
Abstract
Retinoblastoma (RB) arising from the retina’s primitive neural precursor cells is a highly aggressive pediatric ocular malignancy. Ubiquitin-conjugating enzyme E2C (UBE2C) is implicated in carcinogenesis, but its role and mechanism in RB remain unexplored. Here, we aimed to explore the effect of UBE2C and its regulatory mechanism in an N6-methyladenosine (m6A) modification method in RB. The expression of UBE2C and methyltransferase-like 3 (METTL3) was determined by quantitative reverse transcription–polymerase chain reaction (qRT-PCR) and Western blotting. After using shRNA and overexpression vectors to modulate the expression of UBE2C and METTL3 in RB cells, cell viability, proliferation, apoptosis, and phosphoinositide 3-kinase-protein kinase B (PI3K-AKT) pathway activity were assessed via cell counting kit-8, 5-ethynyl-2′-deoxyuridine, flow cytometry, and Western blotting assays, respectively. The correlation between METTL3 and UBE2C in RB cells was verified by qRT-PCR, Western blotting, methylated RNA immunoprecipitation, mRNA stability assays. The results showed that UBE2C with high expression in RB enhanced RB cell survival via elevating cell viability and proliferation, as well as suppressing apoptosis. UBE2C activated the PI3K-AKT pathway by promoting the PI3K and AKT proteins. METTL3 upregulated UBE2C expression and enhanced UBE2C mRNA stability via m6A modification. In addition, upregulating METTL3 partly restored the negative effects of UBE2C downregulation on RB cells. In conclusion, METTL3 drives UBE2C overexpression through m6A modification, thereby activating the PI3K-AKT pathway to foster RB progression. This study identifies the METTL3/UBE2C/PI3K-AKT axis as a novel therapeutic target for RB.
Introduction
Retinoblastoma (RB) arising from the retina’s primitive neural precursor cells is the most common primary intraocular malignancy, primarily affecting in children (Markovic et al., 2023; Pareek et al., 2024). With an incidence of approximately 1/15,000–20,000 live births, RB accounts for approximately 2.5–4% of all pediatric cancers and poses significant risks to vision, neurological health, and survival if not diagnosed early (Markovic et al., 2023; Nag and Khetan, 2024; Sheehan, 2020). The biallelic inactivation of the RB1 tumor suppressor gene, located on chromosome 13q14, disrupts the RB protein—a critical regulator of cell cycle progression and apoptosis, which is a well-established driver of RB initiation (Markovic et al., 2023; Sachdeva and O’Brien, 2012). However, emerging evidence suggests that the molecular pathogenesis of RB also involves epigenetic dysregulation, aberrant posttranslational modifications, and activation of oncogenic signaling pathways (Ma et al., 2024; Pareek et al., 2024; Rathore et al., 2023; Zhou et al., 2024). Despite advances in multimodal therapies, including chemotherapy, radiation, and enucleation, advanced and metastatic RB remains a clinical challenge, underscoring the need to identify novel molecular targets for precision therapy.
Ubiquitin-conjugating enzyme E2C (UBE2C), a member of the E2 ubiquitin-conjugating enzyme family, plays a key role in cell cycle regulation via inducing the degradation of key mitotic regulators such as cyclins and anaphase-promoting complex/cyclosome (Wang et al., 2019). Dysregulated UBE2C expression has been implicated in various cancers to promote tumorigenesis. For example, UBE2C with high expression in cervical cancer was reported to drive tumorigenesis (Chiang et al., 2020). In breast cancer, elevated UBE2C expression showed higher grade, stage, and poor survival (Guo et al., 2023). UBE2C overexpression also could accelerate the growth of gastric cancer (Jiang et al., 2024). Despite the oncogenic function of UBE2C in multiple adult malignancies, the role of UBE2C in pediatric cancers, particularly RB, remains unexplored.
A growing body of research has highlighted that posttranscriptional modifications, particularly N6-methyladenosine (m6A), play a significant role in cancer progression (An and Duan, 2022; Ganesan et al., 2024; Jiang et al., 2021). As a key modification in eukaryotic mRNA, m6A dynamically influences RNA splicing, stability, translation, and degradation by a system of “writers” (methyltransferases), “erasers” (demethylases), and “readers” (binding proteins) (Wang et al., 2022). The m6A methyltransferase complex consists of multiple components, with methyltransferase-like 3 (METTL3) being the catalytic core responsible for depositing m6A marks on target transcripts to modulate the fate of target transcripts (Pinello et al., 2018). Dysregulation of METTL3 has been implicated in diverse cancers, where it can act as either an oncogene or tumor suppressor depending on the cellular context. For example, METTL3 induced m6A modification of HDGF mRNA to enhance gastric cancer cell malignancy (Wang et al., 2020). However, METTL3 could prevent papillary thyroid cancer progression by mediating m6A modification of STEAP2 mRNA (Zhu et al., 2022). In RB, only one study revealed that METTL3 promoted RB progression by regulating the phosphoinositide 3-kinase-protein kinase B (PI3K/AKT)/mTOR pathway (Zhang et al., 2020). However, the specific target genes of METTL3, particularly in the regulation of UBE2C, remain unknown.
Due to multiple studies showing the regulatory effect of UBE2C or METTL3 on PI3K-AKT pathway in cancers (Huang et al., 2024; Wang et al., 2024; Zhang et al., 2020), this study aims to investigate the role of UBE2C in RB progression and explore its regulation by METTL3-mediated m6A modification and PI3K-AKT pathway. Our findings unveil a novel METTL3/UBE2C/PI3K-AKT axis in RB pathogenesis, offering valuable insights into the mechanisms of RB tumorigenesis and providing novel therapeutic targets for RB.
Methods
Bioinformatic analysis
GSE97508 is an mRNA microarray from GEO datasets, which includes six RB samples and three normal retinal samples. GSE208143 also from GEO datasets is another mRNA microarray, which includes 27 RB samples and six normal retinal samples. Then, the upregulated genes in RB samples were screened from GSE97508 and GSE208143 with setting screened criteria as follows: adj.p < 0.01 and logFC >2. Finally, STRING (https://cn.string-db.org/) was applied to construct protein–protein interaction (PPI) networks for the upregulated genes.
Cell culture
Y79 (SNL-532) and WERI-RB-1 (SNL-470) were two RB cell lines from Sunncell company (China), whereas ARPE-19 (SNL-227) was a human retinal pigment epithelial cell line from Sunncell company. RB cells and ARPE-19 cells were cultured in the Roswell Park Memorial Institute-1640 medium (Sunncell) and Dulbecco’s modified Eagle’s medium (Sunncell), respectively, under 5% CO2 and 37°C. All media were added to 10% fetal bovine serum (Sunncell).
Cell transfection
Short hairpin RNA of UBE2C (sh-UBE2C) and sh-NC (negative control) was purchased from GenePharma (China), and the sequences of sh-UBE2C are shown in Supplementary Table S1. Full-length UBE2C and METTL3 were cloned to pcDNA3.1 by GenePharma for constructing UBE2C and METTL3 overexpression vectors, respectively. The empty vector of pcDNA3.1 was applied as negative control of overexpression vector (Vector). Cell transfection was performed by transfecting the vectors mentioned above to RB cells with the help of Lipofectamine 3000 (Invitrogen, USA).
Quantitative reverse transcription–polymerase chain reaction
Total RNA used for quantitative reverse transcription–polymerase chain reaction (qRT-PCR) was isolated by RNA-easy Isolation Reagent (Vazyme, China). Then, cDNA transcribed by HiScript II Q RT SuperMix was also provided by Vazyme. qRT-PCR was performed by PerfectStart Green qPCR SuperMix (TransGen Biotech, China) to detect the mRNA expression with glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as internal reference using 2-ΔΔCt method. Table 1 displays all the primer sequences.
The Primers Used for Quantitative Reverse Transcription–Polymerase Chain Reaction
Cell viability assessment
Cell counting kit-8 (CCK8, Vazyme, China) was purchased to assess cell viability. RB cells with 5 × 103 cells/well were seeded into 96-well plates for cell culture. After incubation for 0, 24, 48, and 72 h, the CCK8 solution (10 μL/well) was added to cells for another 2-h incubation. Finally, the absorbance at 450 nm was determined via microplate reader.
Cell proliferation detection
The SF594 5-ethynyl-2′-deoxyuridine (EdU) cell proliferation detection kit (CA1174, Solarbio, China) was used to detect RB cell proliferation. A total of 5000 cells/well of RB cells were added to a 96-well plate for incubation for 24 h. Then, 10 μM EdU solution was added to RB cells for 4 h, fixed by paraformaldehyde (4%) and permeabilized by Triton X-100 (0.5%). Next, 1× Click-iT and 1× DAPI were added to cells for incubating 30 min without light, respectively. The RB cells were observed by a fluorescence microscope.
Cell apoptotic rate measurement
The annexin V-FITC/PI kit (Beijing Bestech Biotechnology, China) was purchased to measure the RB cell apoptotic rate. Briefly, 5 × 105 RB cells were harvested, resuspended in binding buffer (200 μL), and stained with annexin V-FITC/PI (annexin V-FITC: PI = 1:2). After incubation for 20 min away from light, CaliburC5 flow cytometry (BD, USA) detected the RB cell apoptotic rate.
Western blotting
Radioimmunoprecipitation assay buffer (high) purchased from Solarbio (China) was added to cells for isolating total protein. After detecting protein concentration using the Bicinchoninic Acid Protein Assay Kit (Solarbio), 30 μg of protein were isolated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and delivered to polyvinylidene fluoride membranes. Then, the membranes were blocked by Western Blocking Buffer (Solarbio) and the primary antibodies were incubated, including Phospho-PI3K (p-PI3K, AP0854, ABclonal, China), PI3K (A4992, ABclonal), Phospho-AKT (p-AKT, AP1208, ABclonal), AKT (A17909, ABclonal), GAPDH (AC027, ABclonal), and UBE2C (A24740, ABclonal), overnight at 4°C. Next day, the membranes were incubated with the rabbit secondary antibody (ABclonal) and Enhanced Chemiluminescence (ECL) Enhanced Plus Kit (ABclonal) for protein detection.
Methylated RNA immunoprecipitation
The m6A methylated RNA immunoprecipitation (MeRIP) kit (Bes5203-2) provided by Bersinbio (China) was utilized to perform MeRIP assay. Total RNA (18 μg) isolated from RB cells was fragmented by fragmentation buffer for 6 min at 70°C. Then, the fragmented RNA was mixed with Protein A/G beads coated with 2 μL anti-m6A antibody or anti-IgG for a 1-h incubation. After purifying RNA using 75% ethyl alcohol, qRT-PCR was performed to detect UBE2C expression.
mRNA stability detection
mRNA stability was detected with the help of actinomycin D (53600ES08, YEASEN, China). 5 × 105 cells/well RB cells were added to 6-well plates to treat with 2 μg/mL actinomycin D for 0, 3, 6, and 9 h. After treatment, the cells were collected to isolate RNA for qRT-PCR to detect the level of UBE2C.
Statistical analysis
All data from the triple experiments were displayed as mean ± SD, and GraphPad Prism was applied to analyze the data. Student’s t-test and analysis of variance (ANOVA) were utilized to verify differences between two and several groups, respectively. A p value of <0.05 is indicative of significant difference.
Results
UBE2C is a key gene in RB
After setting screened criteria as adj.p < 0.01 and logFC >2, 5 common upregulated genes, including PCLAF, BIRC5, UBE2C, CCNB2, and NEK2, in RB samples were overlapped from two mRNA microarrays (GSE97508 and GSE208143) downloaded from GEO datasets (Fig. 1A). Then, these common upregulated genes were uploaded to STRING for PPI analysis. The results showed BIRC5 and UBE2C with more connection (Fig. 1B). By qRT-PCR analysis, UBE2C expression in RB cells was more significantly upregulated than BIRC5 expression in RB cells (Fig. 1C). Therefore, UBE2C was selected as the key gene in RB for further investigation in this study.

UBE2C is a key gene in retinoblastoma (RB).
UBE2C induces RB cell survival by regulating proliferation and apoptosis
To verify the function of UBE2C in RB, UBE2C overexpression/knockdown vectors were designed and transfected to two RB cell lines. qRT-PCR analysis showed that UBE2C protein expression in RB cells was upregulated more than twofold after UBE2C overexpression transfection, and downregulated >50% after UBE2C knockdown transfection (Fig. 2A). Western blotting further verified that UBE2C expression in RB cells was upregulated more than eightfold after UBE2C overexpression transfection, and downregulated by ∼70% after UBE2C knockdown transfection (Fig. 2B). After assessing RB cell viability using CCK8 assay, it could be observed that UBE2C overexpression enhanced cell viability, but UBE2C knockdown declined cell viability after transfection for 48 and 72 h (Fig. 2C). EdU detection found that UBE2C induced RB cell proliferation, but RB cell proliferation was inhibited after silencing UBE2C (Fig. 2D). The cell apoptotic rate detected by flow cytometry displayed that upregulating UBE2C reduced apoptotic rate, whereas downregulating UBE2C elevated apoptotic rate in RB cells (Fig. 2E). All results proved that UBE2C enhanced the ability of RB cell survival.
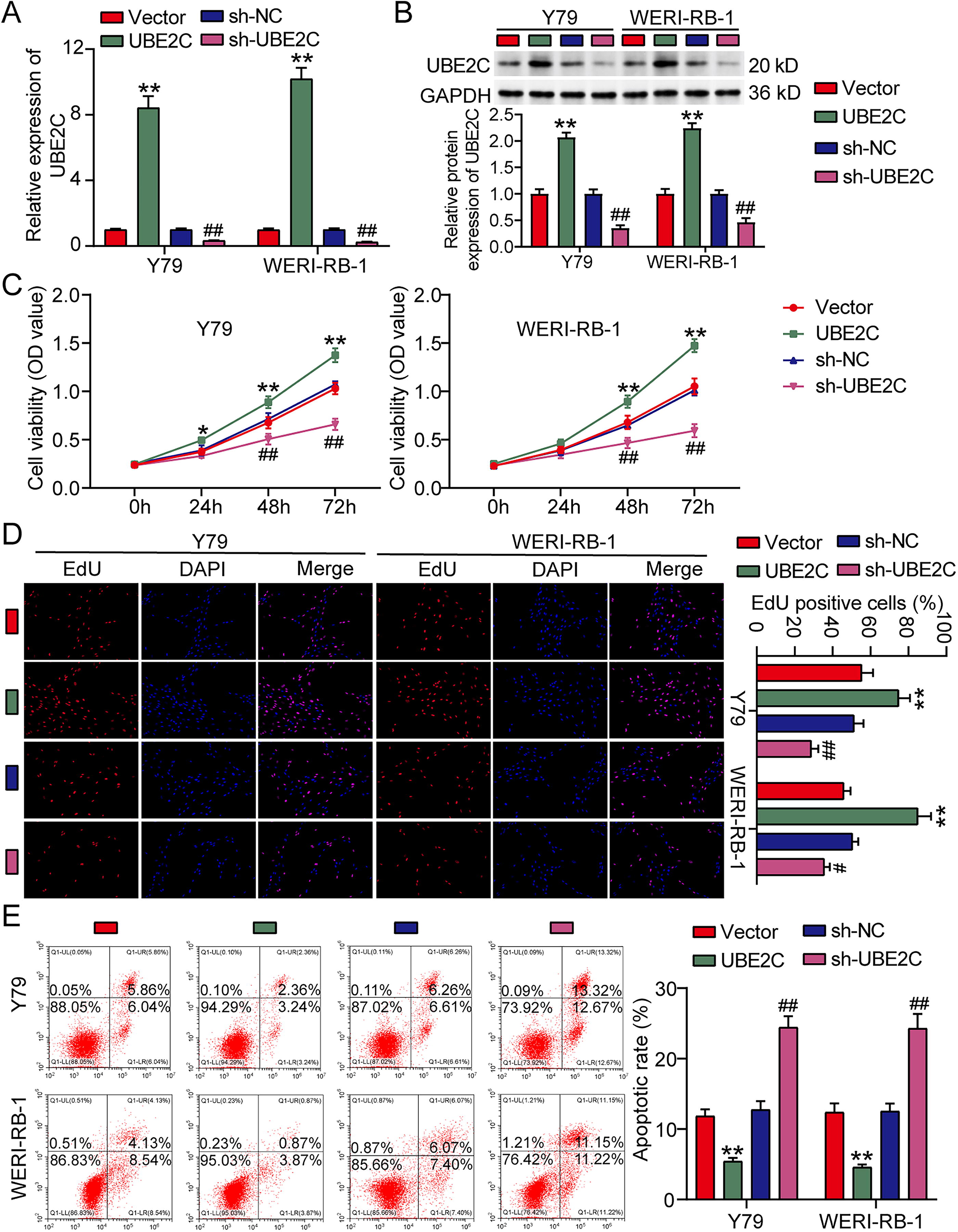
Ubiquitin-conjugating enzyme E2C (UBE2C) induces retinoblastoma (RB) cell survival by regulating proliferation and apoptosis.
UBE2C activates PI3K-AKT pathway
Since UBE2C was reported to regulate the PI3K-AKT pathway in osteosarcoma and acute myeloid leukemia (Huang et al., 2024; Wang et al., 2024), this study would like to confirm whether UBE2C could affect PI3K-AKT pathway in RB. Using Western blotting to detect PI3K-AKT pathway-associated proteins, it was found that UBE2C overexpression elevated the protein levels of phosphorylated PI3K and AKT, whereas UBE2C knockdown reduced the protein levels of phosphorylated PI3K and AKT (Fig. 3). These results indicated that UBE2C could activate PI3K-AKT pathway in RB cells.
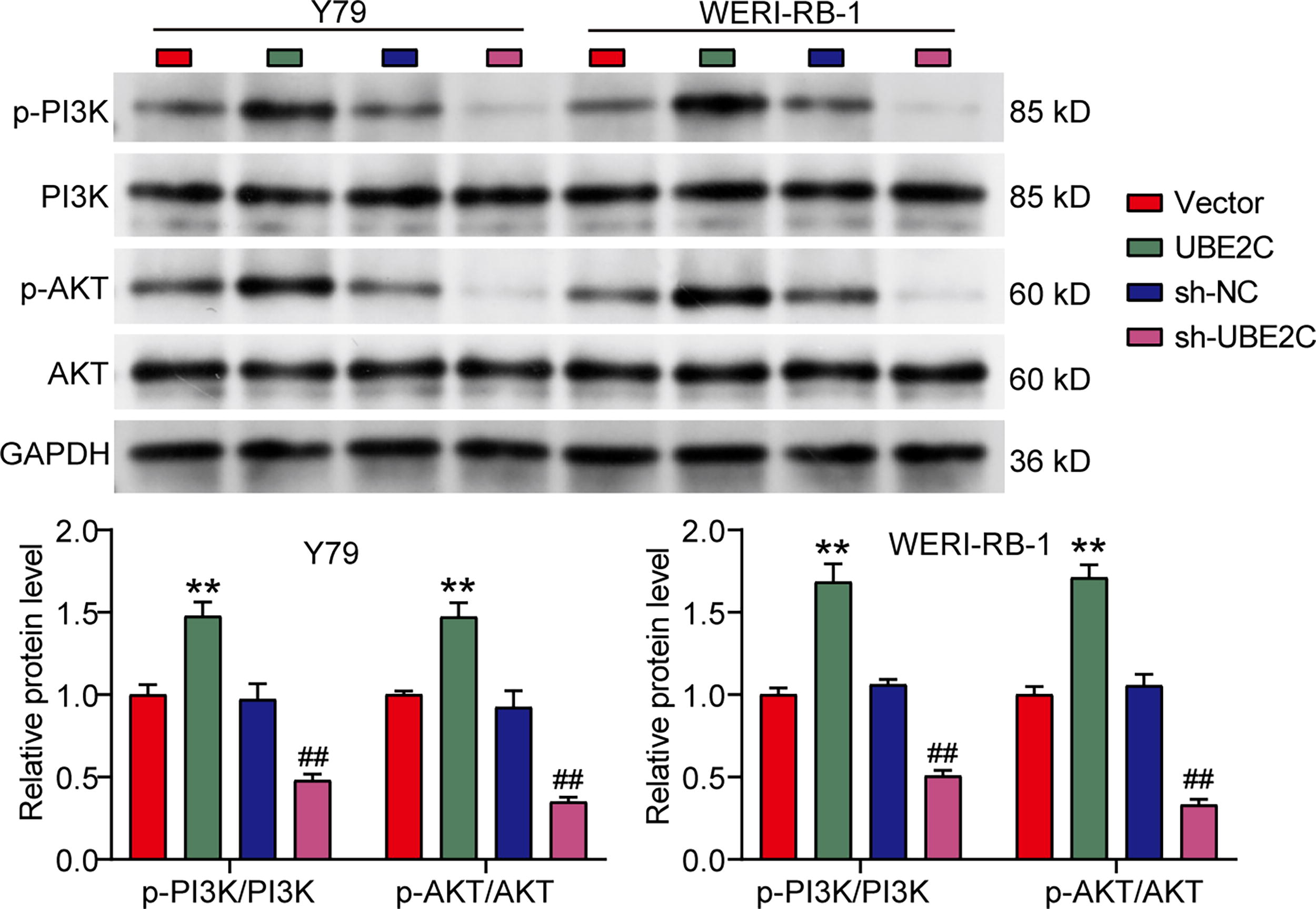
Ubiquitin-conjugating enzyme E2C (UBE2C) activates phosphoinositide 3-kinase-protein kinase B (PI3K-AKT) pathway. Western blotting detected the expression of PI3K-AKT pathway-related proteins in two retinoblastoma (RB) cell lines (Y79 and WERI-RB-1) transfected with UBE2C overexpression/knockdown vectors. UBE2C, UBE2C overexpression vector. sh-UBE2C, UBE2C knockdown vector. **p < 0.001 versus vector; ##p < 0.001 versus sh-NC.
METTL3 positively regulates UBE2C expression in RB via inducing m6A modification of UBE2C
To further explore the upstream regulator of UBE2C in RB cells, qRT-PCR was performed to assess UBE2C expression in RB cells transfected with overexpression vectors of common m6A writers, including METTL3, METTL14, KIAA1429, WTAP, ZC3H13, and RBM15. The results showed that only METTL3 overexpression increased UBE2C expression in RB cells (Supplementary Table S1). By qRT-PCR, METTL3 was upregulated approximately fourfold in RB cells (Fig. 4A). After transfecting METTL3 overexpression vector to RB cells, MeRIP data displayed that UBE2C m6A level was upregulated ∼10-fold (Fig. 4B). qRT-PCR analysis detected more than fourfold upregulation of UBE2C mRNA in RB cells transfected with METTL3 overexpression vector (Fig. 4C). Similarly, Western blotting measured approximately twofold upregulation of UBE2C protein in RB cells transfected with METTL3 overexpression vector (Fig. 4D). Moreover, METTL3 overexpression also enhanced the stability of UBE2C mRNA in RB cells (Fig. 4E). Taken together, METTL3 could enhance UBE2C expression in RB via promoting m6A modification of UBE2C.
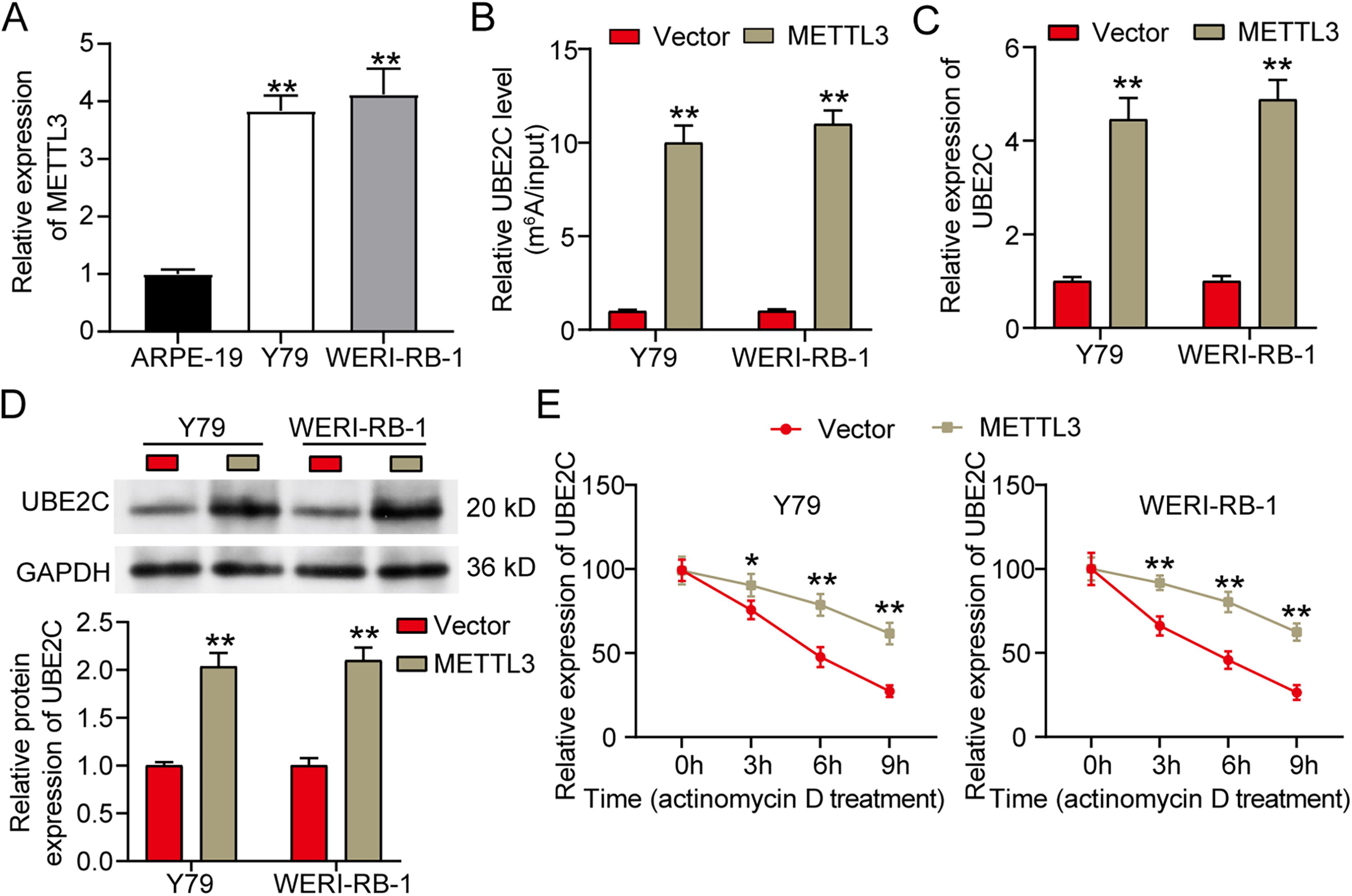
Methyltransferase-like 3 (METTL3) positively regulates ubiquitin-conjugating enzyme E2C (UBE2C) expression in retinoblastoma (RB) via inducing N6-methyladenosine (m6A) modification of UBE2C.
Upregulating METTL3 partially recovered the antitumor role of UBE2C downregulation in RB cells
To unravel the mechanism by which METTL3 regulated UBE2C to affect RB cells, METTL3 overexpression vector was transfected to UBE2C-silenced RB cells for cell functional detection. CCK8 assay showed that the inhibitory cell viability by sh-UBE2C was partially relieved after cotransfection of METTL3 overexpression vector (Fig. 5A). Similarly, cotransfection of METTL3 overexpression vector also could partly recover cell proliferation ability that was impaired by sh-UBE2C (Fig. 5B). As for cell apoptosis, the elevated apoptotic rate caused by sh-UBE2C was reduced by METTL3 overexpression (Fig. 5C). These results confirmed that upregulating METTL3 partially relieved the antitumor effect of UBE2C downregulation on RB cells.
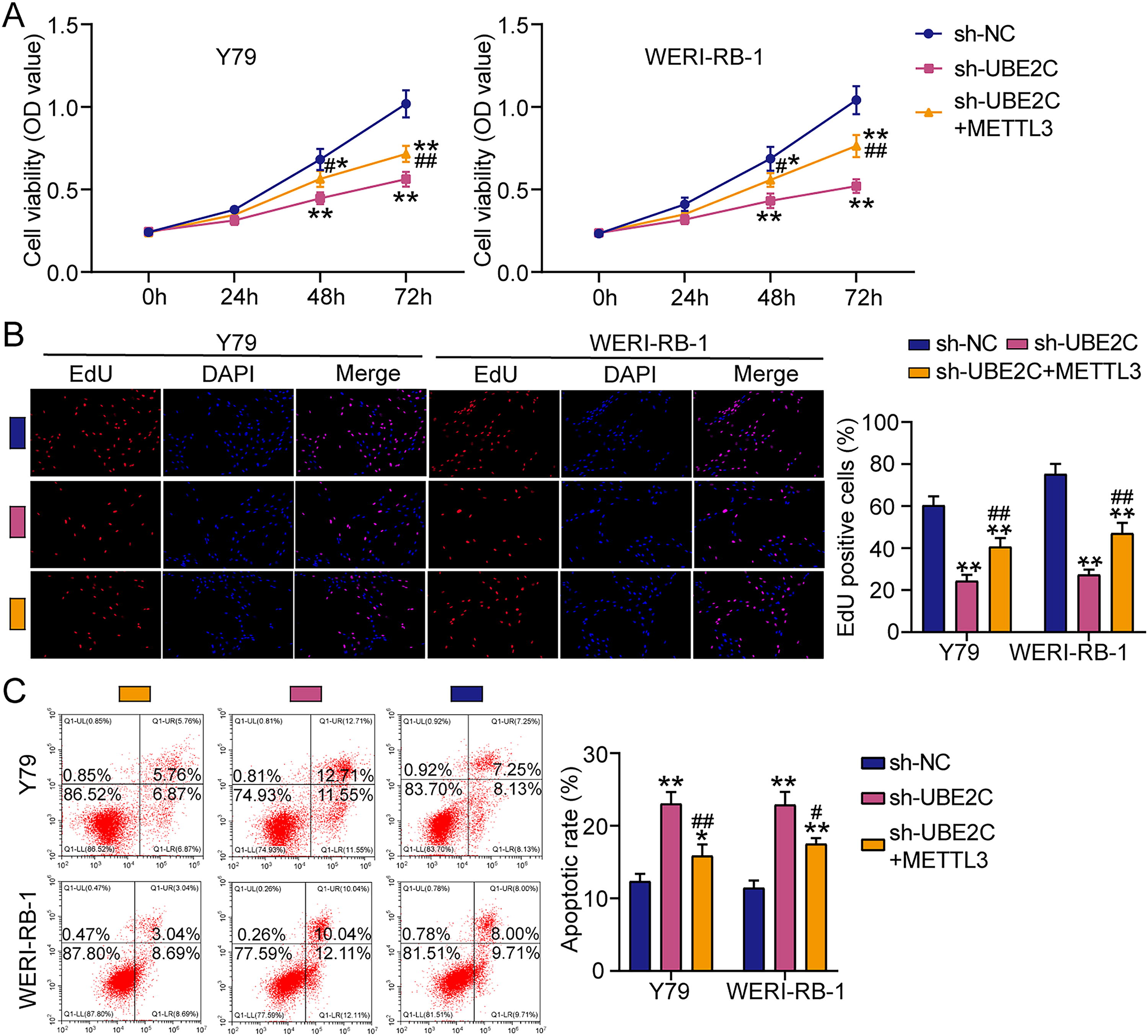
Methyltransferase-like 3 (METTL3) partially reversed the inhibitory effect of ubiquitin-conjugating enzyme E2C (UBE2C) knockdown on retinoblastoma (RB) cells.
Discussion
RB, a devastating pediatric ocular malignancy, remains poorly understood at the molecular level despite advances in clinical management. In this study, we identified that UBE2C exerted the oncogenic effects on RB progression through activation of the PI3K-AKT pathway. Importantly, METTL3-mediated UBE2C upregulation via m6A modification partially reversed the inhibitory effects of UBE2C knockdown, further supporting the oncogenic role of the METTL3/UBE2C axis in RB (Fig. 6). Our findings provide novel insights into the mechanisms driving RB progression and potential therapeutic targets.
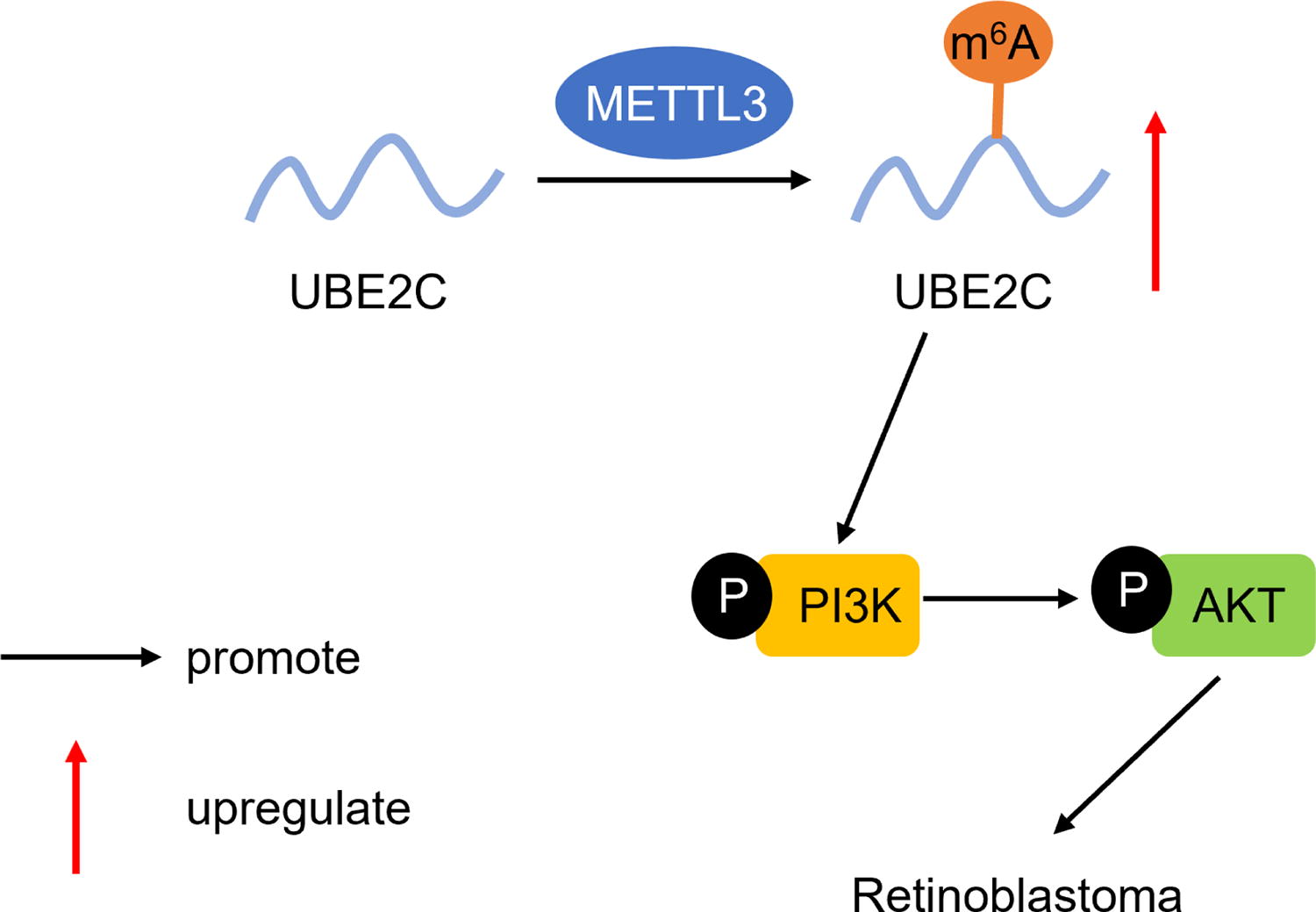
The mechanism diagram of methyltransferase-like 3 (METTL3)-mediated ubiquitin-conjugating enzyme E2C (UBE2C) N6-methyladenosine (m6A) modification to regulate phosphoinositide 3-kinase-protein kinase B (PI3K-AKT) pathway in retinoblastoma (RB).
Dysregulated UBE2C expression has been implicated in various malignancies. For example, UBE2C with high expression in cervical cancer could enhance cervical cancer cell malignancy in vitro and in vivo (Chiang et al., 2020). The upregulation of UBE2C was correlated with poor survival in patients with pancreatic cancer, and its overexpression promoted the malignant activity of pancreatic cancer cells (Cao et al., 2022). In endometrial cancer, elevated UBE2C expression displayed higher histological grade and shorter overall survival, as well as UBE2C could also enhance endometrial cancer cell proliferation and metastasis (Liu et al., 2020b). The previous studies seem to confirm the oncogenic role of UBE2C in cancers although no study revealed the role of UBE2C in RB. Our work is the first to establish UBE2C’s role in RB, showing that elevated UBE2C expression significantly enhances RB cell survival by promoting proliferation and inhibiting apoptosis. Our findings suggest that silencing UBE2C may serve as a promising strategy for inhibiting RB cell malignancy.
PI3K-AKT pathway as a key regulator was frequently hyperactivated in cancers through genetic alterations or upstream signaling aberrations (Ganesan et al., 2024; Rascio et al., 2021; Xie et al., 2025). Activation of PI3K-AKT signaling confers survival advantages to cancer cells by accelerating proliferation and cell cycle progression, inhibiting apoptosis, and enhancing metabolic reprogramming (He et al., 2021; Hoxhaj and Manning, 2020; Liu et al., 2020a). In RB, PI3K-AKT activation has been associated with tumor progression and antitumor drugs (Lv et al., 2022; Tang et al., 2022; Yin et al., 2024). Previous studies have suggested that UBE2C in cancers may contribute to cancer progression by activating PI3K-AKT pathway. For instance, UBE2C accelerated the proliferation of acute myeloid leukemia cells by activation of PI3K-AKT pathway (Wang et al., 2024). UBE2C in osteosarcoma was reported to activate the PI3K-AKT pathway to promote tumor progression (Huang et al., 2024). These studies imply a potential link between UBE2C and PI3K-AKT signaling in RB as well. Our study reveals that UBE2C activates PI3K-AKT pathway in RB cells, as evidenced by increased phosphorylation of PI3K and AKT proteins upon UBE2C overexpression. Mechanistically, UBE2C promoted PI3K-AKT activation to facilitate RB tumorigenesis. Given the critical role of PI3K-AKT signaling in RB pathogenesis, the UBE2C-PI3K-AKT axis represents a potential therapeutic target for RB treatment.
Another innovation of this study is the discovery that METTL3, a core m6A methyltransferase, regulates UBE2C expression via m6A modification in RB. The m6A epitranscriptomic machinery has emerged as a critical player in cancer progression, with METTL3 often acting as an oncogene or a suppressor by enhancing the expression of target mRNAs (Wang et al., 2020; Zhu et al., 2022). In RB, only Zhang H et al. revealed the upregulation of METTL3 in RB, and its overexpression accelerated RB progression by the PI3K-AKT pathway (Zhang et al., 2020). However, their study did not explore the downstream target genes of METTL3 in RB. This study innovatively found that METTL3 promotes UBE2C expression by enhancing its m6A modification, which increases UBE2C mRNA stability. In addition, this study demonstrated that METTL3 overexpression partly recovered the antitumor roles of UBE2C silence in RB cell survival. Our findings indicate that the METTL3-UBE2C axis may represent a promising therapeutic node for RB.
Our study has some limitations. While in vitro models provide mechanistic insights of METTL3/UBE2C/PI3K-AKT axis in RB, this study lacks the investigation in vivo by constructing a xenograft animal model. In addition, the clinical relevance of METTL3-UBE2C-PI3K-AKT signaling needs verification in RB tumor cohorts with matched outcome data by collecting RB clinical samples.
Conclusion
This study proposes a mechanism in which METTL3-mediated m6A modification upregulates UBE2C, which in turn activates PI3K-AKT signaling to drive RB progression. Our findings confirm novel insights into the mechanisms underlying RB tumorigenesis and indicate the METTL3/UBE2C/PI3K-AKT axis as a promising therapeutic target for RB treatment.
Footnotes
Ethics Approval
This study does not perform any studies involving animals or human participants.
Consent for Publication
The participants provided their consent for publication.
Availability of Data and Material
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
Author Disclosure Statement
No conflicts of interest were declared by the authors.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Table S1
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.