
Abstract
Patients with colorectal cancer (CRC) and diabetes share many risk factors. Despite a strong association between diabetes and CRC being widely studied and confirmed, further genetic research is needed. This study found higher AL049796.1 and TEA domain transcription factor 1 (TEAD1) levels (both mRNA and protein) in CRC tissues of diabetic patients compared with nondiabetics, but no significant difference in miR-200b-3p levels. A positive correlation between AL049796.1 and TEAD1 protein existed regardless of diabetes status, whereas miR-200b-3p was only negatively correlated with TEAD1 protein in nondiabetic CRC tissues. In vitro experiments have shown that high glucose (HG) treatment increased AL049796.1 in CRC cells, and AL049796.1 silencing reduced HG-induced proliferation, migration and invasion, as well as connective tissue growth factor, cysteine-rich angiogenic inducer 61, and epidermal growth factor receptor protein expression. Mechanistic investigations indicated that AL049796.1 could mitigate suppression of miR-200b-3p on TEAD1 posttranscriptionally by acting as a competitive binder. In vivo, subcutaneous CRC tumors in streptozotocin (STZ)-induced mice grew significantly faster; AL049796.1 silencing did not affect the growth of subcutaneous CRC tumors but significantly reduced that of STZ-induced mice. Our study suggests that AL049796.1 independently contributes to the risk of CRC in diabetic patients, highlighting its potential as both a therapeutic target and a novel biomarker for CRC among individuals with diabetes.
Introduction
Epidemiological studies underscore a direct linkage between diabetes and increased cancer incidence and mortality, positioning cancer as a potential diabetes complication (Buysschaert and Sadikot, 2013; Yang et al., 2017). Recent findings from the German Cancer Research Center (DKFZ) indicate that diabetes patients face a heightened risk of colorectal cancer (CRC) onset 5 years earlier compared with nondiabetics among those aged 50 years and older (Ali Khan et al., 2020a; Ali Khan et al., 2020b). It was also observed that there is a significantly greater prevalence of colon adenomas and advanced adenomas in diabetic patients. Furthermore, type 2 diabetes mellitus (T2DM) was found to affect adverse drug reactions, even when accounting for multiple confounders (Ottaviano et al., 2020). Therefore, early interventions to prevent T2DM and the prescription of antidiabetic medication could potentially reduce the risk of CRC. Current research indicates that hyperglycemia in diabetic patients plays a significant role in driving tumor progression and regulating metastasis in CRC. For instance, Ran et al.’s study highlights that stearoyl-CoA desaturase1 enhances CRC metastasis through monounsaturated fatty acid production and transforming growth factor-β1-regulated and epithelial cell-enriched phosphatase suppression in response to glucose (Ran et al., 2018). Furthermore, it has been proven that diabetes promotes tumor cell proliferation by inhibiting the adenylate-activated protein kinase activity (Han et al., 2013). These findings collectively underscore diabetes as a significant risk factor for CRC, emphasizing the need for mechanistic studies to support the development of advanced therapeutic strategies for treating CRC in diabetic patients.
In this study, we present the first evidence of a long noncoding RNA (lncRNAs), AL049796.1, playing a key role in the modulation of high glucose (HG)-induced CRC progression through the miR-200b-3p/TEA domain transcription factor 1 (TEAD1) axis. AL049796.1, situated on human chromosome 1 (94312730–94314495), emerged as one of the most upregulated lncRNAs in CRC tissues from diabetic patients, identified by our team. To date, no studies have explored the biological function of AL049796.1 in relation to either CRC or diabetes. Our primary objective is to elucidate the mechanism of AL049796.1 and to preliminarily evaluate the efficacy of silencing AL049796.1 in arresting CRC progression under HG conditions.
Methods
Patients and sample collection
This study involved 48 CRC tissue samples collected from patients with and without T2DM, who underwent tumor resection at the Harbin Medical University Cancer Hospital from January 2018 to December 2021. All patients signed the written informed consent. The study was approved by the Ethics Committee of the Harbin Medical University cancer hospital (Approval. No. HMUIRB0222021). Procedures operated in this research were completed in accordance with the standards set out in the Announcement of Helsinki and laboratory guidelines of research in China. Detailed exclusion and inclusion criteria, clinicopathological information, and tumor classification are outlined in Supplementary Table S1. Tissue samples were harvested at least 2 cm from the tumor margin, measuring approximately 50 mm3, and were immediately snap-frozen in liquid nitrogen postsurgery for preservation at −80°C until analysis. Total RNA extracted from these CRC tissues was analyzed to determine the expression levels of AL049796.1, miR-200b-3p, and TEAD1 mRNA through real-time quantitative PCR (RT-qPCR). Concurrently, total protein was extracted for TEAD1 protein expression analysis using enzyme-linked immunosorbent assay (ELISA), following the instructions provided by the Human TEAD1 ELISA Kit (CUSABIO, Wuhan, China). Pairwise correlation analyses between AL049796.1, TEAD1, and miR-200b-3p expressions were then conducted for each group of CRC tissues.
Cell culture methods
The CRC cell lines, Caco-2 and HCT-116, were acquired from the American Type Culture Collection (ATCC, Maryland, USA) and cultured in Dulbecco’s modified Eagle’s medium (DMEM, Thermo Fisher Scientific Inc., CA, USA) enriched with 10% fetal bovine serum (FBS, Thermo Fisher Scientific Inc.). These cells were maintained in an adherent culture, with subculturing performed through 0.25% trypsin digestion upon reaching 80% confluence. All cells were incubated in a humidified environment at 37°C, 5% CO2, and 95% humidity.
Lentivirus-mediated TEAD1 overexpression and AL049796.1 silencing in CRC cells
We prepared recombinant lentiviruses (Lv-shRNA-AL049796.1, Lv-TEAD1, and Lv-NC) according to the protocol provided by the pPACK Packaging Plasmid Mix Reagent Kit (System Biosciences, CA, USA). Lv-shRNA-AL049796.1 contains an HI promoter for the production of siRNA-AL049796.1 (5′-GUAAACAUCCAUGUUAUUC-3′) selected by using the online siRNA design software “siRNA Finder” (Thermo Fisher Scientific Inc.); the coding sequence of the human TEAD1 gene (NM_021961.6) was cloned into Lv-TEAD1 under a cytomegalovirus promoter, and Lv-NC was used to transcript a scrambled sequence of siRNA (5′-GAAGCCAGAUCCAGCUUCC-3′), which was used as a negative control (NC) to overcome the off-target effects. The Caco-2 and HCT-116 cells in logarithmic growth phase were digested and reseeded into a 6-well culture plate (5 × 104 cells/well) followed by overnight culture. Subsequently, the recombinant lentiviruses (Lv-NC, Lv-shRNA-AL049796.1, or Lv-TEAD1) were added into the cell supernatants with a multiplicity of infection (MOI) of 10 followed by culturing under normal conditions. At 72 h after infection, the infection efficiency of lentivirus on Caco-2 and HCT-116 cells was analyzed by observing the expression of green fluorescent protein (GFP) under a fluorescence inverted microscope (IX72, Olympus, Japan). Furthermore, the infected cells were used for detecting the expression levels of AL049796.1 and TEAD1 mRNA by RT-qPCR. We also extracted the total proteins from these cells, which were then used to detect TEAD1 protein expression using western blotting.
Luciferase assay
TargetScan7.1 (Whitehead Institute for Biomedical Research, MI, USA) was used to predict potential binding sites between miR-200b-3p and 3′-untranslated region (UTR) of TEAD1 mRNA. After that, we synthesized DNA sequences complementary to 3′-UTR of TEAD1 5′-TTCTCACACATGTTTTCTTAACCTATTTGCAGAAACTTTCAAAAGGCATTTGATTAAACCTCTTGGCAGTACAGTATTCTTGTATTTGTTAACGTCTGTGTTTAGGTACTGGTACCTT-3′, which was used to construct pGl3-wt-TEAD1, containing a wild-type binding site of miR-200b-3p. The binding site in pGL3-wt-TEAD1 was then mutated from 5′-CAGTATT-3′ to 5′-TAGTATC-3′ to construct pGL3-mt-TEAD1. HEK293 cells were cotransfected with miR-200b-3p mimics (5′-UAAUACUGCCUGGUAAUGAUGAtt-3′)/inhibitors (5′-UCAUCAUUACCAGGCAGUAUUAtt-3′)/NC (5′-UAAUACUGCCUGGUAAUGAUGAtt-3′) (Gene Pharma, Shanghai, China) and pGL-wt-TEAD1 or pGL3-mt-TEAD1 using the X-tremeGENE 360 transfection reagent (Roch, Basilea, Switzerland). Luciferase activity was measured 48 h posttransfection using the Dual-Luciferase Assay Kit (Beyotime, Jiangsu, China). In addition, we also verified the intervention of AL049796.1 on the binding of endogenous miR-200b-3p on TEAD1 mRNA in CRC cells through the luciferase assay. Briefly, we first transfected pGL3-wt-TEAD1 into HCT-116 cells, and then the Lv-shRNA-AL049796.1 or Lv-AL049796.1, respectively. At 48 h after transfection, the luciferase activity in cells was evaluated.
RNA-binding protein immunoprecipitation assay
TEAD1 protein in Caco-2 cells was immunoprecipitated using a TEAD1-specific antibody (ab133533, Abcam, Cambridge, UK). The RNA bound to the TEAD1 protein served as a template for amplifying a segment of AL049796.1 through the SuperScript IV UniPrime One-Step RT-PCR System (Thermo Fisher Scientific). We confirmed the desired PCR products by analyzing 5 µL of the product on a 2% agarose gel. The PCR primer sequences are listed in Supplementary Table S2.
Cell proliferation, invasion, and migration assays
Caco-2 and HCT-116 cells, after 72-h infection with lentiviruses (Lv-NC or Lv-shRNA AL049796.1), were reseeded in a 96-well culture plate (3 × 104 cells/well) and cultured for an additional 72 h in DMEM supplemented with 35 mmol/L glucose and 10% FBS. Subsequently, cell proliferation was assessed using the Cell Counting Kit-8 Assay (CCK-8, Dojindo, Japan). The absorbance at 450 nm was measured using a microplate reader (Multiskan SkyHigh, Thermo Fisher Scientific Inc), reflecting cell proliferation activity through a reliable linear relationship. The invasion assay was conducted following the QCM-Fluorimetric Cell Invasion Assay Kit (ECM554, Millipore, MI, USA) protocol. Briefly, cells were placed in the upper chamber with 500 µL of serum-free DMEM (4.0 × 104 cells each), whereas the lower chamber was filled with 750 µL of DMEM supplemented with 35 mmol/L glucose and 10% FBS. After incubation at 37°C and under 5% CO2 for 24 h, cells that penetrated the membrane were fixed with methanol, stained with Hoechst33342 (C1022, Beyotime), and counted in five random fields using a light microscope. Simultaneously, cell migration was assessed using a wound healing assay. Briefly, a 200-μL pipette tip was used to scratch the cell monolayer, creating wounds. The cells were then washed twice with Dulbecco’s phosphate-buffered saline (pH = 7.4) to remove detached cells. Plates were returned to the incubator for 24 h and 72 h before the wounds were examined using a phase-contrast light microscope.
Investigating the impact of AL049796.1 silencing on TEAD1 expression and downstream genes in CRC cells under HG conditions
To investigate the impact of HG conditions on AL049796.1 expression in CRC cells, we seeded logarithmic-phase Caco-2 cells in a 6-well plate at 1 × 105cells/well with DMEM containing 10% FBS. After overnight culture, the medium was replaced with DMEM containing 35 mmol/L glucose and 10% FBS. Cells were harvested after 24 and 48 h of culture to assess AL049796.1 and miR-200b-3p levels using RT-qPCR. Meanwhile, HCT-116 cells, infected for 72 h with recombinant lentiviruses (Lv-NC, Lv-shRNA-AL049796.1, or Lv-shRNA-AL049796.1+Lv-TEAD1), were reseeded in a 6-well plate at 1 × 105 cells/well in DMEM supplemented with 35 mmol/L glucose and 10% FBS. After 48 h of culture, cells were harvested and used to detect the TEAD1 protein expression, as well as its downstream regulatory proteins, connective tissue growth factor (CTGF), cysteine-rich angiogenic inducer 61 (Cyr61), and epidermal growth factor receptor (EGFR), by western blotting. To control for osmotic effects, 35 mmol/L mannitol was used, and Lv-TEAD1 served as a pathway control.
Subcutaneous tumor-bearing experiment
Experimentation on animals was implemented by strictly following the National Institutes of Health Guide for the Care and Use of Laboratory Animals. The animal experiments were approved by the Ethics Committee of Harbin Medical University cancer hospital (Approval. No. HMUIRB0222021). Forty male BALB/c nude mice, 6 weeks old and weighing approximately 18 ± 2 g, were acquired from SLAC Laboratory Animal Co. Ltd. (Shanghai, China). Initially, half of the mice were fed a high-energy diet comprising 58% fat, 25% protein, and 17% carbohydrates for 4 weeks followed by an intraperitoneal low-dose streptozotocin (STZ) injection over 5 days to establish a T2DM model referring to published reports (Al-Awar et al., 2016; Furman, 2021). To prepare the injection solution, 2.10 g of citric acid was dissolved in 100 mL of double-distilled water, creating a citric acid stock solution (Solution A); similarly, 2.94 g of trisodium citrate was dissolved in 100 mL of double-distilled water to form a sodium citrate stock solution (Solution B). Solutions A and B were mixed in a 1:1.32 ratio, and the pH was adjusted to 4.0 with a pH meter, resulting in the desired 0.1 M sodium citrate buffer for STZ preparation. STZ was then dissolved in this buffer to create a fresh 10 mg/mL STZ solution, which was sterilized through a 0.22-μm filter in a light-protected environment and used immediately. Mice were fasted for 12 h before the procedure. The induction group received an intraperitoneal injection of STZ at a dose of 50 mg/kg once a day for 5 consecutive days, whereas the control group was administered an equivalent volume of sodium citrate buffer. One week after the final STZ injection, fasting blood glucose levels were measured. Mice with blood glucose levels exceeding 11.1 mmol/L were selected for further experiments.
HCT-116 cells (1 × 107) suspended in 50 μL of FBS were subcutaneously injected into the flank regions of the mice (12 mice were selected randomly from the normal and STZ-induced groups). When tumors reached approximately 2 mm in diameter, mice were randomly allocated into four groups (n = 6 each): control group, STZ group, Lv-shRNA-AL049796.1 group, and Lv-shRNA-AL049796.1 +STZ group. For Lv-shRNA-AL049796.1 injection, each mouse received weekly injections of Lv-shRNA-AL049796.1 (1 × 107 ifu) for 6 consecutive weeks, whereas the control group received an equivalent volume of saline. Tumor diameters were measured weekly to calculate tumor volume. At the intervention endpoint, blood was collected from the tails of the mice to measure random blood glucose levels, and subcutaneous tumors were excised and cryopreserved in liquid nitrogen for subsequent analysis of AL049796.1 and TEAD1 protein expression levels.
RNA isolation and RT-qPCR
The CRC cells and tissues were fully lysed in TRIzol reagent (Thermo Fisher Scientific Inc) and used to isolate total RNA. Two micrograms of total RNA were reverse transcribed into cDNA by using the QuantiTect Rev. Transcription Kit (Qiagen, Germany). A specific primer was used for the reverse transcription of miR-200b-3p, as listed in Supplementary Table S2. Then the quantitative PCR was conducted using a GoTaq qPCR Master Mix (Promega) and the Applied Biosystems7500 Fast (Applied Biosystems/Thermo Fisher) with three repeated reactions with the primers listed in Supplementary Table S2. The differences between groups in expression levels of AL049796.1, miR-200b-3p, and TEAD1 mRNA were analyzed using the 2-ΔCt method, and U6 or β-actin was used as internal reference.
Western blotting
The CRC cells and tissues were collected and lysed for 15 min on ice in lysis buffer containing protease and phosphatase inhibitors. The lysates were centrifuged at 12,000 ×g and 4°C for 10 min. Sodium dodecyl sulfate (SDS)-loading buffer was added to the supernatant, and then the sample was heated at 95°C for 5 min before loading on the SDS polyacrylamide gel. After centrifugation, 15 μg protein of each sample were equally loaded on 11% SDS polyacrylamide gels and transferred onto nitrocellulose membranes, which were blocked with bovine serum albumin (5%) at 4°C for 2 h. The blots were probed with primary antibodies against human TEAD1 (1:500), CTGF (1:500), Cyr61 (1:600), EGFR (1:500), and β-actin (1:2000) (Abcam), followed by probing with HRP-conjugated goat anti-rabbit IgG (1:3500, Abcam). After washing the membranes twice with Tris-buffered saline and Tween 20 (TBST. pH = 7.4) and visualized by enhanced chemiluminescence, following incubation for 2 min, membranes were exposed to X-ray films. The protein bands were scanned and analyzed. The relative expression levels of the target proteins were calculated as ratio of optical density to β-actin by using ImageJ 2x software (National Institutes of Health, Maryland, USA).
Statistical analysis
All data are expressed as the means ± SD of at least three independent experiments and analyzed using the GraphPad Prism 8.0 software package and SPSS software (Version 15.0). One-way analysis of variance with the Tukey–Kramer post hoc multiple comparisons test were used to identify significant differences among groups. *p < 0.05, **p < 0.01.
Results
Expression analysis of AL049796.1, miR-200b-3p, and TEAD1 in CRC tissues from patients with and without diabetes
Analysis through RT-qPCR revealed that the expression levels of AL049796.1 and TEAD1 mRNA were significantly elevated in the tumor tissues of CRC patients with diabetes, compared with those without diabetes (p < 0.01, vs. CRC patients without diabetes). Conversely, the expression levels of miR-200b-3p did not show a significant difference between the two groups (p > 0.05, vs. CRC patients without diabetes) (Fig. 1A). Further examination through ELISA demonstrated that TEAD1 protein levels were notably higher in the tumor tissues of CRC patients with diabetes when compared with those without diabetes (p < 0.01) versus CRC patients without diabetes (Fig. 1B). In addition, the findings indicated that the rate of change in TEAD1 mRNA levels was considerably less pronounced than that observed in its protein levels.

Analysis of AL049796.1, miR-200b-3p, and TEAD1 expression in tumor tissues from CRC patients with and without diabetes.
Exploring pairwise correlations in the expression of AL049796.1, miR-200b-3p, and TEAD1 in CRC tissues from patients with and without diabetes
A detailed analysis indicated a positive correlation between AL049796.1 and TEAD1 protein expression in the tumor tissues of CRC patients, regardless of their diabetes status. In contrast, a negative correlation between miR-200b-3p and TEAD1 protein was observed exclusively in tumor tissues from CRC patients without diabetes. In addition, no correlation was found between the levels of miR-200b-3p and TEAD1 mRNA in the tumor tissues of CRC patients, whether they had diabetes or not (Fig. 2).
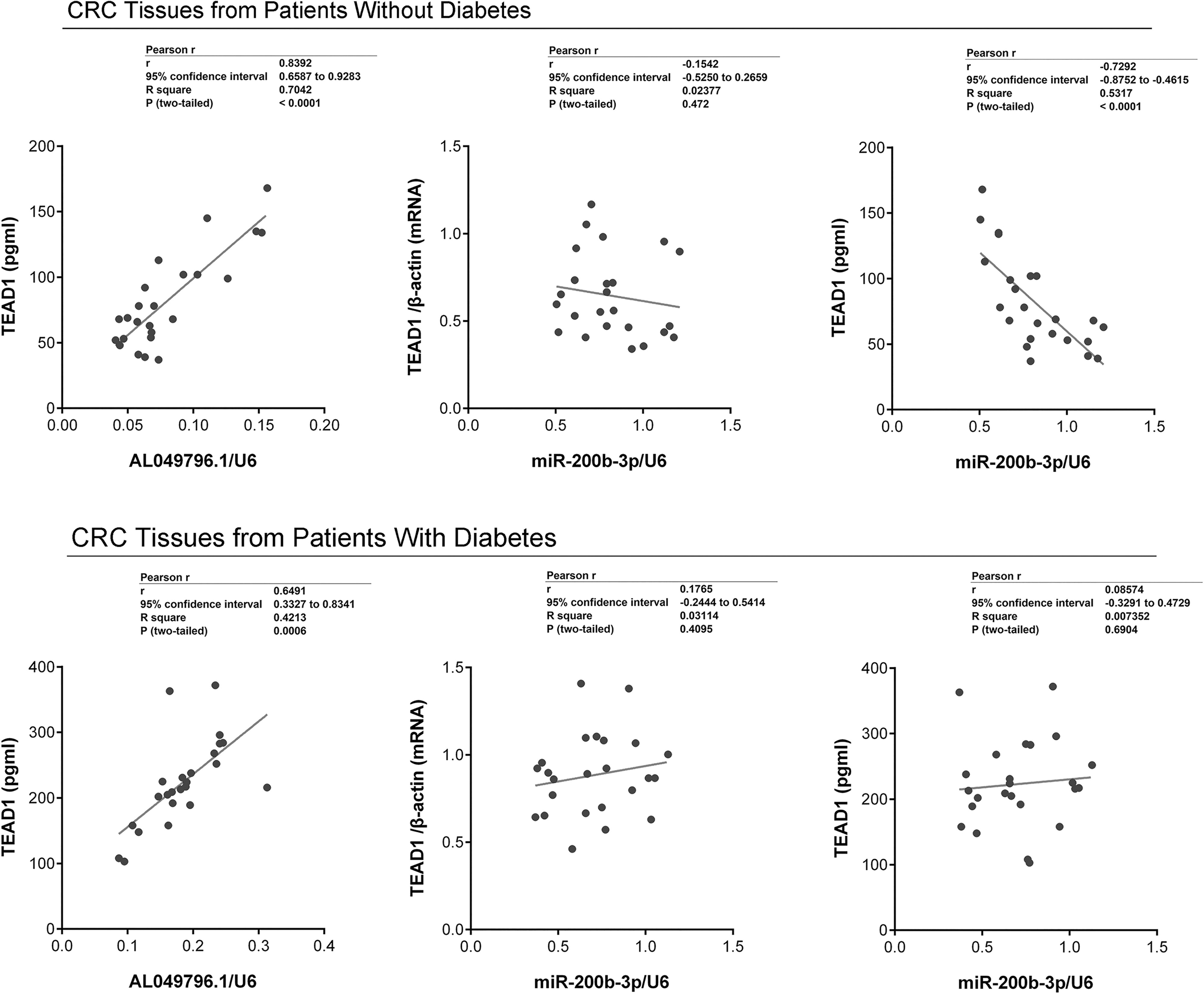
Cross-correlation analysis of AL049796.1, miR-200b-3p, and TEAD1 expression in tumor tissues of CRC with and without diabetes. Performed using Spearman’s correlation analysis. Data are summarized as mean ± SD (n = 24).
Effective silencing of AL049796.1 and overexpression of TEAD1 in CRC cells through lentiviral delivery
At 72 h postinfection, a high prevalence of GFP signals was observed in the majority of CRC cells under fluorescent microscopy, indicating an infection efficiency of the recombinant lentivirus in Caco-2 and HCT-116 cells close to 100% (Fig. 3A). RT-qPCR analysis demonstrated a significant reduction in AL049796.1 expression levels in the Caco-2 and HCT-116 cells treated with Lv-shRNA-AL049796.1 (p < 0.01, vs. control group), whereas no significant expression differences were noted between the control, Lv-NC, and Lv-TEAD1 groups (p > 0.05). Conversely, TEAD1 mRNA levels increased significantly in the Caco-2 and HCT-116 cells of the Lv-TEAD1 group (p < 0.01, vs. control group), with no notable differences observed among the control, Lv-NC, and Lv-shRNA-AL049796.1 groups (p > 0.05) (Fig. 3B). Western blot analysis revealed that, relative to the control group, TEAD1 protein expression was significantly reduced in CRC cells of the Lv-shRNA-AL049796.1 group and significantly increased in those of the Lv-TEAD1 group (p < 0.01, vs. control group) (Fig. 3C). These findings suggest that silencing AL049796.1 can downregulate TEAD1 protein expression (but not its mRNA levels), whereas TEAD1 overexpression does not significantly affect AL049796.1 levels in CRC cells, implying that AL049796.1 acts upstream of TEAD1.
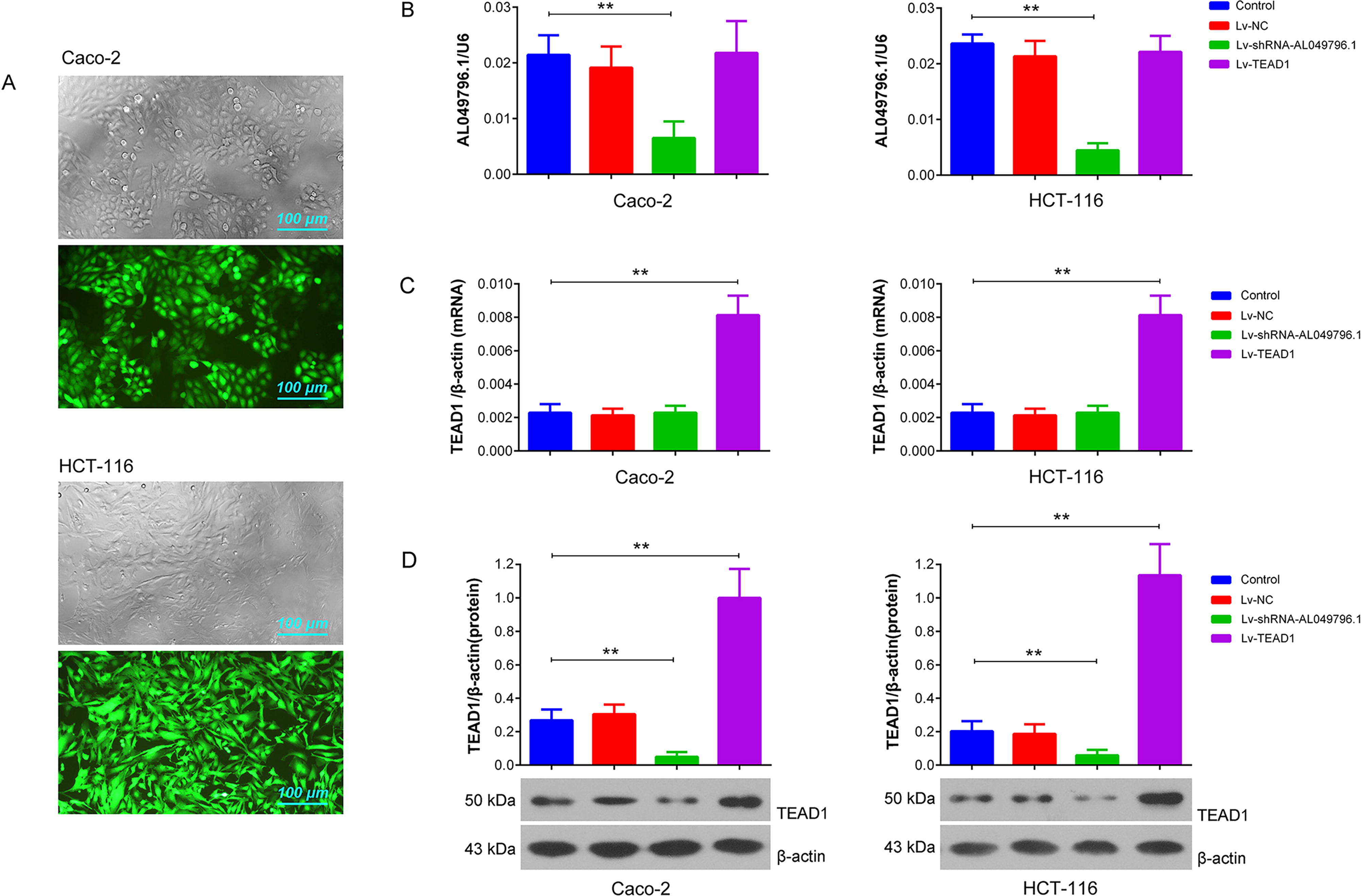
Lentivirus-mediated AL049796.1 silencing and TEAD1 overexpression in Caco-2 and HCT-116 cells.
MiR-200b-3p interaction with the 3′-UTR of TEAD1 mRNA
Bioinformatics analysis identified a conserved 7-base sequence, 5′-CAGUAUU-3′, in the 3′-UTR of TEAD1 mRNA that serves as a binding site for miR-200b-3p (Fig. 4A). Luciferase assay results demonstrated significant upregulation of luciferase activity in HEK293 cells transfected with either pGL3-wt-TEAD1 or pGL3-mt-TEAD1 (p < 0.01, vs. control group). However, miR-200b-3p mimics significantly reduced the luciferase activity in cells transfected with pGL3-wt-TEAD1 (p < 0.01, vs. pGL3-wt-TEAD1 group), whereas the miR-200b-3p inhibitor increased luciferase activity in these cells (p < 0.01, vs. pGL3-wt-TEAD1 group). In contrast, the transfection of miR-200b-3p mimics or inhibitor did not significantly alter luciferase activity in HEK293 cells transfected with pGL3-mt-TEAD1 (p > 0.05, vs. pGL3-mt-TEAD1 group) (Fig. 4B). This suggests a specific regulatory interaction between miR-200b-3p and the wild-type 3′-UTR of TEAD1 mRNA.

Luciferase and RIP assays.
AL049796.1 inhibits miR-200b-3p binding to TEAD1 3′UTR
In HCT-116 cells, silencing of AL049796.1 led to a significant decrease in luciferase activity in cells transfected with pGL3-wt-TEAD1 (p < 0.01, vs. pGL3-wt-TEAD1 group), whereas the overexpression of AL049796.1 significantly increased this activity (p < 0.05 vs. pGL3-wt-TEAD1 group) (Fig. 4C). These results suggest that the modulation of TEAD1 protein expression by AL0v49796.1 might be through its role as a competitive endogenous RNA (ceRNA), inhibiting miR-200b-3p’s interaction with TEAD1 mRNA in these CRC cells. In addition, RNA-binding protein immunoprecipitation (RIP)-PCR analysis confirmed that AL049796.1 does not directly bind to the TEAD1 protein in HCT-116 cells (Fig. 4D), further supporting the ceRNA mechanism hypothesis.
Silencing AL049796.1 significantly reduces HG-induced proliferation, migration, and invasion in CRC cells
Proliferation assays revealed that HG culture markedly increased proliferative activity in Caco-2 and HCT-116 cells compared with the control group (p < 0.05 at 48 h, p < 0.01 at 72 h). Silencing AL049796.1 notably curtailed the HG-induced proliferation in these cells (p < 0.01, vs. HG group at 72 h) (Fig. 5A). Wound healing assays demonstrated that 48 h of HG culture significantly enhanced migration capabilities in Caco-2 and HCT-116 cells compared with the control group (p < 0.01), and the above effect was significantly mitigated by AL049796.1 silencing (p < 0.01, vs. HG group) (Fig. 5B). Transwell assays confirmed that the invasion capabilities of Caco-2 and HCT-116 cells mirrored the trends observed in their migration abilities (Fig. 5C). These findings collectively suggest that the targeted knockdown of AL049796.1 can effectively counteract the HG-induced advancement of CRC, highlighting its potential as a therapeutic target for controlling disease progression.
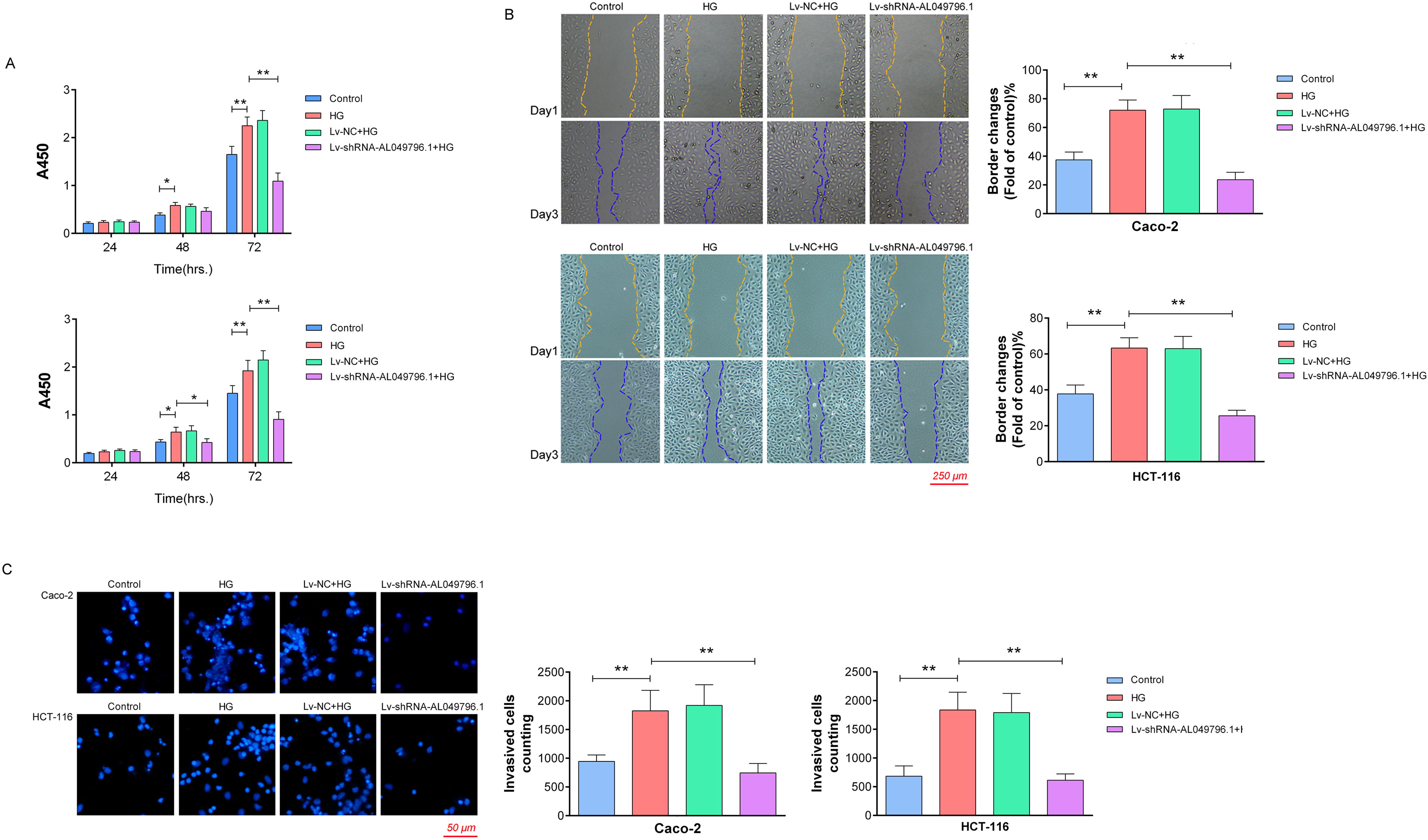
Assessment of proliferation, invasion, and migration in CRC cells under various treatment conditions.
Silencing AL049796.1 reverses the disruption of the miR-200b-3p/TEAD1 pathway by HG in CRC cells
RT-qPCR analyses indicated that AL049796.1 expression levels were significantly elevated in Caco-2 cells exposed to HG conditions (p < 0.01, vs. control group), with a notable increase observed at 48 h compared with 24 h (p < 0.01). Although miR-200b-3p levels also rose significantly after 48 h of HG treatment (p < 0.05, vs. control group), there was no significant change between the 24 and 48 h HG treatment intervals (p > 0.05) (Fig. 6A). Western blot analysis revealed that 48 h HG treatment significantly upregulated TEAD1 protein expression in Caco-2 cells (p < 0.01, vs. control group). Crucially, AL049796.1 silencing markedly reduced the HG-induced increase in TEAD1 protein (p < 0.01, vs. HG group), and the effects were counteracted by the overexpression of exogenous TEAD1 (p < 0.01, vs. Lv-shRNA-AL049796.1+HG group). Moreover, changes in the protein expressions of CTGF, Cyr61, and EGFR between groups mirrored those observed for TEAD1 protein expression (Fig. 6B). These findings suggest that HG enhances TEAD1 oncogene expression and silencing AL049796.1 can effectively mitigate the HG-induced elevation of TEAD1 by reinstating the inhibitory influence of miR-200b-3p on TEAD1 protein expression, highlighting a pivotal therapeutic avenue for modulating disease progression.
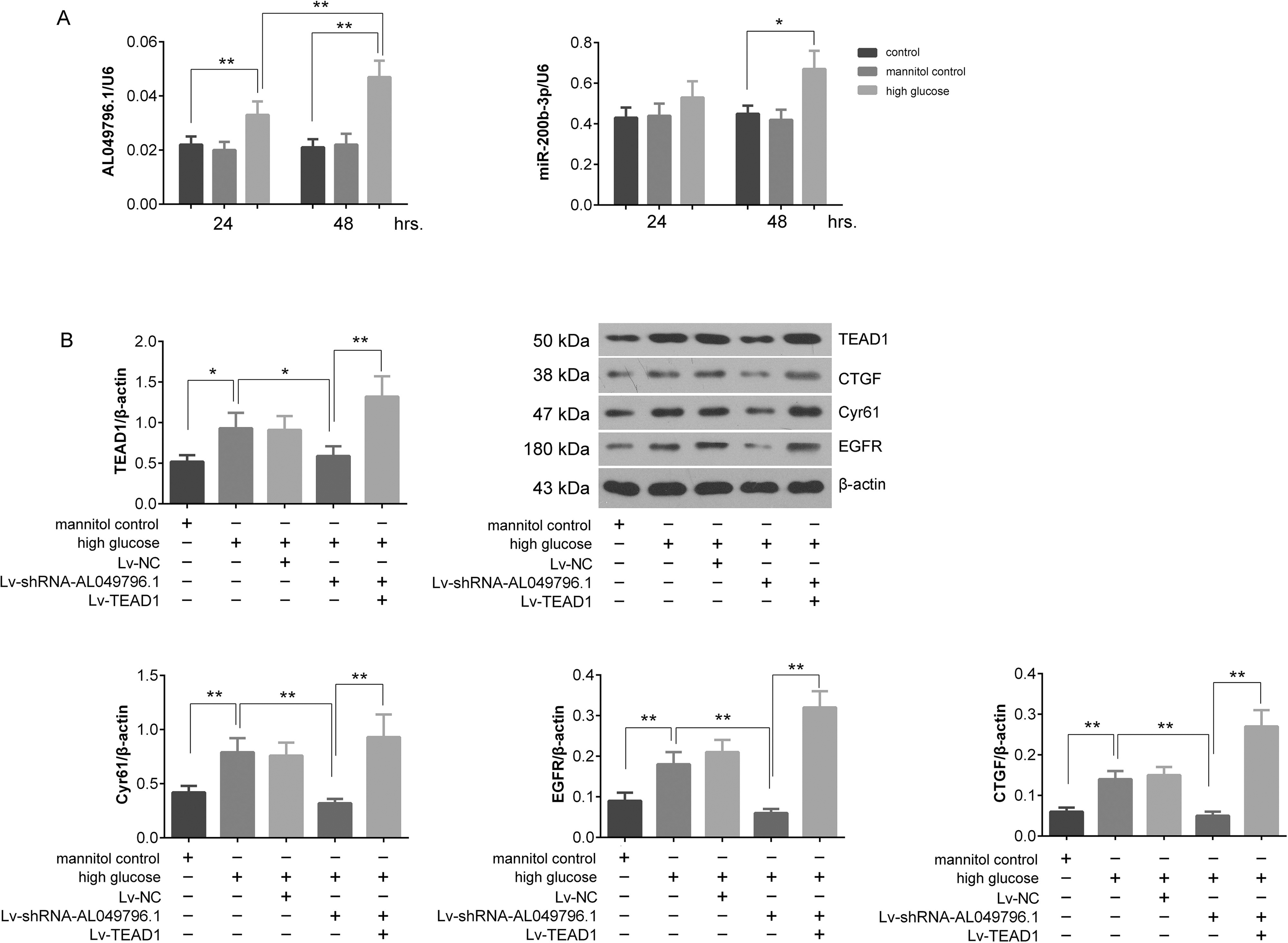
Detection of the expression of AL049796.1, miR-200b-3p, and TEAD1-related proteins in Caco-2 cells with different treatments.
AL049796.1 silencing mitigates hyperglycemia-induced subcutaneous CRC tumor growth in xenografts
Analysis of subcutaneous CRC tumor growth curves revealed a significant acceleration in tumor growth in the STZ group when compared with the control group (p < 0.01), injection of Lv-shRNA-AL049796.1 did not have a significant effect on tumor growth compared with the control group (p > 0.05). However, mice of Lv-shRNA-AL049796.1+STZ group experienced a notable deceleration in tumor growth rates when compared with the STZ group (p < 0.01) (Fig. 7A, B). Blood glucose assessments after 6 weeks of STZ induction demonstrated that random blood glucose levels exceeded 16.7 mmol/L across the STZ group and the Lv-shRNA-AL049796.1+STZ group, with no significant intergroup differences (p > 0.05). However, these levels were significantly higher than those observed in the control and Lv-shRNA-AL049796.1 groups (p < 0.05) (Fig. 7C). RT-qPCR analysis indicated a significant increase in AL049796.1 levels within the subcutaneous tumor tissues of the STZ group compared with the control group (p < 0.05), and it was significantly decreased in the Lv-shRNA-AL049796.1 group or Lv-shRNA-AL049796.1 +STZ group (p < 0.01) (Fig. 7D). Western blotting revealed a significant increase in TEAD1 protein expression in the subcutaneous tumor tissues of the STZ group compared with the control group (p < 0.05). Remarkably, TEAD1 protein expression significantly decreased in the Lv-shRNA-AL049796.1 group and Lv-shRNA-AL049796.1+STZ group compared with STZ group (p < 0.01) (Fig. 7E). These findings suggest that hyperglycemia can elevate the expression levels of AL049796.1 and TEAD1 protein in CRC tissues, thereby enhancing tumor growth. Although silencing AL049796.1 alone has no significant effect on the growth of subcutaneous CRC tumors, it can effectively inhibit tumor growth under STZ induction conditions. In addition, silencing AL049796.1 has no significant impact on blood glucose levels in STZ-induced mice (p > 0.05, vs. control group), but effectively inhibits TEAD1 protein expression (p < 0.01, vs. control group), aligning perfectly with the conclusions from in vitro mechanistic studies.

Subcutaneous tumor-bearing experiment in mice.
Discussion
Diabetes, particularly T2DM, currently poses a significant health threat globally as a chronic disease. It is the more prevalent form of diabetes, distinct from type 1 diabetes. Numerous epidemiological and clinical investigations have established a close link between T2DM and an increased risk for several types of cancer, including liver, pancreatic, endometrial, breast cancers, and CRC (Grote et al., 2010; Tsilidis et al., 2015). CRC ranks as the third most prevalent and the second deadliest cancer worldwide. A growing body of research confirms that diabetic individuals face a higher risk of developing CRC, with their treatment outcomes and prognosis generally being poorer compared with nondiabetic CRC patients. Shared risk factors, such as central obesity, a sedentary lifestyle, and a diet high in refined carbohydrates but low in fiber, underscore the critical interplay between diabetes and CRC (Luo et al., 2012). A comprehensive meta-analysis spanning 17 studies identified T2DM as a significant risk factor for colorectal adenomas and advanced adenomas, suggesting that enhanced adenoma screening in diabetic patients could serve as a preventive measure against CRC (Yu et al., 2016). Further studies reinforce the heightened risk of CRC in diabetic patients, with a particular study indicating that diabetes patients experience the highest risk within 4–8 years of diagnosis (Flood et al., 2010; Larsson et al., 2005). A meta-analysis involving 24 studies quantified this increased risk, presenting a relative risk of 1.27 (95% confidence interval [CI]: 1.14–1.42), with a notably stronger correlation in men. Collectively, these findings highlight the intricate association between diabetes and CRC. In this study, we established a T2DM nude mouse model through a high-fat diet combined with STZ induction, followed by a tumor-bearing experiment with CRC HCT-116 cells. Our data monitoring of tumor growth showed that compared to the control group, the growth of subcutaneous CRC tumors in the T2DM mice was significantly accelerated, which is completely consistent with the conclusions of the aforementioned literatures.
While numerous studies have established a connection between diabetes and CRC, the precise mechanisms underlying this association remain unresolved. Recent collaborative research by the DKFZ and other prestigious institutions, analyzing clinical data from over 12.61 million patients, revealed that diabetic individuals reach the CRC screening risk threshold approximately 5 years earlier than the general population, with this period being even shorter, 12–21 years earlier, for those with a family history of CRC (Ali Khan et al., 2020a). Research by Lorenzo F. Ottaviano et al. indicated a significantly higher incidence of adenomas and advanced adenomas in diabetic patients compared with nondiabetics. Their analysis pointed to T2DM as a significant factor in increased adenoma risk (odds ratio [OR] = 1.49, 95% CI: 1.13–1.97, p = 0.0047). Notably, T2DM patients not on diabetic medication were at a higher risk of developing adenomas than those on medication (OR = 2.38, 95% CI: 1.09–5.2, p = 0.03) (Ottaviano et al., 2020). This suggests that early intervention and diabetes management might decrease the likelihood of colon adenoma formation, potentially preventing CRC. Despite these insights, the exact mechanism through which diabetes enhances CRC risk remains to be fully elucidated. Cross-sectional studies in the American medical system have indicated that long-term insulin treatment might increase colon adenoma risk in T2DM patients, possibly due to hyperinsulinemia combating insulin resistance-induced hyperglycemia (Wong et al., 2012). In addition, hyperinsulinemia has been shown to escalate colon adenoma incidence and diminish apoptosis in colon mucosal cells (Keku et al., 2005). The receptor for advanced glycation end products and its ligands have also been implicated in CRC development and progression (Azizian-Farsani et al., 2020). These findings collectively highlight diabetes as a significant risk factor, emphasizing the need for further investigation to develop advanced therapeutic strategies for CRC in diabetic patients. In our research, we first identified an lncRNA, AL049796.1, which plays a pivotal role in the modulation of CRC progression among T2DM patients. Analysis of clinical samples indicated a notably higher expression of AL049796.1 in the CRC tissues of T2DM patients compared with those from nondiabetic individuals. Moreover, our in vitro studies demonstrated that elevated HG could trigger an increase in AL049796.1 expression within CRC cells. Targeted silencing of AL049796.1 was found to significantly curtail the HG-induced enhancement of CRC cells proliferation, invasion, and migration. Similarly, our animal experiments showed an elevated expression of AL049796.1 in the subcutaneous CRC tumor tissues of T2DM mice compared with those in standard tumor-bearing mice, with the silencing of AL049796.1 markedly inhibiting tumor growth in T2DM mice. These findings suggest that AL049796.1 could be a key mediator in the acceleration of HG-induced CRC progression.
Within mammals, the TEAD family comprises four members: TEAD1, TEAD2, TEAD3, and TEAD4, all sharing a similar DNA-binding domain known as TEA at their N-terminal (Stauffer et al., 2012). Biochemical and functional analyses have demonstrated that this TEA domain can bind to the DNA sequence 5′-GGAATG-3′ within the promoter regions of TEAD target genes (Chen et al., 2012). As evolutionarily conserved transcription factors, TEADs play pivotal roles in development and also function as oncogenes in the emergence and progression of various cancers (Sun and Irvine, 2011). They regulate several oncogenic genes, including CTGF, Cyr61, Anexelekt, myelocytomatosis oncogene, and baculoviral IAP repeat-containing 5, positioning them as key mediators in normal cellular growth and carcinogenesis. Particularly, TEAD1 and TEAD4 have been closely linked to cancer development. Research has highlighted the synergistic action of TEAD1 and YAP in stimulating cell growth and oncogene upregulation, contributing to the advancement of skin melanoma and embryonic rhabdomyosarcoma (Yuan et al., 2015). Liang et al. (Liang et al., 2014) observed an upregulation of TEAD1 in 50 CRC tissue samples, correlating positively with the TNM staging of tumors. Yu and Zhang (2016) reported that TEAD1 overexpression could enhance CRC cell proliferation, whereas its silencing led to reduced cancer cell growth. Furthermore, TEAD1 is known to augment the expression of specificity protein 1 by directly binding to its promoter, indicating tumor progression. Despite these findings, there has been no report on the unique expression patterns of TEAD1 in CRC tissues of patients with diabetes to date.
To elucidate the distinctive features between CRC patients with diabetes and those without, we investigated TEAD1 expression in CRC tissues from both groups and discovered a notable increase in TEAD1 expression in the CRC tissues of diabetic patients compared with nondiabetic individuals. Interestingly, the increase in TEAD1 mRNA was less pronounced than that observed at the protein level, hinting at posttranscriptional regulatory anomalies in diabetic patients. Focusing on miRNAs, known posttranscriptional regulators, we conducted a bioinformatics analysis and subsequent verification through luciferase assays. This revealed that miR-200b-3p can bind to the 3′UTR of TEAD1 mRNA, leading to negative regulation of its protein expression. To determine if diabetes influences the miR-200b-3p/TEAD1 pathway in CRC progression, we examined possible correlations in CRC tissues from patients with and without diabetes. Our findings indicated that the negative correlation between miR-200b-3p and TEAD1 protein was exclusive to nondiabetic CRC tissues. There was no correlation between miR-200b-3p and TEAD1 mRNA levels in diabetic versus nondiabetic CRC tissues, nor was there a significant difference in miR-200b-3p levels in CRC tissues between the two patient groups.
What disrupts the ability of miR-200b-3p to negatively regulate TEAD1 protein expression without altering its abundance? Current research and literature suggest the involvement of specific lncRNA acting through the ceRNA hypothesis. According to this hypothesis, lncRNAs can bind to microRNAs at their seed regions, competitively inhibiting the ability of miRNA to regulate target RNAs. Our findings show a marked increase in AL049796.1 levels in the CRC tissues of diabetic patients compared with nondiabetic ones. Furthermore, in vitro assays reveal that the expression of AL049796.1 is upregulated upon HG induction in CRC cells, and silencing AL049796.1 significantly dampens the HG-triggered upsurge in cell proliferation, migration, and invasion. Mechanistic studies suggest that AL049796.1 hampers the binding of miR-200b-3p to TEAD1 mRNA, thus inhibiting TEAD1 mRNA translation. Interestingly, our animal experiments have shown that the increased expression of AL049796.1 in tumor tissues of CRC xenograft mice is not a cause of hyperglycemia, but a consequence of it. More importantly, knocking down of AL049796.1 did not affect the growth of CRC xenografts in conventional nude mice. However, it significantly inhibited the growth of CRC xenografts in nude mice with T2DM, suggesting that AL049796.1 is an independent regulatory factor for CRC progression enhanced by HG. Overall, our research establishes AL049796.1 as a crucial independent factor influencing the miR-200b-3p/TEAD1 axis in the progression of CRC in diabetic patients, positioning it as a more viable target for precise CRC prevention and treatment strategies in this demographic.
Epidemiological and clinical research increasingly confirms that CRC has genetic components, with various lncRNAs playing crucial roles in its regulation and progression (Ni et al., 2021; Sunamura et al., 2016; Yang et al., 2016). Within the realm of diabetes research, lncRNAs are increasingly recognized for their impact on both the development of the disease and its subsequent complications. Specifically, lncRNA-Diabetes Regulated anti-inflammatory RNA is known to contribute to anti-inflammatory responses, whereas the reduction of taurine upregulated 1 expression under HG conditions adversely affects the functioning of pancreatic islet cells (Reddy et al., 2021; Zhang et al., 2021). Furthermore, SOX2 overlapping transcript has been shown to mitigate diabetic kidney disease, and small nucleolar RNA host gene 16 improves vascular function in diabetes cardiomyopathy through ceRNA mechanisms (Cai et al., 2021; Chen et al., 2021). Despite these insights, research on the role of lncRNAs in modulating CRC progression in diabetic patients remains limited, indicating a gap in understanding and potential therapeutic targets. In our research, we specifically discovered that when shielding the pathological conditions associated with diabetes, the silencing of AL049796.1 no longer has the effect on inhibiting the proliferation, invasion, and migration of CRC cells, as well as the growth of subcutaneous CRC tumors. We posit that the silencing of AL049796.1 can only exert its anti-CRC progression effect under HG conditions, which may be related to its higher expression in CRC tissues of diabetics compared with those without diabetes. However, this does not mean that the conclusion that AL049796.1 promotes CRC progression is flawed, but rather emphasizes its response attribute only to HG. Although further investigation is required to identify the upstream regulatory factors of AL049796.1, our results compellingly demonstrate that AL049796.1 autonomously promotes the progression of CRC in HG environments.
Our study is the first to identify AL049796.1 as a promising therapeutic target and early screening marker for CRC in diabetic patients. Given the known vulnerability of lncRNAs to degradation due to their single-stranded structure, AL049796.1’s application faces inherent limitations. However, emerging research provides solutions by highlighting the potential of short or small open reading frames (smORFs) within lncRNA sequences to encode stable polypeptides (SEPs), offering enhanced stability over lncRNAs. This advancement suggests a more reliable clinical testing method for SEPs in bodily fluids, overcoming the challenges of lncRNA detection. Our preliminary investigation has discovered an smORF within AL049796.1, setting the stage for future studies to explore its ability to encode an SEP and its association with CRC progression in diabetics. Furthermore, we aim to validate the role of AL049796.1 in inhibiting CRC progression in diabetic conditions using mouse models and investigate precise siRNA therapies targeting CRC cells. This foundational research provides robust theoretical backing for exploring AL049796.1 as a novel intervention in CRC management for diabetic patients.
In summary, our findings robustly endorse AL049796.1 as an innovative candidate for the targeted prevention and treatment of CRC in individuals with diabetes. The independent role in driving forces of tumorigenesis in CRC underscores the potential of AL049796.1 and its related products used as valuable markers for evaluating CRC risk in diabetic patients.
Footnotes
Ethics Statement
All patients signed the written informed consent. The study was approved by the Ethics Committee of the Harbin Medical University cancer hospital (Approval. No. HMUIRB0222021). Procedures operated in this research were completed in keeping with the standards set out in the Announcement of Helsinki and laboratory guidelines of research in China. Furthermore, all procedures involving mice, including their care, were conducted in strict compliance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals, having been sanctioned by the Ethics Committee of Harbin Medical University cancer hospital (Approval. No. HMUIRB0222021).
Availability of Data and Materials
Data will be made available on request.
Authors’ Contributions
Yan Liu, Qi Wang, Zicheng Sun, Xiaohong Lv, and Xiaojun Zhou: concept, article writing, editing, and review; Haijun Chen, Luxiao Yue, Jiachen Yang, and Zhe Li: data collection and analysis, and article preparation. All authors have read and approved the submission of the article.
Disclosure Statement
The authors have declared that no competing interests exist.
Funding Information
This study was supported by the Natural Science Foundation of Heilongjiang Province (
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.