs1800795 was associated with increased risk of LCPD (odds ratio [OR]: 3.8, 95% confidence interval [CI]: [1.08–13.54], p = 0.033), adjusting data by body mass index (BMI) and coagulation factor V (FV), the association with increased risk remained (OR: 4.9, 95% CI: [1.14–21.04], p = 0.025). The OPG polymorphism rs2073618, specifically GC-GG carriers, was associated with a more than fourfold increased risk of developing LCPD (OR: 4.34, 95% CI: [1.04–18.12], p = 0.033) when data were adjusted by BMI-FV. There was no significant association between RANK rs3018362, RANKL rs12585014, IL-6 rs1800796, COL1A1 rs1800012, and rs2586498 polymorphisms and LCPD in a sample of Mexican children. The rs1800975 and rs2037618 polymorphisms in the IL-6 and OPG genes, respectively, are informative markers of increased risk of LCPD in Mexican children.
Background
Legg–Calve–Perthes disease (LCPD) is considered an idiopathic avascular necrosis of the pediatric femoral head (Azarpira et al., 2018; Pavone et al., 2019). LCPD is a rare disease that presents annual incidence of 0.4–29.0/100,000 children, under 15 years of age, presenting a peak incidence between 4 and 8 years of age and being five times more common in boys than in girls (Loder and Skopelja, 2011). There is a wealth of literature describing the possible etiological factors, considering the disruption of the blood supply to the femoral head epiphysis as a key event and proposing vascular, mechanical, genetic, thrombotic, environmental, and systemic conditions to explain the pathogenesis of LCPD (Kim and Herring, 2011).
The most-studied mechanism is the interruption of the blood supply to the femoral epiphysis, with evidence that this may be due to thrombophilia (an increased tendency for thrombus formation) or hypofibrinolysis (a reduced capacity for thrombolysis), processes that have been suggested to play essential roles in the pathogenesis of osteonecrosis (Vokkerla and Chidrawar, 2019). Studies have suggested a hereditary tendency toward hypercoagulable states and thrombophilic disorders as causes of vascular thrombosis, correlating them with the risk of developing LCPD (Basit and Khoshhal, 2022). Some studies have found that protein C (PC) and protein S (PS) deficiencies are common in LCPD patients, and other reports have identified an increase in the presence of the Factor V Leiden (FVLeiden) mutation in LCPD patients (Asadollahi, 2021). Because of their involvement with the risk of thrombosis, polymorphisms in the methylenetetrahydrofolate reductase gene (MTHFR) and in the prothrombin (PT) genes have been studied in relation to LCPD. The MTHFR rs1801133 polymorphism has been associated with LCPD risk in Mexican patients (Buendía-Pazarán et al., 2022) and with severe forms of the disease in Spanish children (García-Alfaro et al., 2021). Reports with the G20210A variant of the PT gene have presented contradictory results. Vosmaer et al. (2010) found an association with LCPD, while other studies have reported no association.
Structural bone genes have also been associated with the etiology of LCPD. Mutations in the type 2 collagen (COL2A1) gene have been reported in LCPD patients (Li et al., 2014; Su et al., 2008). Type 1 collagen (COL1A1) gene mutations or polymorphisms have been associated with several bone pathologies, but there is only one report about the association of the COL1A1 gene with LCPD (Buendía-Pazarán et al., 2022; Kostik et al., 2013; Xie et al., 2015).
The natural course of the pathogenesis of LCPD progresses through four phases: synovitis, avascular necrosis (femoral epiphysis condensation) and fragmentation (resorption), reossification, and remodeling (healing). In the healing phase, the femoral head may be remodeled until it reaches its normal shape or, by contrast, may present some mechanical alterations in varying degrees of intensity (Kim and Herring, 2011; Loder and Skopelja, 2011; Spasovski et al., 2023).
Alterations in bone metabolism can occur in several LCPD phases; therefore, genes involved in bone metabolism and regulation could participate in LCPD onset and progression. The bone remodeling pathway formed by the Receptor Activator for Nuclear Factor κappa B (RANK), the Receptor Activator for Nuclear Factor κappa B Ligand (RANKL), and osteoprotegerin (OPG), named the RANK-RANKL-OPG system, is the principal mechanism that controls the equilibrium of bone formation and bone resorption (Kenkre and Bassett, 2018). Interestingly, it has been shown in an animal model with femoral head deformity that bone resorption and deformity decrease with exogenous OPG administration (Kim et al., 2006). When the equilibrium is lost, several bone pathologies can occur, and a similar situation could be involved in LCPD. Polymorphisms in these genes have not been explored to assess the participation of these genes in LCPD.
An important process involved in LCPD is inflammation. This process plays an important role in the regulation of bone metabolism. One of the most-studied proinflammatory cytokines is interleukin-6 (IL-6), which stimulates osteoclast differentiation and bone resorption, both of which are key events in LCPD phases (Wang and He, 2020). The levels of proinflammatory cytokines, including IL-6, are elevated in patients with LCPD (Kamiya and Kim, 2020; Kamiya et al., 2015). This could be a link between thromboembolic risk factors in LCPD and the genes involved in bone remodeling. IL-6 is a modulator of skeletal homeostasis and osteoclast differentiation, as well as a mediator of coagulation, which is another important function that is relevant to this work. The role of IL-6 as a mediator of the coagulation system includes an increase in fibrinogen and platelet counts and the activation of an extrinsic coagulation cascade (Akbarian-Bafghi et al., 2021; Srzentić et al., 2014).
The association between polymorphisms of IL-6 in the promoter region in relation to LCPD has been studied in Iranian children, and an increased risk of developing LCPD was observed (Akbarian-Bafghi et al., 2021). On the other hand, no association was reported by Srzentić et al. (2014) in Serbian children. There are no reports of an association of IL-6 with the risk of LCPD in the Mexican population.
In view of this background, there is a possibility that several common polymorphisms in bone remodeling and bone structural genes, as well as coagulation factors, are associated with the risk of LCPD. The purpose of the present investigation was to search for associations of RANK, RANKL, OPG, IL-6, and COL1A1 gene polymorphisms with LCPD in Mexican children.
Materials and Methods
Ethics approval and consent to participate
This study was approved by the Ethics and Research Committee of the INR-LGII with registration number 54/16. All participants signed a written informed consent form. The controls were selected under the guidelines of the official Mexican standard NOM-253-SSA1-2012. All the procedures performed in this study were in accordance with the ethical standards of the Research and Ethical Committee of the INR-LGII and with the Declaration of Helsinki revised in 2013.
Study subjects
The study included a total of 69 children: 23 with LCPD (cases) and 46 without the disease (controls). The patients were diagnosed with LCPD using clinical and radiological criteria and controls were declared healthy (without LCPD) by the same radiological criteria and by their values of thrombosis and inflammation parameters which were within the reference values (Data set in Supplemental Data). The controls were matched with the LCPD patients according to age, sex, weight, and body mass index (BMI) at a ratio of 2:1. All participants who were diagnosed with other bone diseases and coagulation-related abnormalities, and those who were under drug treatment that modified bone metabolism, were excluded. The participants received oral and written information, and parents or guardians signed a letter of consent. All patients were recruited from the Orthopedic Service at the Instituto Nacional de Rehabilitación “Luis Guillermo Ibarra Ibarra.”
DNA extraction and genotyping
Genomic DNA was extracted from peripheral blood and stored at −80°C. The DNA concentration was measured with a Nanodrop ND-100 spectrophotometer (Thermo Fisher Scientific, Inc., Wilmington, DE). The single nucleotide polymorphisms (SNPs) included were RANK rs3018362 (G/A), RANKL rs12585014 (G/A), OPG rs2073618 (C/G), IL-6 rs1800795 (G/C) and rs1800796 (G/C), and COL1A1 rs1800012 (C/A) and rs2586498 (G/A). Genotyping was performed by real-time polymerase chain reaction (PCR) using predisigned and custom TaqMan probes (Thermo Fisher Scientific, Waltham, Massachusetts) in a StepOne Real-Time PCR System (Applied Biosystems, Foster City, CA). The real-time PCR was performed in a total mixture volume of 25 μL, containing 1 × TaqMan PCR master mix, probe at 100, 900 nM of each primer (Table 1 shows the primers sequences of the assays designed by the authors) and 25 ng of genomic DNA. The cycling conditions were as follows: denaturation at 95°C for 10 min, followed by 40 cycles at 92°C for 15 s and then 60°C for 1 min. Primers used to genotyping are shown in Table 1.
Sequence of the Primers Designed to Genotype the Samples with TaqMan Assays
RANKL 12585014, OPG 2073618, IL-6 1800796, COL1A1 1800012 and COL1A1 2586498, were predesigned assays and primers sequences are not available by the provider.
COL1A1, type 1 collagen; IL, interleukin; OPG, osteoprotegerin; RANK, Receptor Activator for Nuclear Factor κappa B; RANKL, Receptor Activator for Nuclear Factor κappa B Ligand.
Statistical analysis
The sample size calculated for this study was 21 cases and 21 controls, considering α = 0.05 (two sided), β = 0.20, expected difference between cases and controls = 0.35, without the Fleiss–Tytun–Ury continuity correction (Browner et al., 1993).
The results of the variables are described as the mean ± standard deviation 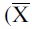 ± SD) or as percentages (%). The comparison of variables between controls and cases was performed using Student's t-test or Mann–Whitney U-test, when appropriate. The frequencies of genotypes and alleles of the polymorphisms were compared using the chi-squared test; Hardy–Weinberg equilibrium was calculated. The odds ratio (OR) and the 95% confidence interval (CI) were calculated to express risk, and the associations were considered significant with a p < 0.05. Statistical analysis was performed with PAST 4.03. Univariate and multivariate nonconditional logistic regression was performed to estimate the effect of each SNP under the inheritance models, (SNPStats. https://www.snpstats.net/start.htm).
± SD) or as percentages (%). The comparison of variables between controls and cases was performed using Student's t-test or Mann–Whitney U-test, when appropriate. The frequencies of genotypes and alleles of the polymorphisms were compared using the chi-squared test; Hardy–Weinberg equilibrium was calculated. The odds ratio (OR) and the 95% confidence interval (CI) were calculated to express risk, and the associations were considered significant with a p < 0.05. Statistical analysis was performed with PAST 4.03. Univariate and multivariate nonconditional logistic regression was performed to estimate the effect of each SNP under the inheritance models, (SNPStats. https://www.snpstats.net/start.htm).
Results
Characterization of study subjects
In total, 23 LCPD patients (male/female: 21/2; mean age: 16.87 ± 11.31 years) and 46 healthy subjects (male/female: 42/4; mean age: 16.37 ± 11.04 years) were included in the final analysis. The general characteristics of the study population are summarized in Table 2. There were no significant differences between cases and controls with respect to these variables.
General Characteristics of the Legg–Calve–Perthes Disease Cases and Controls
Variables expressed as mean ± standard deviation.
BMI, body mass index.
In Table 3, the determination of coagulation factors, anticoagulant proteins, and homocysteine (Hcy) is summarized. Hernández et al. carried out a study to determine the specific reference values for children of the coagulation factors FI, FII, FV, FVII, FVIII, FIX, FX, FXI, and FXII, as well as the anticoagulant proteins PC, antithrombin, and Hcy (Hernández-Zamora et al., 2023). The suggested reference values were determined from 200 healthy minors from México, according to the report by Monagle et al. (2006) to establish reference values. Using these values, in this work, significant differences in FV, FIX, and Hcy were found.
Determination of Coagulation Factors, Thrombophilic Factors and Homocysteine in Cases with Legg–Calve–Perthes Disease and Controls
Variables expressed as mean ± standard deviation; significant values are shown in bold.
AT, antithrombin; FV, factor V; Hcy, homocysteine; PC, C protein; VWF, Von Willebrand Factor.
Association of RANK, RANKL, and OPG polymorphisms with LCPD
The polymorphisms of interest in this work were in Hardy–Weinberg equilibrium. The RANK rs3018362 variant allele (A) was present in 50% of the children with LCPD and in 45% of the control group; the frequency of the AA genotype was 34.8% in cases with LCPD and 23.9% in the control group. This polymorphism did not show significant differences.
The RANKL rs12585014 variant allele was present in 33% of children with LCPD and in 37% of the control group; the frequency of the AA genotype was 4.3% in cases with LCPD and 13% in the control group. No significant differences were detected. The OPG rs2073618 variant allele C was present in 48% of the cases with LCPD compared to 37% of the controls. The GG genotype was present in 17.4% of the cases with LCPD and 15.2% of the controls. This polymorphism showed no significant trend toward high risk under a codominant model adjusted by BMI (Table 4). When data were adjusted by BMI and FV, it was found that carriers of the GC-GG genotypes had a fourfold increased risk of developing LCPD (OR: 4.34, 95% CI: 1.04–18.12, p = 0.03). When adjusting data by FIX and Hcy, the significance was lost (Table 5).
Association of RANK, RANKL, OPG, IL-6, and COL1A1 Polymorphisms with Legg–Calve–Perthes Disease
Significant values are shown in bold.
p-Value crude data.
p-Value adjusted by BMI.
CI, confidence interval; NA, not analyzed; OR, odds ratio.
Association of Polymorphisms with Legg–Calve–Perthes Disease Adjusted by Coagulation Factors
Significant values are shown in bold.
p-Value adjusted by BMI-FV.
p-Value adjusted by BMI-FIX.
p-Value adjusted by BMI-Hcy.
Association of IL-6 polymorphisms with LCPD
The IL-6 polymorphisms rs1800795 and rs1800796 were in Hardy–Weinberg equilibrium. The variant allele of rs1800795 was present in 24% of the cases with LCPD and in 10% of the controls. The IL-6 rs1800795 CC genotype was present in 8.7% of the LCPD cases and was absent in the controls. The C allele was associated with a higher risk of LCPD (OR: 2.89; 95% CI: 1.104–7.61; p = 0.03). In the dominant model, after adjusting data by BMI, it was found that carriers of the GC-CC genotypes had a 3.8-fold increased risk of developing LCPD (OR: 3.82, 95% CI: 1.08–13.55, p = 0.03; Table 4). Adjusting the data by BMI and FV, the association remained significant in the dominant model, and carriers of the GC-CC genotypes had an almost fivefold increased risk of developing LCPD (OR: 4.9, 95% CI: 1.14–21.04, p = 0.02, Table 5). When data were adjusted by FIX and Hcy, there was a trend toward a high-risk association, but significance was lost. For IL-6 rs1800796, the frequency of the variant allele (C) was 30% in cases and 39% in controls; this polymorphism did not show an association.
Association of COL1A1 polymorphisms with LCPD
The genotype frequencies of the COL1A1 polymorphisms rs1800012 and rs2586498 were in Hardy–Weinberg equilibrium. The variant allele (A) of COL1A1 rs1800012 was present in 7% of the cases with LCPD and in 8% of the controls. For the COL1A1 rs2586498 polymorphism, the frequency of the variant allele (A) was 11% in patients and 10% in controls. The AA genotype was not found in any group, and no significant associations were observed in the COL1A1 polymorphisms rs1800012 (C/A) and rs258698 (G/A).
Discussion
LCPD is a pediatric disease that affects the epiphysis of the femoral head and is considered avascular osteonecrosis of the femoral head. However, the etiology of LCPD is now considered multifactorial, involving the participation of genetic and environmental factors, coagulation disorders, and inflammation, among other factors (Asadollahi et al., 2021; Rodríguez-Olivas et al., 2022; Srzentić et al., 2015).
The pathophysiology of LCPD comprises four phases. In the fragmentation phase, there is excessive bone resorption related to the development of a severe deformity of the affected hip. Due to a predominance of osteoclasts and the replacement of necrotic bone with fibrovascular granulation tissue, the femoral head is more vulnerable to damage due to repeated loading; this imbalance is related to the collapse of the femoral head (Boyle et al., 2003; Pouya and Kerachian, 2015). An abnormal collagen turnover was found in children with LCPD (Crofton et al., 2005). The RANK/RANKL/OPG pathway and some cytokines play important roles in the progress of bone remodeling in the necrotic area (Kostik et al., 2013; Melchiorre et al., 2017). These facts raise the question of whether genes involved in bone remodeling and bone structural genes could be associated too with LCPD.
This is the first study to explore the association of these genes with Mexican LCPD patients. In this investigation, a total of 23 patients with Perthes disease with a mean age of 16.87 ± 11.31 were studied and compared with a control group of 46 healthy individuals with similar ethnic backgrounds, whose ages, sex, and BMI were matched. This work analyzed variants in RANK, RANKL, OPG, IL-6, and COL1A1 genes, involved in bone remodeling and bone structure, to deepen the knowledge of the role of several genetic factors in the pathophysiology of LCPD. The variants IL-6 rs1800795 and OPG rs2073618 were associated with increased risk of LCPD (OR: 3.8 [1.08–13.54], p = 0.033 and OR: 4.34 [1.04–18.12], p = 0.033, respectively) in a sample of Mexican children.
The interruption of the blood supply to the femoral head, causing pathological and structural changes in the developing femoral head, is a key event (Crofton et al., 2005; Kim, 2010). Thrombophilic disorders have been suggested as the causal factor for LCPD and for the pathological changes in the femoral head, such as antithrombin deficiency, PC or PS deficiencies, the presence of FV Leiden or PT C20210A and elevated levels of lipoprotein A or fibrinogen, which has been investigated in children with LCPD (Vosmaer et al., 2010).
In this work, differences were found in FV, FIX, and Hcy. Vokkerla et al. (2019) found significant differences in the activity of FVIII in patients; although no significantly higher activity of FVIII was found in this work, the LCPD patients showed a trend toward higher values, implicating that the results of this work and those of Vokkerla et al. (2019) could correspond with a thrombophilic state or hypercoagulability, which has been proposed as the causal factor of LCPD. This thrombophilic and hypercoagulable state may be what initiates the cascade of events and promotes the development of bone deep vein thrombosis. Glueck et al. (2008) proposed that hypercoagulability could promote the occlusion of the epiphyseal vessels of the femoral head due to increased intraosseous pressure, causing reduced arterial flow, ischemia, and infarction triggering LCPD.
There are very few reports about the study of candidate genes for LCPD. The polymorphism rs1801133 in the MTHFR gene has been explored in several populations, with conflicting results (Azarpira et al., 2018; García-Alfaro et al., 2021; Srzentić et al., 2015; Woratanarat et al., 2014). Indeed, in the only report with Mexican patients in which this MTHFR polymorphism was analyzed, an association was obtained (Buendía-Pazarán et al., 2022). Another example of controversy in the association is the IL-6 polymorphism rs1800795; the CC genotype was associated with a high risk of LCPD in Iranian children, while in Serbian children, the CC genotype did not show an association, and the genotype with one copy of the C allele (GC) was associated with a low risk of developing LCPD (Akbarian-Bafghi et al., 2021; Srzentić et al., 2014). These facts emphasized the importance of continuing to explore the genetic factors of LCPD in Mexican population.
There are no specific risk factors for LCPD described in the Mexican population. Certain gene variants that are associated with risk in some populations may not be associated with other populations, and they have to be explored to confirm or reject their use as genetic indicators of risk in the population of interest. The varied ancestry of the Mexican population is because of the mixture of Native American genes (with Asiatic ancestry contributed by the migrations across the Bering Strait), European genes (from the Spanish conquest), and African genes (due to the arrival of African slaves). The genetic complexity of this population and the fact that association studies can provide contradictory results was the origin of the interest to search for associations of genetic variants with the risk of LCPD in this specific population to confirm the associations reported with other populations and/or to expand the research landscape of LCPD genetic factors.
The modulation of the RANK/RANKL/OPG pathway plays a key role in regulating osteoclast function, and an imbalance in the pathway can lead to pathological bone remodeling and abnormal bone remodeling of the femoral head (García-Unzueta et al., 2008; Ney et al., 2013). In this study, the SNPs RANK rs3018362 and RANKL rs12585014 did not present significant associations with LCPD. However, the OPG SNP rs2073618 was associated with an increased risk of LCPD; carriers of the G/C-G/G genotypes had four times higher risk of LCPD. The finding for the genotype of rs2073618 for LCPD in this work coincides with previous reports; it is a high-risk genotype for osteoporosis in different populations (Abdi et al., 2020; Arko et al., 2005; Kim et al., 2007; Pitocco et al., 2009).
The OPG gene is an extremely important element in the regulation of bone mass and bone metabolism. The impact of the rs2073618 SNP, located in the first exon, which encodes the OPG signal peptide, results in a change from lysine to asparagine (Lys3Asn), which may result in a qualitative alteration of OPG synthesis, compromising its function as a decoy receptor, and consequently the OPG/RANKL ratio (Choi et al., 2006). Due to its localization, this polymorphism could influence the trafficking or the secretion of OPG from the cells, since carriers of this polymorphism have lower levels of OPG in serum, which could directly affect LCPD by uncoupling the equilibrium between bone formation and bone resorption in the fragmentation phase of LCPD, similar to that which occurs in osteoporosis (Abdi et al., 2020; Kim et al., 2007; Narazaki et al., 2017).
Of course, given the coregulation and cooperation of RANK and RANKL with OPG in bone metabolism, it is also important to explore other variants of these genes in an attempt to find an association with RANK and RANKL genes with LCPD.
Inflammatory cytokines such as IL-6 can modulate skeletal homeostasis and osteoclast differentiation. One of the main functions of IL-6 is its participation in the regulation of bone metabolism, and dysregulated production of this cytokine is involved in bone pathologies such as hip osteoarthritis, osteoporosis, and inflammatory conditions such as rheumatoid arthritis; that is, IL-6 has anti-osteogenic and pro-osteoclastic effects that lead to a net inhibition of bone formation or activation of bone resorption. Kamiya et al. (2015) hypothesized that IL-6 may mediate the decoupling of bone resorption and bone formation after ischemic osteonecrosis, affecting osteoclasts positively and osteoblasts negatively.
Polymorphisms in the promoter region of the IL-6 gene, in particular the SNPs at positions G-174C (rs1800795) and G-597A (rs1800797) that are involved in transcriptional regulation of the IL-6 gene, are associated with plasma IL-6 levels and with the risk of various bone and joint diseases (Kamiya et al., 2015). In this study, the SNP C allele rs1800795 of IL-6 was associated with a significantly increased risk of LCPD. Individuals who carry the GC and CC genotypes showed between three and five times more risk of having LCPD. These results are consistent with those reported in a study with Iranian children (Akbarian-Bafghi et al., 2021) and contradict the results reported for Serbian children in which the GC genotype was associated with protection from LCPD (Srzentić et al., 2014). On the other hand, the SNP rs1800796 of IL-6 was found not to be associated with LCPD; this supports the results reported by Akbarian-Bafghi et al. (2021), who likewise did not find an association between IL-6 rs1800796 and LCPD.
IL-6 has been shown to be involved in bone metabolism and participates as a resorptive cytokine. According to these results, IL-6 rs1800795 showed a three- to fivefold increased risk of developing the disease, and it could alter bone remodeling in LCPD, moving the remodeling process balance toward bone resorption in the fragmentation stage of the disease.
There is a link between thromboembolic risk factors in LCPD and the genes involved in bone remodeling, which could be mainly due to the participation of IL-6 in both processes as a mediator of coagulation and in bone metabolism. The role as a mediator of the coagulation system includes not only an increase in fibrinogen and platelet counts but also induces activation of an extrinsic coagulation cascade. IL-6 induces the positive regulation of tissue factor (TF) on the cell surface, causing fibrin deposits mediated by thrombin; the activation of coagulation is regulated by natural inhibitors of coagulation, TF pathway inhibitor, activated PC, and antithrombin (Choi et al., 2006). On the other hand, inflammation is also modulated by components of the thrombin/fibrin pathway, the TF/FVIIa complex, the TF/FVIIa/FXa complex, FXa alone, thrombin, and fibrin, which affects inflammatory cell responses (Choi et al., 2006).
Considering the multifactorial nature of LCPD, the present data indicate a strong likelihood of the involvement of both coagulation factors and bone metabolism factors, such as IL-6, since the rs1800795 polymorphism is associated with an increased risk of LCPD with CG and CC genotypes in this and other works (Akbarian-Bafghi et al., 2021; Srzentić et al., 2014). The polymorphism rs1800795 has been associated with high levels of transcription of the IL-6 gene and with high levels of circulating IL-6 in several diseases, and the CG genotype specifically has been associated with the progression of the inflammatory response in inflammatory diseases (Oldakowska et al., 2022).
Interestingly, in coronary arterial disease, the levels of IL-6 are elevated in atherosclerotic plaques, and it is well known that thrombosis and atherosclerosis are intimately linked. It has been proposed that avascular necrosis and LCPD are associated with abnormal lipid metabolism, which is considered a clinical risk factor (Lu et al., 2020; Oldakowska et al., 2022). This study contains no information about lipids. But it would be interesting to include this variable in future works with LCPD patients since one study reported that genetic polymorphisms involved in lipid metabolism were also associated with osteonecrosis of the femoral head in Chinese patients. This was also reported in a study that mentioned that leptin could play a role in bone remodeling in ischemic necrosis of the femoral head in Korean patients, thus suggesting that abnormal lipid metabolism may play an important role in the pathology of LCPD (He and Li, 2009; Lee et al., 2013; Liu et al., 2015).
Type I collagen is the main component of the extracellular matrix; the COL1A1 gene codes for the alpha 1 chains of the collagen protein, which is formed by two alpha 1 chains and one alpha 2 chain. The Sp1 polymorphism (rs1800012) of COL1A1 affects the binding site of the transcription factor Sp1, resulting in the synthesis of homotrimers with 3 alpha 1 chains that show instability compared to a heterotrimer, causing a disorganized narrowing of type I collagen and reduced bone mineralization (Erdem et al., 2019; Posthumus et al., 2009). This polymorphism has been associated with bone metabolism, causing a decrease in bone mineral density, osteoporosis, and rheumatoid arthritis; whereas on the other hand, it could also be involved in bone injuries such as osteoarthritis (Luo et al., 2016). In the context of these studies, it is proposed that rs1800012 could be associated with LCPD.
In this study, two polymorphisms in the COL1A1 gene were explored; however, neither rs1800012 nor rs2586498 presented significant associations with LCPD due to the low frequency of the A allele and the absence of the AA genotype of both polymorphisms. Exploration of other polymorphisms in this structural gene should be considered before ruling out its link with LCPD.
Since it is a rare disease, the sample size of patients properly diagnosed with LCPD in the Mexican population is quite small. The relevance of this investigation lies in the encouraging results about genetic factors, specifically about the contribution of bone remodeling genes to the risk of LCPD. This is the first study to analyze polymorphisms of several bone metabolism genes as candidate genes and to simultaneously include all of these polymorphisms in the RANK, RANKL, OPG, IL-6, and COL1A1 genes, providing new and important insights into the genetic factors involved in LCPD.
These results could lead to the hypothesis that there is a connection between thrombotic events and bone metabolism imbalance, since IL-6 is involved in both mechanisms, and it is possible that IL-6 along with OPG could modify the bone turnover equilibrium simultaneously in several phases of the disease. These findings exploring genetic, coagulation, and biochemical markers involved in the pathogenesis confirm that LCPD is a multifactorial and multigenic disease.
The results of this investigation allow us to obtain a broader understanding of the possible mechanisms that activate the process of necrosis in the femoral head. The determination of these pathways would be relevant for the early detection of LCPD and to develop possible biomarkers for better diagnosis of the disease that could be applied in early stages in children diagnosed with LCPD, thus preventing collapse of the femoral head.
Conclusions
The aforementioned results of this study suggest that IL-6 and OPG have important roles in LCPD and showed that the IL-6 rs1800975 and OPG rs2037618 polymorphisms are informative markers of increased risk of LCPD in Mexican children. However, further studies are required to analyze the effects, if they indeed exist, of IL-6 rs1800795 and OPG rs2037618 polymorphisms directly on LCPD processes.
Likewise, some of the analyzed polymorphisms did not show significant association with susceptibility to LCPD in Mexican children; however, other polymorphisms should be explored before discarding their participation of these genes in the disease.
Footnotes
Acknowledgments
The authors would like to thank Dr. Margarita Valdés Flores for her support and her contributions to the conceptualization and realization of this work. Your working group will always remember you. Rest in the everlasting peace.
Equally, the authors would like to thank all patients who kindly participated in the study.
Authors' Contributions
All authors agree to publish this article. L.C.A.: study conceptualization and project administration. B.L.C.O.: investigation. L.C.A., J.R.L., E.R.M.: resources. B.L.C.O., L.C.A.: formal analysis. B.L.C.O., L.C.A.: writing – review and editing. E.H.Z., E.R.M., C.O.M.R., C.C.H., A.O.R.O.: writing. B.L.C.O., R.L.P.: visualization.
Authors Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.