
Abstract
Bisphenol A (BPA) is the most common endocrine disruptor that has toxicity to the reproductive system and male infertility. However, the underlying mechanisms of BPA’s toxicity to Sertoli cells remain poorly understood. Cellular communication network factor 5 (CCN5) is reported to regulate cell proliferation, apoptosis, and differentiation. Our study demonstrated a significant elevation of CCN5 expression in the testis of nonobstructive azoospermia patients and TM4 Sertoli cells exposed to BPA. Knockdown of CCN5 reduced apoptotic cells after BPA treatment, as determined by flow cytometry and terminal deoxynucleotidyl transferase dUTP nick end labeling assays. Cells exposed to BPA showed increased expressions of Bax and cleaved poly(ADP-ribose) polymerase, decreased expression of Bcl-2, as well as elevated activities of caspase-3 and caspase-9 in BPA-induced TM4 cells, which were reversed by CCN5 inhibition. Loss of CCN5 declined phosphorylation of protein kinase R-like endoplasmic reticulum kinase and eukaryotic translation initiation factor 2A and decreased activating transcription factor 4 and C/EBP-homologous protein in BPA-treated cells. Furthermore, silencing CCN5 blocked BPA-induced phosphorylation of p38 and c-Jun N-terminal kinase (JNK). Administration of anisomycin, a mitogen-activated protein kinase (MAPK) activator, reversed the effects of CCN5 knockdown on BPA-induced endoplasmic reticulum (ER) stress and apoptosis. Taken together, CCN5 promotes apoptosis and ER stress in Sertoli cells exposed to BPA by activating the p38/JNK MAPK signaling pathway.
Introduction
Male infertility is a common public health problem with the decline in fertility worldwide (Aitken, 2024; De Jonge et al., 2024). Azoospermia is classified as obstructive azoospermia (OA) and nonobstructive azoospermia (NOA). OA is mostly due to vas deferens obstruction, and OA patients have normal spermatogenesis (Nicopoullos et al., 2004). However, NOA fails to produce mature sperm because of spermatogenesis dysfunction (Kumar, 2013). Furthermore, various factors, including genetics, environmental endocrine chemicals, diet, and psychological stress, can lead to abnormal spermatogenesis (Knapke et al., 2022; Neto et al., 2016). Thus, it is desirable to investigate the molecular mechanism of NOA.
Bisphenol A (BPA) is a ubiquitous endocrine disruptor in polycarbonate plastics and epoxy resins (Ehrlich et al., 2014). It is shown that BPA may bind to estrogen receptors to act as an estrogen agonist (Gould et al., 1998). At the same time, studies demonstrate that BPA inhibits the binding of androgens with androgen receptors, resulting in an antiandrogenic activity (Lee et al., 2003). Based on its estrogenic and antiandrogenic features, increasing studies have been focused on BPA’s toxic effects on male infertility. Meeker et al. demonstrate an inverse relationship between high BPA concentrations and sperm count and motility, alongside a positive correlation between high BPA levels and sperm DNA damage (Meeker et al., 2010b). In addition, they reveal that high BPA concentrations are associated with increased serum follicle-stimulating hormone (FSH) levels, as well as decreases in inhibin B concentration, the FSH/inhibin B ratio, and the estradiol/testosterone ratio (Meeker et al., 2010a). Vitku et al. report a negative association between seminal BPA levels and sperm concentration, total sperm count, and morphology (Vitku et al., 2016). In animal models, BPA exposure decreases testosterone levels, sperm counts, and sperm motility (Herath et al., 2004; Tiwari and Vanage, 2013). BPA administration in adult mice may alter the spermatogonia/Sertoli cell ratio and adversely affect the offspring (Karmakar et al., 2020). It is known that Sertoli cells are the somatic cells in the testis, and they provide structural support, secrete cytokines, and format the blood–testis barrier during spermatogenesis (Mruk and Cheng, 2015). However, the molecular mechanism by which BPA induces damage to Sertoli cells is not fully understood.
CCN5 (also named WISP2) is a critical member of the cellular communication network (CCN) family of matricellular proteins. CCN5 expression is widely distributed in diverse tissues, such as the brain, heart, lung, testis, and ovary (Gray et al., 2007). Previous studies suggest that CCN5 has potential roles in inhibiting cell proliferation and promoting cell apoptosis and endoplasmic reticulum (ER) stress (Borkham-Kamphorst et al., 2023; Ji et al., 2020; Lake et al., 2003). As an estrogen-responsive gene, CCN5 may be upregulated in human breast cancer cells by environmental estrogens, such as BPA (Inadera et al., 2000). Evidence in young boars further suggests the involvement of CCN5 expression in Sertoli cell proliferation (Berger et al., 2019). However, whether CCN5 mediates BPA-induced spermatogenesis dysfunction is unclear. This study aimed to investigate the potential role of CCN5 and its underlying mechanisms in BPA-induced toxicity of Sertoli cells.
Materials and Methods
Clinical specimens
This work was performed following the Declaration of Helsinki, and the experiments were approved by the Ethics Committee of Shanxi Children’s Hospital (No. IRB-KYYN-2024–002-1). The agreements and informed consent had been obtained from all involved patients. Testicular biopsy tissues were collected from healthy, OA, and NOA patients. Healthy and OA patients with normal spermatogenic function were defined as the controls to compare with NOA patients. The detailed information for these patients is presented in Table 1.
Detailed Information of Clinical Specimens
FSH, follicle-stimulating hormone; LH, leuteinizing hormone; T, testosterone; OA, obstructive azoospermia; NOA, nonobstructive azoospermia.
Cell culture and transfection
The normal mouse testis Sertoli TM4 cells were cultured in DMEM/F12 (Dulbecco's Modified Eagle Medium/Nutrient Mixture F-12; Biosharp Life Sciences, Hefei, China) supplemented with 2.5% fetal bovine serum (TianHang, Huzhou, China) and 7.5% horse serum (Solarbio Science & Technology, Co., Ltd., Beijing, China) in the incubator with 5% carbon dioxide at 37°C.
To determine BPA’s effect, TM4 cells were administrated with different concentrations of BPA (0, 10, 20, 40, 80 µM) (Macklin Biochemical Technology Co., Ltd., Shanghai, China) for 24 h. As is well known, studies on BPA at low doses may be correlated with daily human exposure in real-world scenarios. However, several works concentrate on the toxicological effects and molecular mechanisms of BPA using supraphysiological doses. It may be pertinent to certain poisoning scenarios or occupational exposures. BPA promoted Sertoli cell death dose dependently, with a significant effect at 80 µM. Consequently, high doses of BPA (80 µM) were used to study in further experiments.
To investigate the function of CCN5, three pairs of the short hairpin RNA (shRNA) plasmids against CCN5 (shCCN5#1, shCCN5#2, and shCCN5#3) were synthesized. The empty shRNA plasmid (shNC) was used as a negative control. These sequences used for shRNAs are shown in Table 2. The shRNA plasmids were transiently transfected into TM4 cells using Lipofectamine 3000 (Invitrogen Corporation, Carlsbad, CA, USA) for 48 h. Two pairs of shCCN5 that reduced CCN5 mRNA expression more significantly, according to real-time polymerase chain reaction (PCR), were used for further experiments.
shRNA Sequences
shCCN5, short hairpin cellular communication network factor 5; shNC, short hairpin normal control.
To activate the mitogen-activated protein kinase (MAPK) signaling pathway, cells were treated with 50 ng/mL anisomycin (a MAPK activator) (Macklin Biochemical Technology Co., Ltd.) for 24 h.
3-(4, 5-Dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide assay
For the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay, TM4 cells were seeded in a 96-well plate at the density of 5 × 103 per well in the presence or absence of BPA for 24 h. Next, the medium was added with 50 µL of the MTT solution (Biosharp Life Sciences). Four hours later, the formatted purple crystals were dissolved with 150 µL dimethyl sulfoxide (DMSO; Solarbio Science & Technology, Co., Ltd.). The optical density was measured with a microplate reader (Bio-Tek Instruments, Winooski, VT, USA) at 490 nm.
Flow cytometry assay
The apoptosis of TM4 cells was measured using an Annexin V-FITC/PI Apoptosis Assay Kit (Biosharp Life Sciences). In brief, cells were washed in phosphate buffer saline (PBS) and resuspended with binding buffer. Then, cells were stained with 5 µL Annexin V-FITC and 10 µL propidium Iodide (PI) following the manufacturer’s instructions. Cell apoptosis was analyzed by NovoCyte flow cytometry (Agilent Technologies, Santa Clara, CA, USA).
Terminal deoxynucleotidyl transferase dUTP nick end labeling staining
Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay was performed using the In Situ Cell Death Detection Kit (Roche Diagnostics, Basel, Switzerland) according to the manufacturer’s manual. Cells were permeabilized with 0.1% Triton X-100 (Beyotime Biotechnology, Shanghai, China) for 15 min and incubated with TUNEL reaction solution for 60 min in the dark at 37°C. After washing in PBS, cells were counterstained with 4′,6-diamidino-2-phenylindole (DAPI; Aladdin Reagents Co., Ltd., Shanghai, China) for 5 min. Finally, the slides were observed under a fluorescent microscope (Olympus, Tokyo, Japan), and the apoptotic cells were labeled with red fluorescence.
Measurements of caspase-3 and caspase-9 activities
The activities of caspase-3 and caspase-9 were detected using the commercial kits provided by Beyotime Biotechnology. The experimental procedure was performed according to the description provided by the company. After cell lysis, the protein extracts were quantified using the Bradford Protein Assay Kit (Beyotime Biotechnology). The proteins were incubated with Ac-DEVD-pNA or Ac-LEHD-pNA substrate for 1 h at 37°C. The absorbance of the pNA product was determined by the microplate reader (Bio-Tek Instruments) at 405 nm.
Immunohistochemical and immunofluorescence staining
For immunohistochemical staining, testicular tissues were fixed in 4% paraformaldehyde and embedded in paraffin. The 5-µm tissue sections were subjected to antigen retrieval and blocked in 1% bovine serum albumin (BSA). Then sections were incubated with the primary antibody against CCN5 (ABclonal Biotechnology, Shanghai, China) overnight at 4°C, followed by the incubation with horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG for 1 h at 37°C. The immunohistochemistry color was visualized with diaminobenzidine (DAB; MXB Biotechnologies, Fuzhou, China), and the nucleus was counterstained with hematoxylin (Solarbio Science & Technology, Co., Ltd.).
For immunofluorescence staining, cells were fixed in 4% paraformaldehyde and permeabilized with 0.1% Triton X-100. Then, cells were incubated with 1% BSA, followed by the primary antibodies against CCN5 (ABclonal Biotechnology) and C/EBP-homologous protein (CHOP; Proteintech Group, Inc., Wuhan, China) overnight at 4°C. Next, cells were stained with Cy3-conjugated goat anti-rabbit (Abcam, Cambridge, UK) for 1 h at room temperature. DAPI was added to cells to stain the nucleus. All slides were observed and taken with a microscope (Olympus).
Real-time PCR
To detect gene expression by real-time PCR, total RNA was extracted using the TRIpure reagent (BioTeke Corporation, Beijing, China) and transcribed into cDNA with the All-in-One First-Strand SuperMix Kit (Magen Biotechnology Co., Ltd., Guangzhou, China). Real-time PCR experiments were run on the ExicyclerTM 96 Real-time PCR system (Bioneer Corporation, Daejeon, Korea) using the SYBR Green Real-time PCR Kit (Solarbio Science & Technology, Co., Ltd.). The housekeeping gene GAPDH (glyceraldehyde-3-phosphate dehydrogenase) was used as an internal control. The relative expression data were calculated by the 2−△△CT method. The specific primers are listed in Table 3.
Primer Sequences
CCN5, cellular communication network factor 5; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
Western blot
For the Western blot assay, whole-cell protein extracts and nuclear fractions were prepared using a RIPA buffer contained with PMSF (phenylmethanesulfonyl fluoride; Solarbio Science & Technology, Co., Ltd.) or the Nuclear Protein Extraction Kit (Solarbio Science & Technology, Co., Ltd.). The concentrations of protein extracts were determined with a BCA Protein Kit (Solarbio Science & Technology, Co., Ltd.). Subsequently, protein samples were separated on sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a polyvinylidene fluoride membrane. Membranes were incubated in a blocking solution (Solarbio Science & Technology, Co., Ltd.) for 1 h at room temperature, followed by the following primary antibodies overnight at 4°C: anti-CCN5 (ABclonal Biotechnology), anti-Bax (ABclonal Biotechnology), anti-Bcl-2 (ABclonal Biotechnology), anti-cleaved poly(ADP-ribose) polymerase (cleaved PARP; ABclonal Biotechnology), anti-CHOP (Proteintech Group, Inc.), anti-protein kinase R-like endoplasmic reticulum kinase (PERK; Affinity Biosciences, Changzhou, China), anti-phospho-PERKThr982 (Affinity Biosciences), anti-eukaryotic translation initiation factor 2A (EIF2α; Affinity Biosciences), anti-phospho-EIF2αSer51 (Affinity Biosciences), anti-activating transcription factor 4 (ATF4; Affinity Biosciences), anti-p38 (Affinity Biosciences), anti-phospho-p38Thr180/Tyr182 (Affinity Biosciences), anti-c-Jun N-terminal kinase (JNK; Affinity Biosciences), anti-phospho-JNKThr183/Tyr185 (Affinity Biosciences), anti-GAPDH (Proteintech Group, Inc.), and anti-Histone H3 (GeneTex International Corporation, Alton Pkwy Irvine, CA, USA). Then blots were labeled with HRP-conjugated goat anti-rabbit (Solarbio Science & Technology, Co., Ltd.) and HRP-conjugated goat anti-mouse (Solarbio Science & Technology, Co., Ltd.) secondary antibodies for 1 h at 37°C. Finally, the target proteins were visualized using a Chemical ECL Substrate (Solarbio Science & Technology, Co., Ltd.), and the gray value was analyzed by the Gel-Pro-Analyzer software (Media Cybernetics, Bethesda, MD, USA).
Statistical analysis
Data were expressed as mean ± SD (standard deviation) and analyzed using SPSS software. The comparisons were performed with the one-way analysis of variance followed by the Bonferroni post hoc test with 95% confidence intervals. The partial Eta-squared was calculated to estimate effect size. A p value less than 0.05 was regarded as statistical significance.
Results
CCN5 expression was upregulated in NOA patients and BPA-treated Sertoli cells
To identify the differentially expressed genes (|log2FC| > 1, p < 0.05) in the testes of patients with NOA and OA, we analyzed the database of GSE145467 and GSE108886 from the Gene Expression Omnibus (Fig. 1A). Based on the overlapped upregulated genes in GSE145467 and GSE108886, the enriched Gene Ontology (GO) terms were analyzed using Database for Annotation, Visualization, and Integrated Discovery (DAVID). Results showed that compared with OA patients, the upregulated genes in NOA patients were mainly enriched in pathways related to reproductive function, including “signal transduction,” “endoplasmic reticulum,” “apoptotic process,” “male gonad development,” and “response to steroid hormone” (Fig. 1B). It indicated that ER stress and apoptosis might be associated with spermatogenesis dysfunction. We further analyzed the data in BPA-treated Sertoli cells from GSE4650 and presented the overlapped genes from these databases using a Venn diagram (Fig. 1C). The heatmaps in Figure 1D showed the expression profiles of the 10 co-upregulated genes in the three databases, in which CCN5 expression was the most significantly increased. Then, we validated CCN5 expression alteration in clinical samples. Compared with healthy individuals and OA patients, CCN5 mRNA and protein were significantly elevated in the testis of NOA patients (Fig. 1E, F). Immunohistochemical analysis showed high CCN5 expressions in the Sertoli cells of NOA patients (Fig. 1G). The results suggested a possible role of CCN5 expression in Sertoli cells during spermatogenesis.
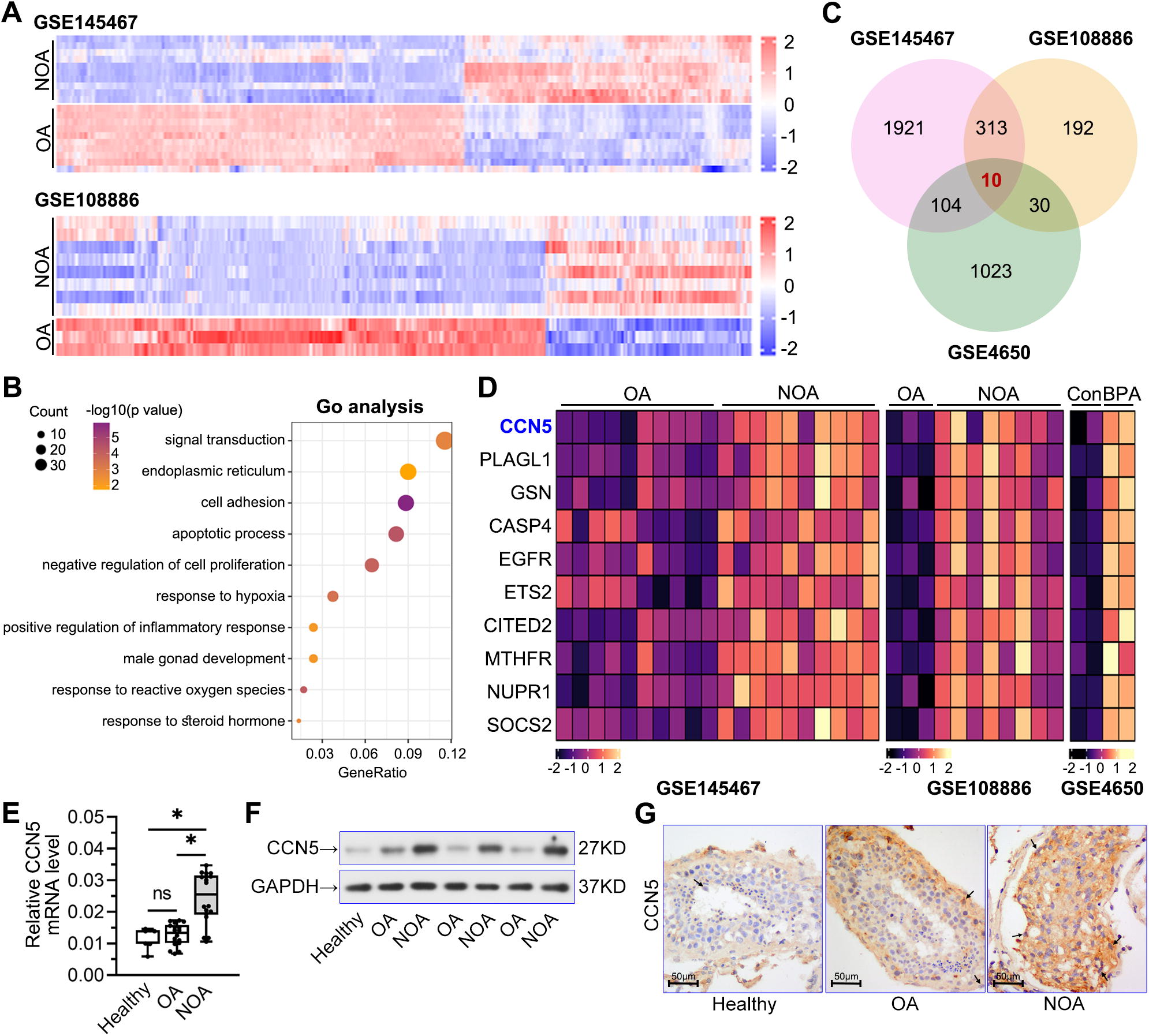
CCN5 expression was upregulated in NOA patients and BPA-treated Sertoli cells.
BPA treatment increased CCN5 expression in TM4 Sertoli cells
To confirm the expression of CCN5 in response to BPA, the murine TM4 Sertoli cells were treated with different concentrations of BPA for 24 h. As shown in Figure 2A–C, BPA administration reduced cell viability and increased CCN5 mRNA and protein levels in a dose-dependent manner, with a significant effect at 80 µM. Immunostaining results in Figure 2D showed that 80 µM BPA increased CCN5 levels in Sertoli cells. These data suggested an involvement of CCN5 in BPA’s toxicity on TM4 Sertoli cells. Although BPA at low doses is correlated with real-world scenarios, supraphysiological concentrations of BPA are used to study the toxicological effects and molecular mechanisms. It may be relevant to specific poisoning scenarios or occupational exposures. Thus, further experiments were performed using the high doses of BPA at 80 µM.

BPA treatment increased CCN5 expression in TM4 Sertoli cells. TM4 Sertoli cells were treated with BPA (0, 10, 20, 40, 80 μM) for 24 h
CCN5 knockdown attenuated BPA-induced apoptosis in TM4 Sertoli cells
Then, we used three pairs of shRNAs against CCN5 to suppress its mRNA and protein expressions (Fig. 3A, B). Among them, shCCN5#1 and shCCN5#2 demonstrated more significant inhibitory effects. Therefore, we selected these two pairs for further experiments. As expected, these two pairs of shCCN5 reduced the increases in CCN5 mRNA and protein levels in BPA-induced TM4 cells (Fig. 3C, D). Flow cytometry and TUNEL staining assays revealed that CCN5 knockdown inhibited BPA-induced cell apoptosis (Fig. 3E, F). The elevated pro-apoptotic proteins (Bax and cleaved PARP) in BPA-treated cells were downregulated by shCCN5, whereas the decreased antiapoptotic protein Bcl-2 was upregulated (Fig. 3G). CCN5 knockdown declined the increases in caspase-3 and caspase-9 activities in cells treated with BPA (Fig. 3H). These findings indicated that inhibition of CCN5 might attenuate BPA-induced apoptosis of Sertoli cells.
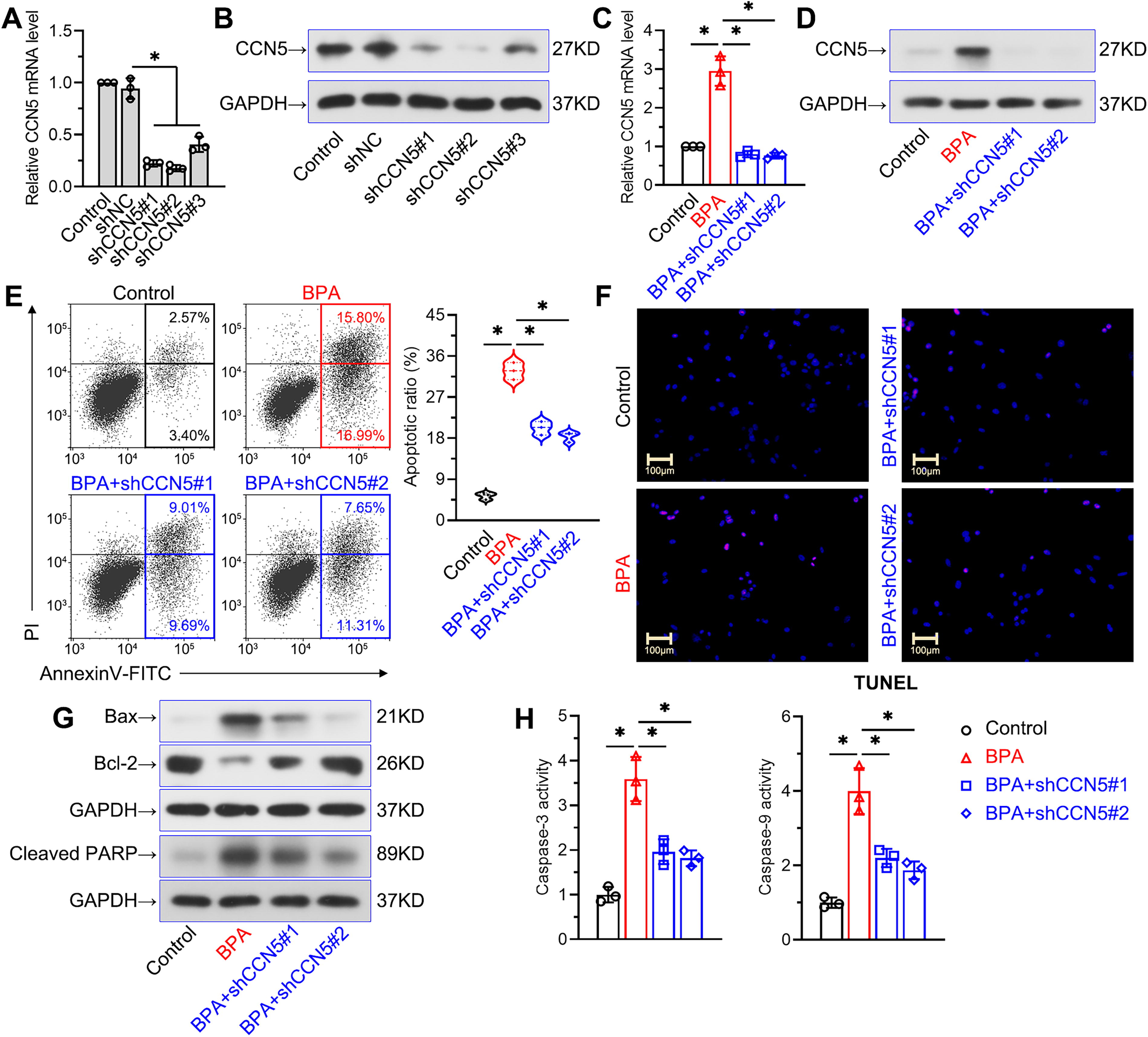
CCN5 knockdown attenuated BPA-induced apoptosis in TM4 Sertoli cells.
CCN5 knockdown inhibited BPA-induced ER stress in TM4 Sertoli cells
We further investigated whether CCN5 mediates the regulation of ER stress in BPA-induced Sertoli cells. As shown in Figure 4A, B, BPA significantly increased the phosphorylation of PERK (p-PERK) and EIF2α (p-EIF2α), along with nuclear ATF4 expression. The knockdown of CCN5 reversed these protein alterations. Furthermore, we observed that CCN5 depletion caused a decline in CHOP expression in BPA-induced TM4 Sertoli cells (Fig. 4C, D). Overall, these findings suggested the positive effects of CCN5 on ER stress in BPA-induced Sertoli cells.
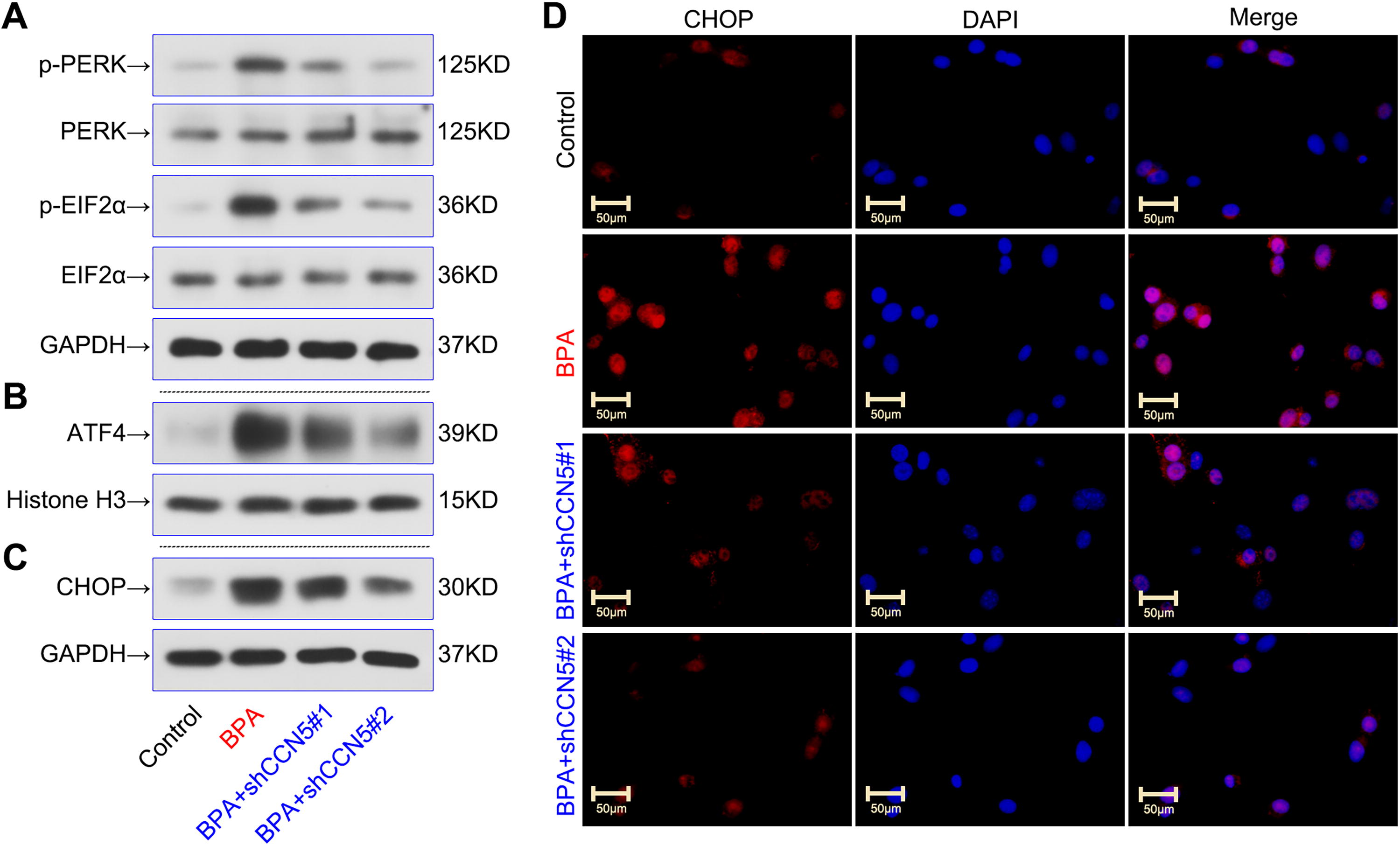
CCN5 knockdown inhibited BPA-induced ER stress in TM4 Sertoli cells. TM4 Sertoli cells were transfected with shCCN5 or shNC for 48 h and then treated with 80 μM BPA for 24 h.
CCN5 affected BPA-induced apoptosis and ER stress by activating p38/JNK MAPK signaling pathway in TM4 Sertoli cells
The overlapped upregulated genes in GSE145467 and GSE108886 were selected for KEGG analysis using DAVID to explore the associated signaling pathways. Results showed that multiple signaling pathways were related to NOA pathogenesis, such as PI3K-Akt, MAPK, Ras, and FoxO (Fig. 5A). It is reported that CCN5 promotes p38 MAPK phosphorylation and inhibits NF-κB signaling activation (Grünberg et al., 2014; Kawaki et al., 2011; Nguyen et al., 2022). NF-κB blocks cell apoptosis by negatively regulating p38/JNK MAPK signaling (Guo et al., 2024). The cross-talk between NF-κB and MAPK signaling pathways prompts the implications of MAPK signaling in CCN5′s effects on Sertoli cell apoptosis. Furthermore, the p38/JNK MAPK signaling pathway is critical for ER stress, inflammation, and apoptosis (Hotamisligil and Davis, 2016; Ono and Han, 2000). Thus, we focused on the p38/JNK MAPK pathway to investigate the mechanisms. Results showed that CCN5 knockdown blocked BPA-induced phosphorylation of p38 and JNK; anisomycin (a MAPK activator) reversed these protein alterations (Fig. 5B, C). In addition, the inhibitory effects of CCN5 knockdown on CHOP expression and cell apoptosis were reversed by anisomycin in BPA-treated cells (Fig. 5D, E). Therefore, these results indicated that the p38/JNK MAPK pathway mediated CCN5′s roles in apoptosis and ER stress of Sertoli cells caused by BPA (Fig. 6).
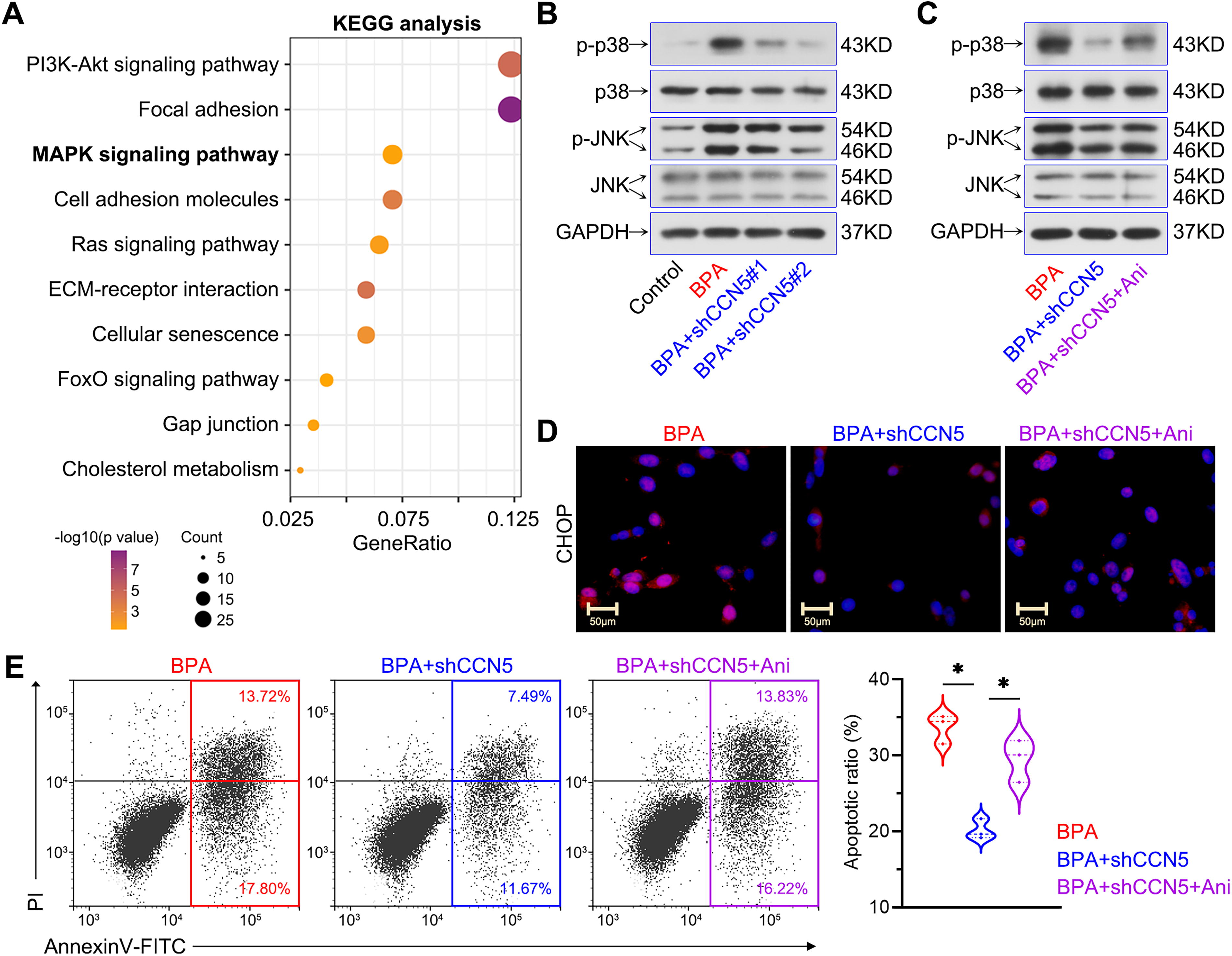
CCN5 affected BPA-induced apoptosis and ER stress by activating p38/JNK MAPK signaling pathway in TM4 Sertoli cells.
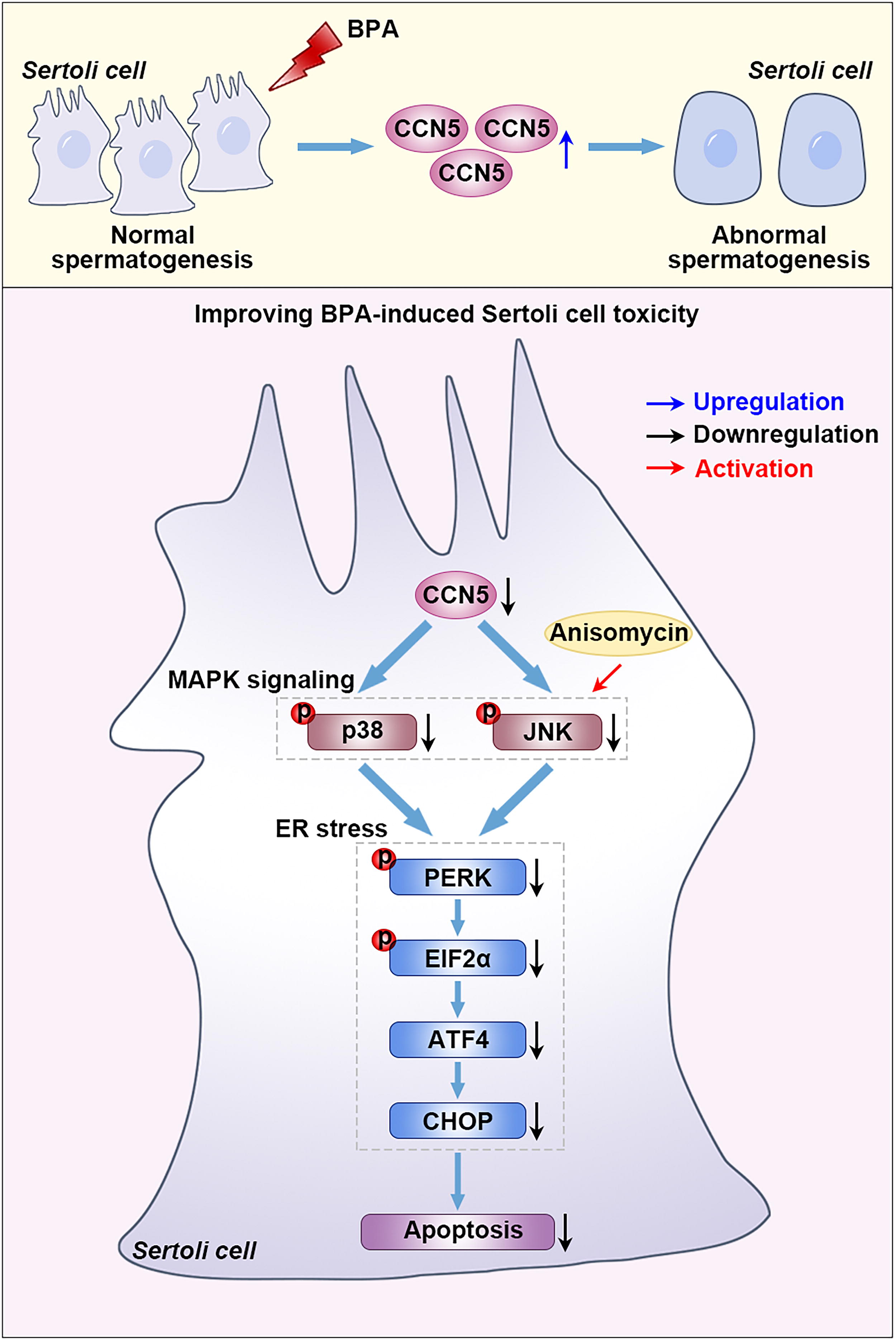
Role of CCN5 in Sertoli cell injury exposed to BPA. Exposure to BPA resulted in an increase in CCN5 expression to induce Sertoli cell injury. Loss of CCN5 in Sertoli cells mediated p38/JNK MAPK signaling inactivation to inhibit ER stress and apoptosis, ultimately attenuating BPA-induced reproductive toxicity. Anisomycin (a MAPK activator) treatment blocked the effects of CCN5 knockdown.
Discussion
In this study, we demonstrated an increased expression of CCN5 in testes of NOA patients and BPA-induced Sertoli cells. Knockdown of CCN5 ameliorated BPA-induced apoptosis and ER stress by inactivating the p38/JNK MAPK signaling pathway.
BPA, a widely utilized chemical, has been reported to exert adverse reproductive effects at environmentally relevant low doses (Peretz et al., 2014). Previous studies report that BPA at low concentrations reduces testosterone levels and impairs sperm motility in rats and mice (Minamiyama et al., 2010; Xi et al., 2011). Mice exposed to low-dose BPA for 28 days exhibit decreased testicular and epididymal sperm counts, along with reduced weights of the testes and seminiferous vesicles (Pflieger-Bruss et al., 2004). Consequently, studies on BPA using environmentally relevant concentrations will be correlated with real-world exposure scenarios. Nevertheless, several studies may utilize supraphysiological doses of BPA to investigate its molecular mechanisms and toxicological effects. For example, BPA exposure over 50 µM promotes Sertoli cell apoptosis by regulating PTEN/Akt pathway and mitochondrial function (Wang et al., 2015; Wang et al., 2017). Inda et al. demonstrate that more than 100 µM BPA induces cell death and apoptosis in Sertoli cells (Iida et al., 2003). Our results showed that high BPA exposure at 80 µM reduced viability and increased apoptosis of Sertoli cells. The high concentration of BPA at 80 µM was used to explore its underlying mechanisms on Sertoli cell toxicity in further experiments.
Based on the public database, we noticed high CCN5 expression in the testes of NOA patients and Sertoli cells exposed to BPA. CCN5 is an estrogen-responsive gene, and its expression is upregulated in BPA-induced MCF-7 cells (Inadera et al., 2000). Our results confirmed a similar alteration of CCN5 expression in BPA-treated TM4 Sertoli cells, indicating a potential role of CCN5 in Sertoli cell injury caused by BPA. Thus, we concentrated on the effects and mechanisms of CCN5 on BPA-induced toxicity in Sertoli cells.
Apoptosis is a programmed cell death process that maintains tissue homeostasis and eliminates damaged or unwanted cells. Evidence reports that Sertoli cell apoptosis induced by BPA leads to testicular dysfunction and infertility (Park et al., 2018; Qian et al., 2014; Wang et al., 2015). In the present work, loss of CCN5 diminished the apoptosis of Sertoli cells exposed to BPA, consistent with the pro-apoptotic role of CCN5 in cardiac myofibroblasts (Jeong et al., 2016). In contrast, Qin et al. show that CCN5 depletion in bone marrow mesenchymal stem cells may promote cell apoptosis in rat livers (Qin et al., 2017). In vascular smooth muscle cells, CCN5 suppresses proliferation and invasion but does not affect apoptosis (Lake et al., 2003; Lake and Castellot, 2003). The differences may be that CCN5 plays different roles in different cell types and diseases.
ER stress is reported to regulate cellular apoptosis by inducing unfolded protein accumulation in the male reproductive system (Adegoke et al., 2023; Karna et al., 2020). The ER sensor protein PERK initiates autophosphorylation and activates EIF2α, leading to ATF4 and CHOP upregulation (Harding et al., 1999; Mei et al., 2013). In rats exposed to BPA, the ER of Sertoli cells shows significant morphological changes (Jiang et al., 2016). Microarray analysis indicates that BPA induces ER stress-related gene upregulation in Sertoli cells (Tabuchi et al., 2006). BPA exposure also activates the ER stress pathway in human hepatocyte cells and mice livers (Hong et al., 2023). Our data suggested that CCN5 deletion reversed the increased expressions of ER stress-related proteins (p-PERK, p-EIF2α, ATF4, and CHOP) in Sertoli cells exposed to BPA. It is consistent with the positive regulation of CCN5 on ER stress and apoptosis in portal myofibroblasts (Borkham-Kamphorst et al., 2023).
To our knowledge, p38/JNK MAPKs are essential for controlling ER stress and apoptosis (Hotamisligil and Davis, 2016; Ono and Han, 2000). Inhibiting MAPKs may abolish the activation of ER stress signaling cascades in vascular smooth muscle cells (Wang et al., 2020). A previous study demonstrates that BPA activates the JNK/p38 MAPK pathway to promote Sertoli cell apoptosis (Qi et al., 2014). CCN5 overexpression induces p38 MAPK phosphorylation in mature adipocytes and osteoblasts (Grünberg et al., 2014; Kawaki et al., 2011). Therefore, we investigated whether the p38/JNK MAPK pathway mediates the mechanism of CCN5 in injured Sertoli cells by BPA. Results showed that CCN5 knockdown reduced the phosphorylated p38 and JNK levels in Sertoli cells exposed to BPA, and activating p38/JNK MAPKs prevented these alterations. It indicated that CCN5 promoted ER stress and apoptosis of BPA-induced Sertoli cells via activating the p38/JNK MAPK pathway. In addition, according to the KEGG analysis, several other pathways may have complementary or competing effects on the regulation of CCN5 in the toxicity of BPA on Sertoli cells, such as the PI3K/Akt signaling pathway. Previous studies have demonstrated that BPA exposure inactivates the PI3K/Akt pathway to impair sperm quality (He et al., 2022; Molangiri et al., 2022). Inhibiting PI3K/Akt leads to an apoptotic cascade to reduce sperm motility after BPA exposure (Koppers et al., 2011). As well, CCN5 has been shown to suppress Akt phosphorylation (Haque et al., 2015; Zhang et al., 2014). Consequently, we speculate that the PI3K/Akt pathway may also mediate CCN5′s pro-apoptotic action on Sertoli cells exposed to BPA. PI3K/Akt may be complementary with p38/JNK MAPK signaling to modulate the role of CCN5 in Sertoli cells. Of course, future research will focus on the precise impacts of PI3K/Akt and pathway cross-talk on CCN5′s effects.
Conclusions
This work suggests that the elevated CCN5 mediates BPA-induced apoptosis and ER stress in Sertoli cells by activating the p38/JNK MAPK pathway (Fig. 6). It proposes a novel insight to clarify the mechanism of BPA-induced toxicity. The CCN5 blocking antibodies, selective inhibitors, small molecular chemicals, or gene editing technology may be promising options to attenuate reproductive toxicity in response to BPA exposure.
Limitations and future directions
Several limitations and promising directions for future research are worth considering. For example, the present study predominantly relies on murine TM4 Sertoli cells. It may not fully represent the biological processes in human Sertoli cells or animals. More experiments using primary human cells or in vivo models will be conducted to address this issue. Second, we focus on the short-term experimental outcomes in this work. However, chronic BPA exposure at environmentally relevant concentrations is related to typical human exposure. Further studies will analyze the long-term effects of BPA at low dosages on reproductive function in male rats, mice, or zebrafish. Long-term follow-up studies in humans are critical for evaluating the long-term effects of occupational BPA exposure. Apart from Sertoli cells, spermatogonia and Leydig cells are also significant for spermatogenesis (Li et al., 2024). We will study the role of CCN5 in other testicular cell types to better clarify its function in spermatogenesis. In addition, possible interventions targeting CCN5 (such as inhibitors or related small compounds) will be explored and developed to determine their impacts on BPA-induced infertility. These strategies may be beneficial in treating spermatogenesis dysfunction and strengthen its therapeutic relevance.
Footnotes
Data Availability
Data used to support the findings of this study are available from the corresponding author upon request.
Authors’ Contributions
L.Z.: Conceptualization, writing—original draft, investigation, and methodology. X.W.: Investigation, methodology, and visualization. Q. Li: Visualization, validation, and resources. Y.S.: Validation and formal analysis. S.Z.: Formal analysis and funding acquisition. J.W.: Methodology and resources. Q. Liu: Conceptualization, project administration, writing—original draft, and funding acquisition.
Author Disclosure Statement
The authors report no conflicts of interest.
Funding Information
This study was supported by the National Natural Science Foundation of China (82472740, 82202323), Tianjin Natural Science Foundation (23JCZDJC00850, 22JCQNJC01640), Tianjin Key Medical Discipline (Specialty) Construction Project (TJYXZDXK-073C), Tianjin Health Science and Technology Project (TJWJ2024XK010, TJWJ2024QN042), and Scientific Research Project of Health Commission of Shanxi Province (2019068).