
Abstract
The taxonomic classification of Babu tea (Family Theaceae) was preliminarily investigated by analyzing its epigenetic morphology and chromosomal features. Morphological observations of three Babu Tea phenotypes (red flower/purple bud, pink flower/purple bud, and white flower/green bud) revealed that the stomata were located exclusively on the lower epidermis, and the anticlinal walls of epidermal cells were deeply undulated. The stomatal apparatus also consisted of two crescent-shaped guard cells, forming a spindle shape with thickened “T-shaped” ends. Furthermore, the outer stomatal arches had smooth surfaces, while the inner ones were shallowly undulated. Finally, subsidiary cells were covered by a shallowly undulated epidermal stratum corneum that was composed of three layers. Their pollen grains exhibited a prolate spherical shape. Based on pollen morphology, the Babu Tea was therefore classified within Sect. Theopsis Coh. St., Camellia. Chromosome counting and karyotype analysis further confirmed that the three phenotypes share a chromosome count of 2n = 2x = 30, with their karyotypes classified as Type 2A. These findings suggest that the three phenotypes share a common evolutionary origin.
Introduction
The selected study area was located in Wangmo, Guizhou, China (106°14'E, 25°19'N), which experiences a tropical humid monsoon climate, an annual average temperature of 16.3°C as well as 1339.7 mm of annual rainfall. Distinct dry and wet seasons also characterize the area, with low precipitation from November to April. In addition, numerous streams and rivers, combined with high humidity from the towering mountains and misty clouds, define the local climate. The terrain is predominantly mountainous, with slopes exceeding 25° and an average altitude of over 1000 m. The soil, which occurs as a thin layer and has a high gravel content, is mostly sandy and is derived from metamorphic rocks. While most tea species may struggle to survive in these challenging conditions, Babu Tea has undergone extensive adaptation, evolving traits such as resistance to cold winters, drought, high summer temperatures, intense sunlight, and poor soil fertility. Currently, over 80,000 Babu Tea plants are cultivated in the study area, and among these, 1763 plants have a diameter at breast height (DBH) of >8 cm, and 1185 are over 100 years old.
Originating from Wangmo, Guizhou, China, Babu Tea is a traditional beverage that has been cherished by the local Miao ethnic group for over a thousand years. It is distinguished by its unique purple buds which are present throughout the plant and remain stable across different seasons. The buds of Babu Tea are rich in catechins, anthocyanins, flavonoids, and amino acids, along with distinct flavor components, thus making it highly valued as a tea. Previous research on the apparent morphology of Babu Tea identified three phenotypes under natural conditions: white flower/green bud, red flower/purple bud, and pink flower/purple bud (Wang et al., 2022). In this study, the similarities and differences among these three phenotypes were explored from morphological and cytological perspectives in order to understand the classification of Babu Tea in theaceae. It is expected that the findings will provide valuable references for future research on germplasm resources and genetic breeding of this unique tea variety.
Materials and Methods
Plant material
In October 2024, the morphological characteristics of three Babu Tea phenotypes (red flower/purple bud, pink flower/purple bud, and white flower/green bud) were measured under natural conditions in Babu Village and Tielu Village, Wangmo, Guizhou, China.
Micro-morphology of leaves
For each phenotype, three plants were randomly selected, and from each plant, three to five mature leaves were collected. The leaves were then carefully cleaned with a brush to remove any dust before cutting 2 × 2 mm samples from the central part of the leaf vein. These samples were subsequently rinsed with 95% ethanol and dehydrated using 100% ethanol, after which they were mounted on double-sided adhesive tape, vacuum coated, and observed under scanning electron microscopy (SEM) Hitachis-800. Images were also captured for analysis.
Micro-morphology of pollen and stigma
Three plants were randomly chosen for each phenotype, with three to five normal, ready-to-bloom flowers subsequently collected from each plant. The pollen was then applied directly onto double-sided adhesive paper without any treatment and coated before being examined under SEM 6490LV to observe pollen morphology, germinal aperture structure and exine ornamentation. In addition, the polar axis length (P) and equatorial axis length (E) of the pollen grains were measured.
Karyotype analysis
Seeds from the three Babu Tea phenotypes were soaked in 200 mg/L of aqueous gibberellin solution for 12 h, after which they were washed and allowed to germinate. Once the root length reached 1–2 cm, samples were collected, washed, and then pretreated with 0.002 mol/L of 8-hydroxyquinoline solution for 1–6 h at room temperature. Following the pretreatment, the samples were rinsed with deionized (DI) water and fixed in Carnoy’s fixative at 4°C for 16–20 h. The washing step was then repeated using DI water, with the samples eventually stored in 70% ethanol at 4°C. When required for further analysis, the preserved materials were washed and the excess water was removed. This was followed by treatment with a mixture of 2% pectinase and 1% cellulase for 1–3 h at room temperature, after which the samples were rinsed with 0.1 mol/L of sodium acetate and subsequently hydrolyzed in 0.1 mol/L HCl solution for 2 min at 60°C. The samples were then washed with DI water, and small sections were cut, placed on slides, stained with DAPI for 2 min, and prepared using a standard squash method. The samples were finally observed in the dark with a fluorescence microscope (Leica DM2500).
For each type of material, 30 cells were selected for chromosome counting. Karyotype data were first obtained from chromosomes from three well-dispersed metaphase cells. Chromosome alignment and measurements were then performed with Photoshop CC, with the karyotype classification system (Stebbins, 1971) used to determine relative lengths and length ratios. Finally, the chromosomes were classified based on the centromere position naming system (Levan et al., 1964), and the karyogram and karyotype patterns were subsequently developed according to standardized recommendations for plant karyotype analysis (Li and Chen, 1985).
Statistical analysis
All measurements were repeated three times and averaged. Microsoft Excel 2007 and SPSS 22.0 (IBM, Inc., Armonk, NY) were used for data processing and analysis.
Results
Micro-morphology of leaves
The micro-morphology of leaves in the three Babu Tea phenotypes shared several common characteristics: the stomata were located exclusively on the lower epidermis, and the anticlinal walls of epidermal cells were deeply undulated. The stomatal apparatus also consisted of two crescent-shaped guard cells, forming a spindle shape with thickened “T-shaped” ends. Furthermore, the outer stomatal arches had smooth surfaces, while the inner ones were shallowly undulated. Finally, subsidiary cells were covered by a shallowly undulated epidermal stratum corneum that was composed of three layers. However, differences in the size of stomatal apparatuses were negligible, with longitudinal lengths of 34.89 μm, 31.26 μm, and 34.29 μm for the red flower/purple bud, pink flower/purple bud, and white flower/green bud phenotypes, respectively (Fig. 1).

SEM images of leaves for the three Babu tea phenotypes. The stomata were located exclusively on the lower epidermis, and the anticlinal walls of epidermal cells were deeply undulated. Stomata apparatus size difference was not obvious, and the longitudinal length of stomata apparatus is 34.89 μm for red flower/purple bud, 31.26 μm for pink flower/purple bud, and 34.29 μm for white flower/green bud. SEM, scanning electron microscopy.
Overall, the micro-morphology of the mesenchymal epidermis in all three Babu Tea phenotypes aligned with the common characteristics observed in the leaf epidermal micro-morphology of Camellia plants. However, the three phenotypes differed in pore size, stomatal apparatus, shape of the adjacent horny membrane, and numbers of stomata, gland scales, and epidermal hairs (Table 1). In contrast, while differences in these features were apparent within the same species, there was general consistency among them in closely related species. As such, these features can provide limited taxonomic differentiation but remain insufficient for intraspecific classification. Based on these findings, the three Babu Tea phenotypes could be classified as belonging to the same plant species.
Sem Data of Babu Tea Leaves
Micro-morphology of pollen and stigma
As shown in Figure 2 and Table 2, the three Babu Tea phenotypes exhibited a pollen P/E ratio of 1.48–1.53, indicating a prolate shape. Pollen size was then classified as medium based on the length of the polar axis but as large when considering the mean equatorial axis. Furthermore, the exine ornamentation displayed a rough reticulation pattern, and three types of germinal apertures were observed, namely sutural germinal grooves, banded germinal apertures, and shuttle-like germinal apertures. These findings suggested that the three Babu Tea phenotypes were conspecific. Notably, the exine ornamentation and morphology of the germinal aperture exhibited more advanced evolutionary characteristics. SEM images of Babu Tea stigmas (Fig. 3) further revealed that the stigma length for all three phenotypes was approximately 3 mm. Additionally, the stigmas had a connate base and a three-lobed structure in the middle to upper region. However, differences were observed in the connate basal length as well as the extent of pollen presence as follows: for the white flower/green bud phenotype, the three-lobed structure was located at the midpoint (1/2) of the stigma, with a small amount of pollen visible; for the red flower/purple bud, the three-lobed structure was at 1/4 of the stigma, with a large amount of pollen observed; finally, for the pink flower/purple bud, the three-lobed structure was at 1/5 of the stigma, and no pollen was observed entering it. These results from SEM observations were consistent with macroscopic observations of the stigma, hence supporting the conclusion that the three Babu Tea phenotypes should belong to the same species.
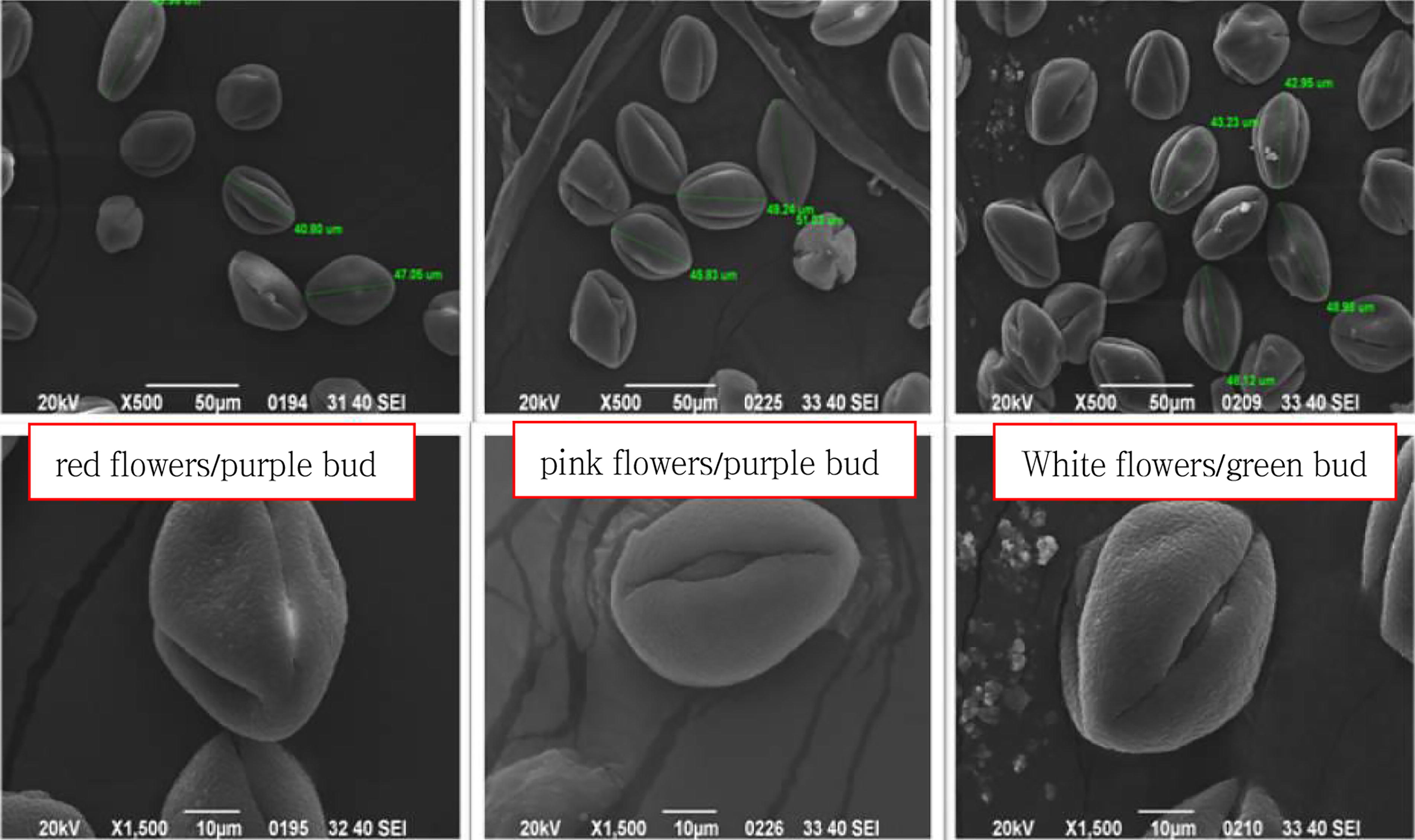
SEM images of pollen for the three Babu tea phenotypes. The 3 Babu Tea phenotypes had pollen P/E ratio of 1.48–1.53 and prolate shape. The exine ornamentation was a rough reticulation; and the germinal aperture had three types: sutural germinal grooves, banded germinal aperture, and shuttle-like germinal aperture.
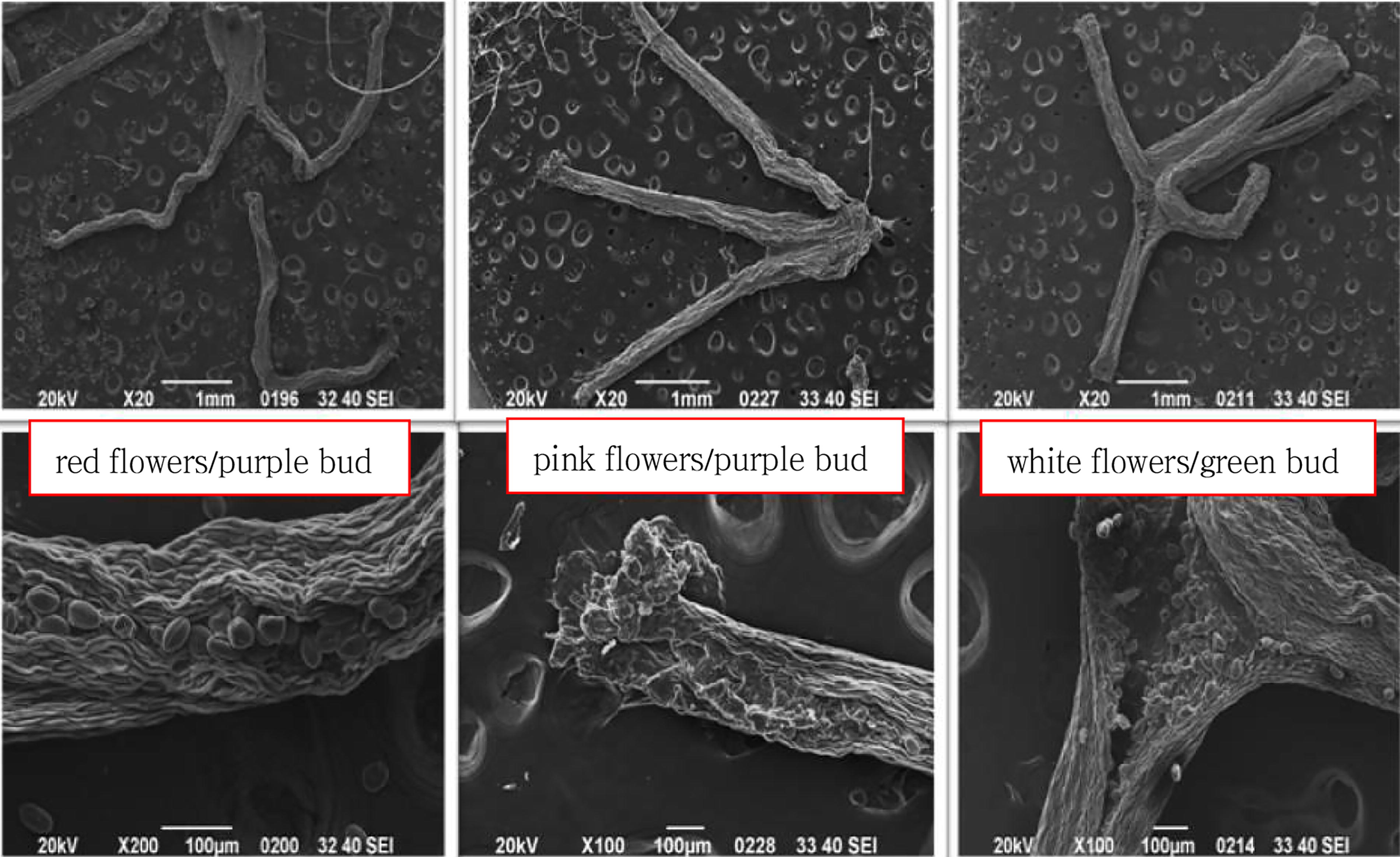
SEM images of stigma for the three Babu tea phenotypes. The stigma length of the three Babu Tea phenotypes was around 3 mm, with a connate base, and 3-lobed was in the middle and upper part; the differences were the length of the basal connate part, 3-lobed was at 1/2 of the stigma of white flower/green bud and a small amount of pollen was observed; 3-lobed was at 1/4 of the stigma of red flower/purple bud, and a large amount of pollen was observed; 3-lobed was at 1/5 of the stigma of pink flower/purple bud, and no pollen was observed to enter the stigma.
Pollen Morphology for the Three Babu Tea Phenotypes
Chromosome count and karyotype analysis
Chromosome count
Chromosome counts, conducted on 30 cells from each of the three Babu Tea phenotypes, revealed that 28 cells (93.33%) of the white flower/green bud phenotype, 26 cells (86.67%) of the red flower/purple bud phenotype and 28 cells (93.33%) of the pink flower/purple bud phenotype had a chromosome count of 30. Based on the criteria for plant karyotype analysis (Li and Chen, 1985), cells with a chromosome count of 30 accounted for over 85% of the total cells examined in all three phenotypes. Therefore, the chromosome count of all three Babu Tea phenotypes was determined to be 2n = 2x = 30 (diploid) (Fig. 4).
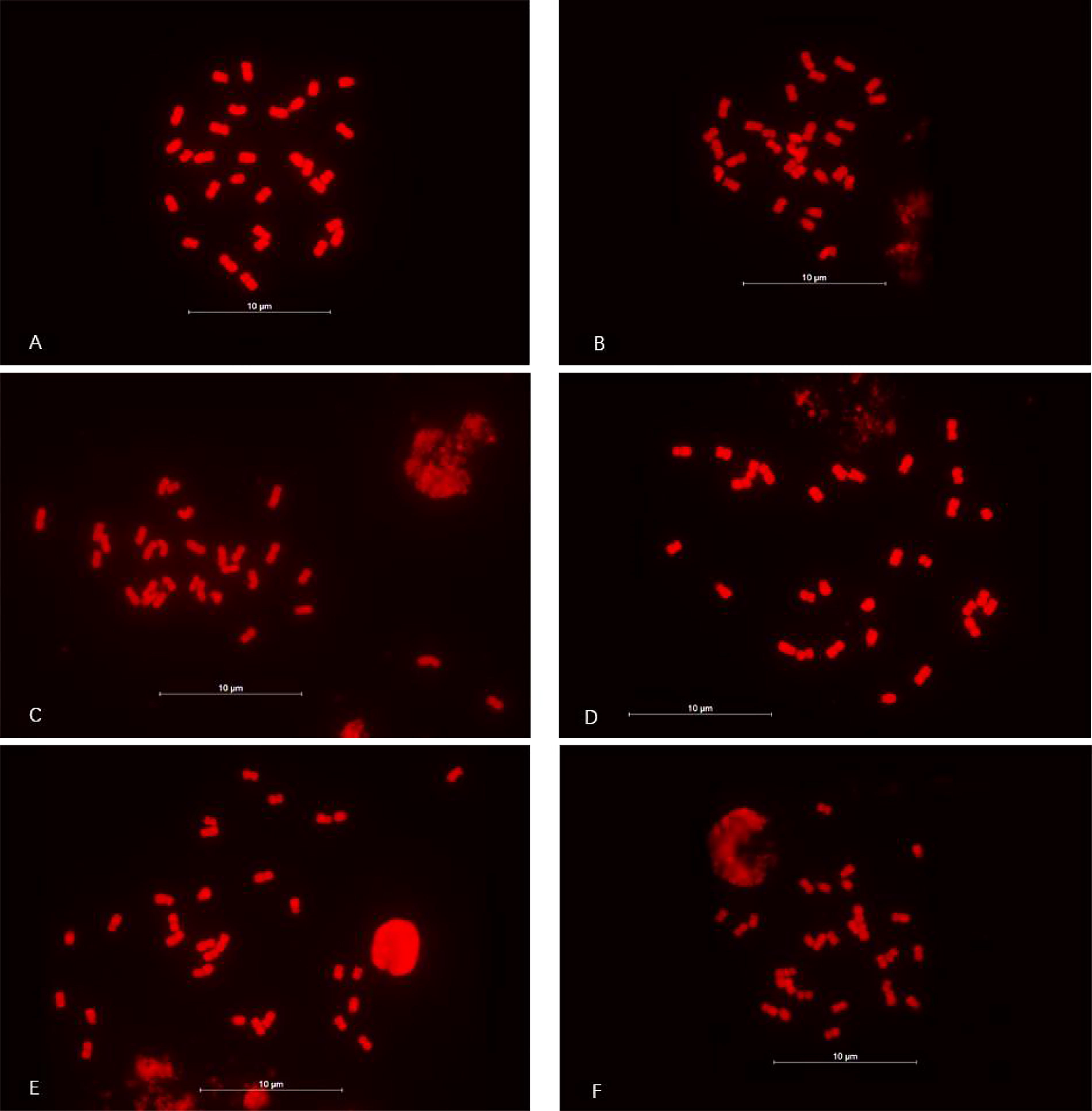
Chromosome count for the three Babu Tea phenotypes. (
Karyotype analysis
The white flower/green bud phenotype exhibited a chromosome count of 2n = 2x = 30 (diploid), with a karyotype configuration of 2n = 2x = 30 = 4M + 20 m + 6st. Additionally, the relative chromosome length was 5.25% to 8.31%, with an average arm ratio of 1.91. The maximum-to-minimum chromosome length ratio was also found to be 1.58, and three chromosomes (20%) had an arm ratio >2:1. Based on these characteristics, this phenotype was classified as Type 2A, with a karyotype asymmetry coefficient of 60.00% (see Table 3). The karyogram and karyotype pattern for the white flower/green bud phenotype are shown in Figures 5 and 6.
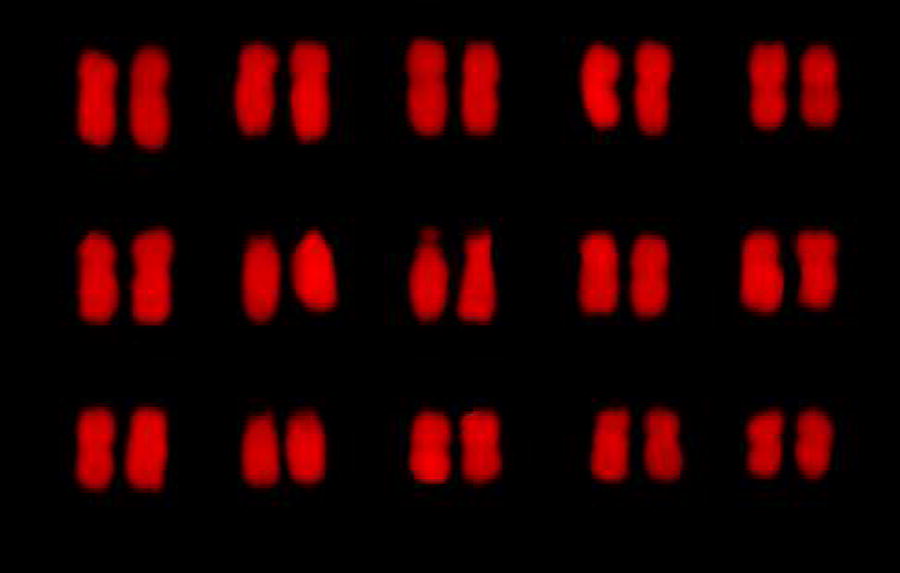
Karyogram of the white flower/green bud Babu Tea phenotype. The white flower/green bud had a chromosome count of 2n = 2x = 30 (diploid).

Idiogram of the white flower/green bud Babu Tea phenotype. The relative length was 5.25%−8.31%, the maximum to minimum chromosome length was 1.58, the count of chromosomes with arm ratio >2:1 was 3 (20%); it was Type 2A; the karyotype asymmetrical coefficient was 60.00%.
Chromosome Parameters for the White Flower/Green Bud Babu Tea Phenotype
The red flower/purple bud phenotype also exhibited a chromosome count of 2n = 2x = 30 (diploid), with a karyotype configuration of 2n = 2x = 30 = 24 m + 4sm + 2st. In this case, the relative chromosome length ranged from 4.61% to 8.24%, while the average arm ratio was 1.53. In addition, the maximum-to-minimum chromosome length ratio was 1.79, and two chromosomes (13.33%) had an arm ratio of >2:1; hence, the phenotype was similarly classified as Type 2A, with a karyotype asymmetrical coefficient of 58.65% (see Table 4). The karyogram and karyotype pattern for the red flower/purple bud phenotype are shown in Figures 7 and 8.

Karyogram of the red flower/purple bud Babu Tea phenotype. The red flower/purple bud had a chromosome count of 2n = 2x = 30 (diploid).

Idiogram of the red flower/purple bud Babu Tea phenotype. The relative length was 4.61%−8.24%, the maximum to minimum chromosome length was 1.79, the count of chromosomes with arm ratio >2:1 was 2 (13.33%); it was Type 2A; the karyotype asymmetrical coefficient was 58.65%.
Chromosome Parameters for the Red Flower/Purple Bud Babu Tea Phenotype
The pink flower/purple bud phenotype exhibited a chromosome count of 2n = 2x = 30 (diploid), with a karyotype configuration of 2n = 2x = 30 = 4M + 20 m + 2sm + 4st. Additionally, the relative chromosome length ranged from 5.06% to 7.78%, while the average arm ratio was 1.57. Moreover, the maximum-to-minimum chromosome length ratio was 1.54, with two chromosomes (13.33%) having an arm ratio of >2:1. Therefore, this phenotype was also classified as Type 2A, with a karyotype asymmetrical coefficient of 57.50% (see Table 5). The karyogram and karyotype pattern for the pink flower/purple bud phenotype are shown in Figures 9 and 10.

Karyogram of the pink flower/purple bud Babu Tea phenotype. The pink flower/purple bud had a chromosome count of 2n = 2x = 30 (diploid).

Idiogram of the pink flower/purple bud Babu Tea phenotype. The relative length was 5.06%−7.78%, the maximum to minimum chromosome length was 1.54, the count of chromosomes with arm ratio >2:1 was 2 (13.33%); it was Type 2A; the karyotype asymmetrical coefficient was 57.50%.
Chromosome Parameters of the Pink Flower/Purple Bud Babu Tea Phenotype
Discussion
Micro-morphology of leaves
In plant classification, leaves rank as the second most important taxonomic feature after flowers and fruits. Indeed, the micro-morphology of leaves has become increasingly significant for the taxonomic identification of challenging taxa in modern plant taxonomy. In Camellia plants, the leaf epidermis exhibits the following common characteristics: epidermal cells are undulated or deeply undulated; stomatal apparatuses are present only on the lower epidermis and are annular, being formed by three subsidiary cells of varying sizes surrounding two crescent-shaped guard cells. Some of these subsidiary cells are often covered by well-developed cuticles, making the boundaries of the ring around the guard cells less distinct; the surface of outer stomatal arches is generally smooth, while the inner margins are nearly smooth or shallowly undulated; the cuticular membranes typically exhibit striped patterns but may also appear granular or scaly. Overall, differences in these leaf epidermal features among species hold taxonomic and identification value for classification within the same genus.
Zhang and Zhuang (2004) examined the leaf epidermal morphology of 25 Camellia species and identified three main types of leaf epidermal stomatal complexes: annular, planar, and irregular, with a minor occurrence of radial types. Their findings suggested that the planar and irregular stomatal complexes evolved directly or indirectly from the annular type, with the irregular one being the most advanced among those observed in Camellia plants. Similarly, Ao et al. (2002) studied the leaf epidermal morphology of 36 Camellia plants and found that the stomatal apparatus was absent in the upper epidermis and restricted to the lower epidermis. The stomatal complexes were also predominantly annular, being characterized by three subsidiary cells of varying sizes surrounding the guard cells. In some cases, these subsidiary cells were covered by well-developed cuticles, making the ring boundaries around the guard cells less distinct. Furthermore, the stomata were shuttle-shaped or elongated ovate-shaped, with the inner edges of the outer stomatal arches exhibiting smooth to shallowly undulated/undulated patterns. Finally, the cuticular membranes near the stomatal apparatus ranged from flat to wrinkled, with surfaces that were either smooth or decorated with multiple ornamentations. These characteristics provide a basis for distinguishing species or varieties but they are not correlated with the plant’s external morphology.
Deng et al. (2018) conducted a quantitative analysis of the microscopic features of leaf blades from 20 Camellia plants and observed variations in the anticlinal wall patterns of the leaf epidermal cells. Specifically, Subgen. Camellia plants predominantly exhibited shallowly undulated patterns, with occasional straight or shallow to undulated forms. In contrast, Subgen. Thea Chang plants were characterized by undulated patterns, while Subgen. Metacamellia Chang plants exhibited deeply undulated ones. These anticlinal wall patterns proved to be valuable for distinguishing taxonomic units, such as genera and subgenera, and for reflecting the phylogenetic relationships within the genus, thereby serving as an important taxonomic tool in the systematic classification of Camellia plants. Furthermore, most Camellia species possess glandular dots, while the presence of epidermal hairs varies significantly among individuals of the same species, making it unsuitable as a taxonomic criterion. In this study, the micro-morphological leaf characteristics of Babu Tea samples aligned with the common traits of leaf epidermal morphology observed in Camellia plants. At the same time, no significant differences were noted among the three Babu Tea phenotypes, thus confirming that they belong to the same species. Moreover, the anticlinal wall pattern of the Babu Tea phenotypes closely resembled that of the Subgen. Metacamellia Chang, is characterized by a deeply undulated structure.
Micro-morphology of pollen and stigma
The exine ornamentation of pollen, the shapes of the germinal apertures, and pollen size are key characteristics used to differentiate plant species within the same genus. The surface ornamentation of tea pollen can be broadly classified into three types: smooth reticulation, rough reticulation, and burrowing reticulation. Among wild primitive tea resources, smooth reticulation accounts for 69.7%, rough reticulation for 15.2%, and burrowing reticulation for 15.1%. In contrast, cultivated tea resources are predominantly characterized by rough reticulation (59.0%), followed by smooth reticulation (30.8%) and burrowing reticulation (10.2%). Thus, cultivated tea resources are primarily associated with rough reticulation, while primitive ones are dominated by smooth reticulation (Shu, 1996; Shu and Chen, 1996; Shu et al., 1998). Similarly, the germinal aperture (groove) can be classified into three types: sutural germinal grooves, banded germinal apertures, and shuttle or ellipsoidal germinal apertures, with resource materials having the latter apertures generally considered to be more evolutionarily advanced. Pollen grain size is typically assessed based on the length of the polar axis as follows: very small pollen: <10 μm; small pollen: 10–24 μm; medium pollen: 25–49 μm; large pollen: 50–99 μm; super-large pollen: 100–199 μm; and ultra-large pollen: ≥200 μm. Alternatively, pollen size can also be classified using the mean equatorial axis length as follows: large pollen: ≥30 μm and small pollen: <30 μm. In addition, the following classification is used for pollen shape, determined by the Polar/Equatorial (P/E) ratio: super-oblate sphere: P/E ratio <0.50; oblate sphere: P/E ratio = 0.50–0.75; nearly-oblate sphere: P/E ratio = 0.75–0.88; round sphere: P/E ratio = 0.88–1.14; nearly-prolate sphere: P/E ratio = 1.14–1.33; prolate sphere: P/E ratio = 1.33–2.00; and super-prolate sphere: P/E ratio ≥2.00. Most tea tree resources from large-leaf species produce large pollen, while those from medium- and small-leaf species produce small pollen. Pollen shapes include round, oblate, nearly-prolate spheres, and prolate spheres, with pollen size evolution generally following a trend from larger to smaller grains.
Li et al. (2005) analyzed the pollen morphology of six Sect. Theopsis Coh. St. plants and found that the characteristics aligned with the general traits of pollen from Camellia plants, including a prolate spherical shape, with the primary interspecies variations apparent in the size and exine ornamentation. For instance, Camellia costei H. Lév. exhibits three-lobed pollen in polar view, an ellipsoid shape in equatorial view, and a pollen size of 45 μm (Ao, 2004), closely resembling the pollen morphology and size of Babu Tea. However, the exine ornamentation of Camellia costei H. Lév. is granular to wrinkled (Ao et al., 2002) and differs significantly from the rough reticulation observed in Babu Tea, thus indicating that they belong to distinct species. Similarly, Xu et al. (2010) noted that the pollen morphology of Camellia rosthorniana was oblate spherical, with pollen dimensions of about 29.13 μm × 40.33 μm. These features were significantly different from the more conservative prolate spherical shape seen in other Sect. Theopsis Coh. St. plants (Ao et al., 2002) and from the pollen morphology of Babu Tea, hence confirming them as different species.
Chromosome karyotype analysis
Chromosomes are the carriers of genetic material, with chromosomal variations serving as a critical source of genetic diversity (Wang, 2012). Unlike external morphological traits, chromosomes are less influenced by environmental factors, hence making them a more reliable indicator for identifying differences and determining genetic relationships among species (Lu et al., 1988). Therefore, this study also explored the chromosomes of Babu Tea which belongs to Sect. Theopsis Coh. St., Subgen. Metacamellia Chang. Li and Liang (1990; Li, 2001) compiled the karyotype data of Camellia plants and observed a stable chromosomal base number of x = 15 across the genus. Most Camellia species are diploid, and their karyotypes, predominantly classified as type 2A or 2B, often exhibit low asymmetrical coefficients. In the present study, karyotype analysis revealed that Babu Tea has a chromosome count of 2n = 2x = 30. Furthermore, the karyotypes of all three Babu Tea phenotypes were classified as Type 2A, with their asymmetrical coefficients ranging from 57.50% to 60.00%. Additionally, the chromosome counts and karyotype classifications of the three phenotypes were identical based on Stebbins’s classification system. Altogether, these findings suggest a common evolutionary origin for the three Babu Tea phenotypes.
Footnotes
Authors’ Contributions
W.Y. and W.J.H. designed the research. W.Y. and W.S. performed the research. L.L.J., W.C., and L.Q.S. analyzed the data. W.Y. wrote the article. W.J.H. revised the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
Doctoral research project of Guizhou Academy of Sciences, grant NO. qiankeyuanRzi [2021]9; Scientific and technological innovation talent team building of Guizhou Institute of Biology, grant NO.2024002; Youth Science Foundation of Guizhou Academy of Sciences, grant NO.qiankeyuanJzi [2023]18.