
Abstract
Abstract
Solubilization of sewage sludge by thermophilic bacteria isolated from a hot spring was investigated as a pretreatment for the methane fermentation process. The bacterial strain, designated strain MU3, was grown at an optimum pH of 7.5–8.0 and optimum temperature of 60°C. A phylogenetic analysis based on 16S rRNA gene sequences indicated that strain MU3 formed a distinct clade within the genus Anoxybacillus. Levels of sequence similarity of strain MU3 were the closest to Anoxybacillus rupiensis DSM 17127T. Among the strains of genus Anoxybacillus, only MU3 had an ability to hydrolyze casein. Casein hydrolysis activity of thermophilic extracellular protease produced by the strain MU3 showed an optimum temperature of 70°C. We attempted the application of strain MU3 culture into the sewage sludge, which was treated at 80°C for solubilization. Unfortunately, the isolate showed a decrease in the colony formation unit into the heat-treated sewage sludge. In batch experiments, solubilization of sewage sludge was enhanced by addition of the culture. Solubilization rates of sewage sludge supplemented with and without the culture were 32.0% and 18.8% after 67 h of incubation, respectively. The effect of the number of doses (one or two) of the culture on solubilization of the sewage sludge was investigated. Solubilization of a twofold dose was significantly enhanced as compared to a single dose after 45 h. Therefore, the solubilization of sewage sludge may be enhanced by thermophilic protease secreted by stain MU3.
Introduction
In production of biogas, methane fermentation is a useful technology capable of converting organic wastes such as sewage sludge to methane gas and providing energy recovery. Methanogenesis is formation of methane from high molecular organic substances by microorganisms known as methanogens. In the methane fermentation process, the organic substances in the sludge are hydrolyzed by enzymes secreted by the first group of microorganisms. The monosaccharides and amino acids produced by microbial degradation of the organic substances are subsequently converted to fatty acids, hydrogen, and acetic acid. In the second stage, the fatty acids, such as propionic and butyric acids, are converted to hydrogen, carbon dioxide, and acetic acid by hydrogen-producing acetogenic bacteria (Zeikus, 1980; McInerney et al., 1981). Finally, in the third stage, the acetate is converted to methane and carbon dioxide by methanogens (Thauer et al., 1977; Wolfe, 1985). Therefore, the initial solubilization of sludge, microbial degradation, is an important process in the methane fermentation.
A thermophilic bacterial strain, designated strain MU3, which produces extracellular thermophilic protease, was isolated from a hot spring located in Beppu, Oita, Japan. Strain MU3 is a motile Gram-positive rod bacterium, and is a moderate thermophile with an optimum temperature for growth at 60°C. Strain MU3 produced large amounts of extracellular thermophilic protease (Fig. 1). To determine whether the solubilization of sewage sludge in methane fermentation process is promoted by strain MU3, we examined the effect of strain MU3 culture on the solubilization of heat-treated sewage at 80°C. This temperature is selected to distil away ammonia, which is an inhibitor of methane fermentation.
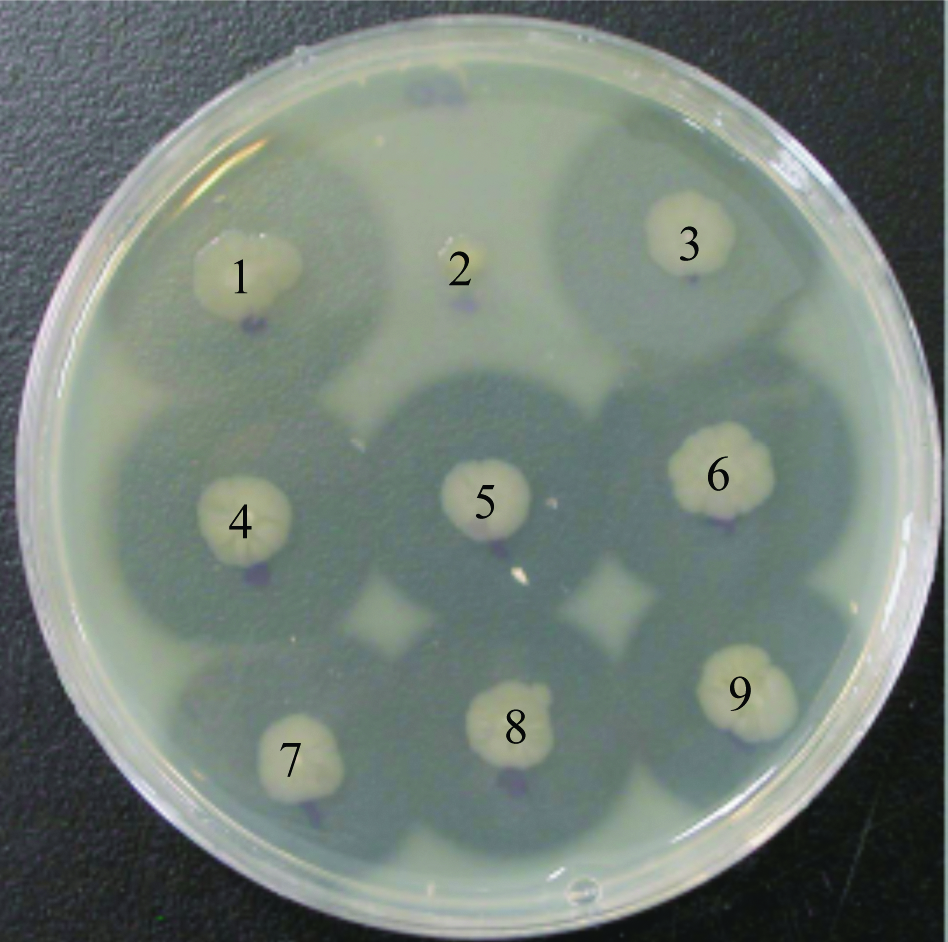
Screening of thermophiles producing extracellular protease. After accumulation culture with nutrient medium containing 0.3% skim milk, nine thermophiles producing extracellular protease were picked up. The producing proteases were observed in eight thermophiles, except for no. 2.
Experimental Section
Bacterial strain
A thermophilic bacterium was isolated from sediments collected at a hot spring in Beppu-city, Oita, Japan. The water temperature of the hot spring was about 90°C. The sediments were immediately spread onto ISP-2 (Becton Dickinson and Co.) containing 0.3% skim milk (Becton Dickinson and Co.) to obtain thermophiles that produce extracellular thermophilic protease. The secreting quantity of protease was determined by the size of the skim milk-halo-zone lysed (Fig. 1). Since eight thermophiles that produced protease were identified or very similar in morphology, strain no. 3, we named it MU3, was selected as a representative of thermophiles produced thermophilic protease.
The almost complete 16S rDNA sequences of strain MU3 were directly determined following polymerase chain reaction (PCR) amplification. A fragment of the 16S rDNA (corresponding to positions 9-1510 in the Escherichia coli numbering system) was amplified by PCR as described previously (Nakashima et al., 2005). The reaction involved 35 cycles of denaturation at 98°C for 1 min, annealing at 55°C for 15 s, and extension at 72°C for 1 min. PCR products were purified using a Microcon Spincolumn (Millipore Co.). Both strands of purified fragments were sequenced using an ABI Prism BigDye Terminator v3.1 kit (Applied Biosystems) under conditions recommended by the manufacturer as well as an automated sequence analyzer (model 3100; Applied Biosystems). The 16S rDNA sequences of strain MU3 were compared with available 16S rDNA sequences from GenBank/DDBJ/EMBL using the BLAST program (Altschul et al., 1997). Phylogenetic trees were created by the neighbor-joining method (Saitou and Nei, 1987) using the CLUSTAL W program (Thompson et al., 1994). The topological support of the internal branches from the resulting trees was evaluated by bootstrap analysis (Felsenstein, 1985) of the neighbor-joining method based on 1,000 replications.
Thermophilic protease
Strain MU3 was grown in 1 L of ISP-1 (Becton Dickinson and Co.) at 60°C for 1 day. The bacterial cells were separated by centrifugation (8,000 rpm, 15 min), and the supernatant was fractionated by precipitation with 90% saturation of ammonium sulfate. The protein was resolved in 50 mM sodium phosphate buffer, pH 7.0, and dialyzed against several changes of the same buffer. The dialyzed protein was subjected to Sephadex G-200 gel chromatography (Pharmacia) to estimate molecular weight. The column (2.5 × 98 cm) was eluted with the same buffer as described above.
The protease activity was determined as follows: the thermophilic protease fraction (0.2 mL) was incubated with 2 mL of 1% casein solution in 50 mM sodium phosphate buffer (pH 7.0) at 60°C for 20 min. The reaction was stopped by addition of 3 mL of 5% trichloroacetic acid. After 10 min at ambient temperature, the sample was filtrated with filter paper (Advantec No.5). The resulting free amino acids and peptides liberated in the filtrate solution were determined by absorbance at 280 nm. The pH stability of the protease was performed by incubating the enzyme with buffers at various pH values (pH 5–6 in 0.2 M acetate buffer; pH 7–8 in 0.2 M phosphate buffer; pH 8–10 in 0.2 M glycine buffer) at 4°C overnight. The effect of pH on the activity was measured using 1% casein as a substrate at different pH values as described above. The thermal stability of the protease was tested by incubation of the enzyme at 60°C–100°C.
Addition of strain MU3 culture in sewage sludge
The sewage sludge was obtained from the Nagasaki municipal government. Five hundred milliliters of sewage sludge was dispensed into a 1,000 mL separable flask. Meanwhile, strain MU3 was cultured with 100 mL of ISP-1 medium in a 500 mL Erlenmeyer flask, the cultivation being carried out in a rotary shaker (120 rpm) at 60°C overnight. After heating of the sewage sludge at 80°C, 10 mL of the cultured broth (5 × 105 cells/mL) was added to the separable flask. Five milliliters of the heat-treated sewage sludge was taken by the batch method at certain intervals. Sewage sludge without strain MU3 culture was used as a negative control.
Number of doses of strain MU3 culture in sewage sludge
After heating of the sewage sludge at 80°C, 10 mL of the cultured broth (5 × 105 cells/mL) was added to the separable flask at 0 time (single dose) or 0 time and 45 h (twofold dose). Five milliliters of the heat-treated sewage sludge was taken by the batch method at certain intervals. In addition, the heat-treated sewage sludge was exchanged with 125 mL of fresh sewage sludge at 45, 50, 64, and 77 h to be consistent with an actual plant.
Microbial lawn analysis and solubility of sewage sludge
The specimens taken from the solubilization system were diluted to 10−3 with sterile saline. The original and diluted samples were spread to ISP-2 medium supplemented with 0.3% skim milk. The bacteria grown at 60°C were measured as colony forming units (CFU). Total (T) and soluble (S) chemical oxygen demand (CODcr) were determined by the usual methods (APHA/AWWA/WEF, 1998). The solubilization rate of sewage sludge was calculated from the following equation:
Statistical analysis
Data are presented as the mean ± standard deviation of triplicate measurements. Individual groups were compared with appropriate controls using two-factor repeated measure analysis of variance.
Results
Characterization of strain MU3
The strain MU3 was observed to produce a large amount of extracellular thermostable protease on ISP-2 medium supplemented with 0.3% skim milk (Fig. 1). Light and electron microscopic observation revealed that cells of the isolate were Gram-positive, endospore-forming rods, and motility by means of peritrichous flagella (Fig. 2). This isolate grew well under aerobic conditions on ISP-2, but not anaerobic conditions. Growth of strain MU3 had an optimum pH of 7.5–8.0 and optimum temperature of 60°C (Table 1). The almost complete (1,515 bp) sequence of the 16S rDNA gene of strain MU3 was aligned and compared to the sequences of related bacteria. A homology search of their 16S rDNA sequences in the GenBank database indicated that the rDNA sequences from the isolate belongs to the genus Anoxybacillus. The sequences were aligned with each other using Clustal W along with reference strains (Fig. 3). The levels of sequence similarity of strain MU3 were closest to Anoxybacillus rupiensis DSM 17127T (Derekova et al., 2007).
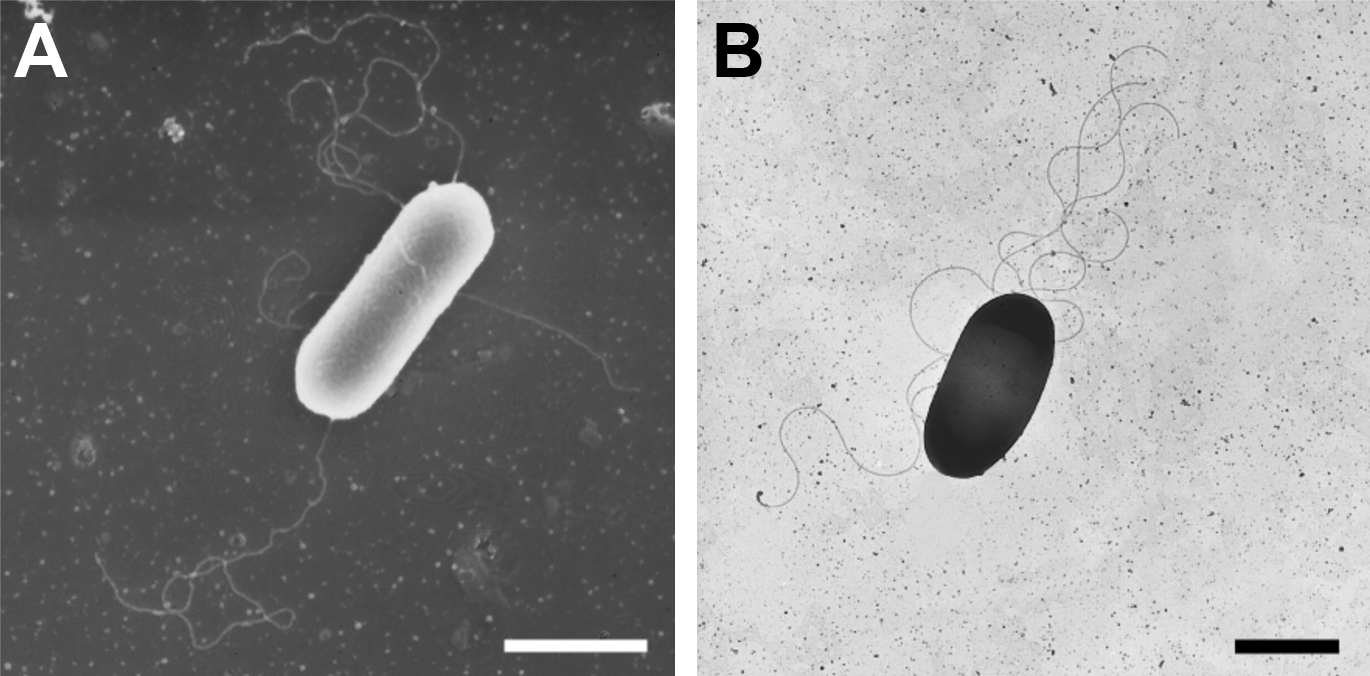
Scanning and transmission electron microscopy.
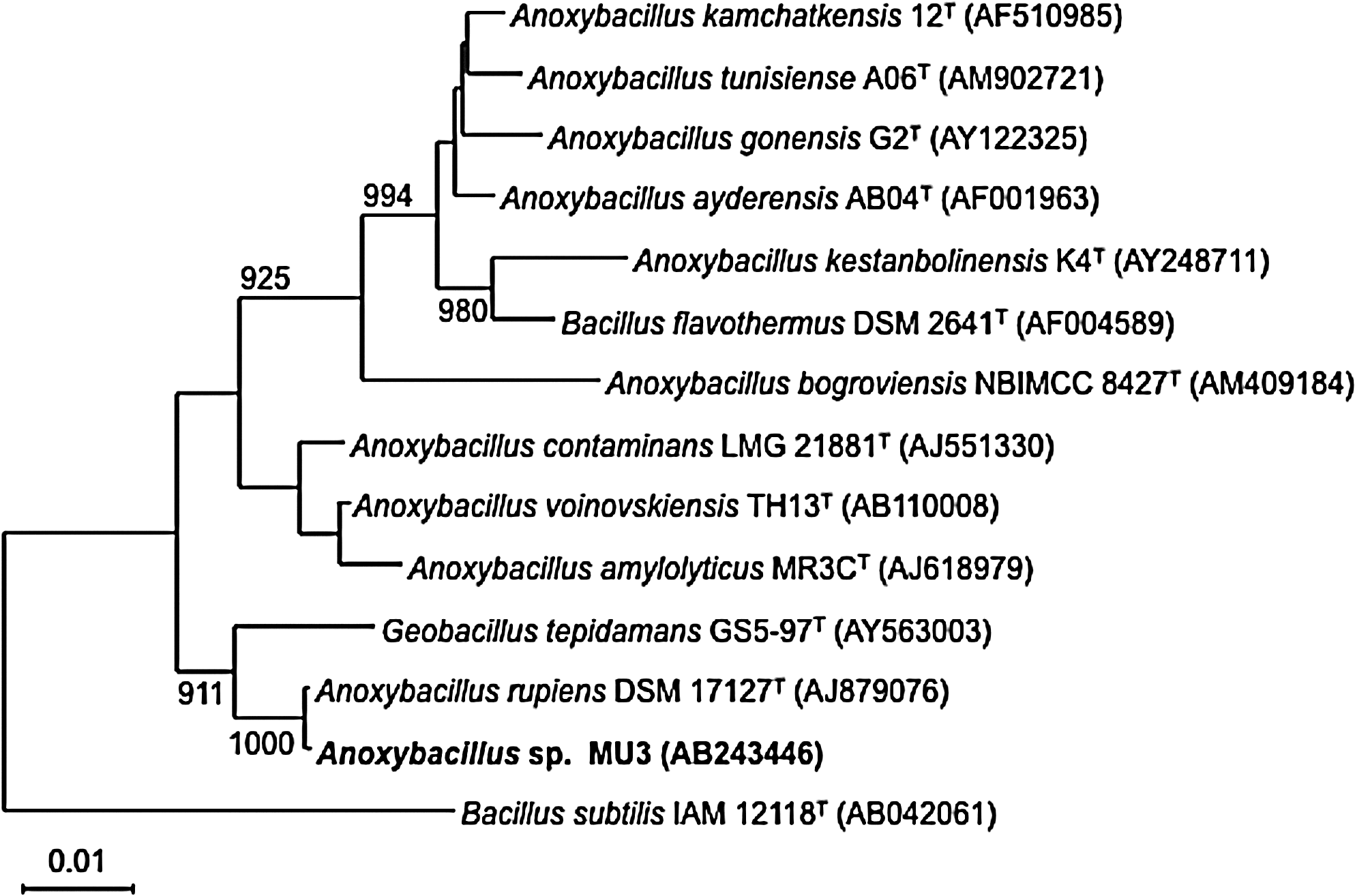
Neighbor-joining tree based on almost complete 16S rRNA gene sequences showing relationships between strain MU3 and members of the Anoxybacillus and Geobacillus sp. Bacillus subtilis IAM 12118T (AB042061) were used as the outgroup. Percentage numbers indicate the levels of bootstrap values for the branching point based on neighbor-joining analyses of 1,000 resampled data sets. Scale bar, 0.01 nucleotide substitutions per position.
Properties of thermophilic protease
The extracellular protease produced by strain MU3 was a thermophilic protease with a molecular weight of about 57,000 (data not shown). The thermophilic protease hydrolysed casein at a range of 50°C–90°C and pH 5–10, with an optimum temperature of 60°C and an optimum pH of 6 (acetate buffer) and 8 (glycine buffer) (Table 1).
Change of total bacterial numbers and solubilization rate in the sewage sludge with and without strain MU3 culture
The number of bacteria that appeared on the ISP-2 medium supplemented with 0.3% skim milk was measured. As shown in Fig. 4, the CFU in the heat-treated sewage sludge without strain MU3 showed a marked increase at 15 h and then decreased. The predominant colonies at 15 h showed nonsolubilization of skim milk. On the other hand, the CFU in the sewage sludge with strain MU3 showed a marked increase at 38 h and then decreased. Several different types of colonies were observed on the ISP-2 medium after 38 h of incubation, and these predominant colonies showed solubilization of skim milk. However, the morphology of predominant colonies was different from that of stain MU3.
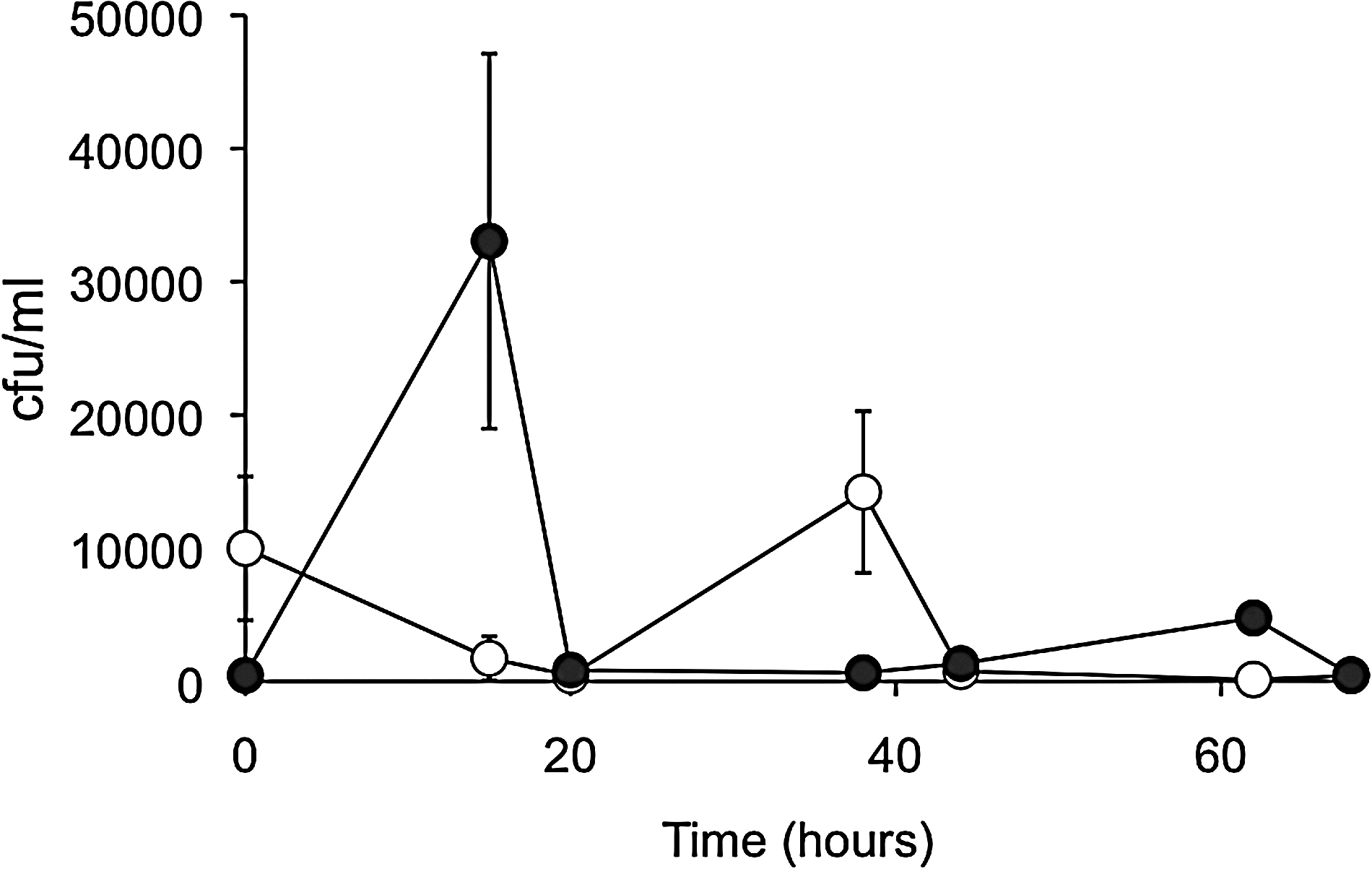
Fluctuation of number of total bacterial colony into heat-treated sewage sludge supplement with and without strain MU3 cultured broth. Colony forming unit in the sewage sludge with (○) and without (•) strain MU3 cultured broth.
The solubilization rates of sewage sludge with and without strain MU3 culture increased in a time-dependent manner (Fig. 5). However, the sewage sludge with strain MU3 culture showed a significantly higher solubilization rate than that without the culture (Table 2).
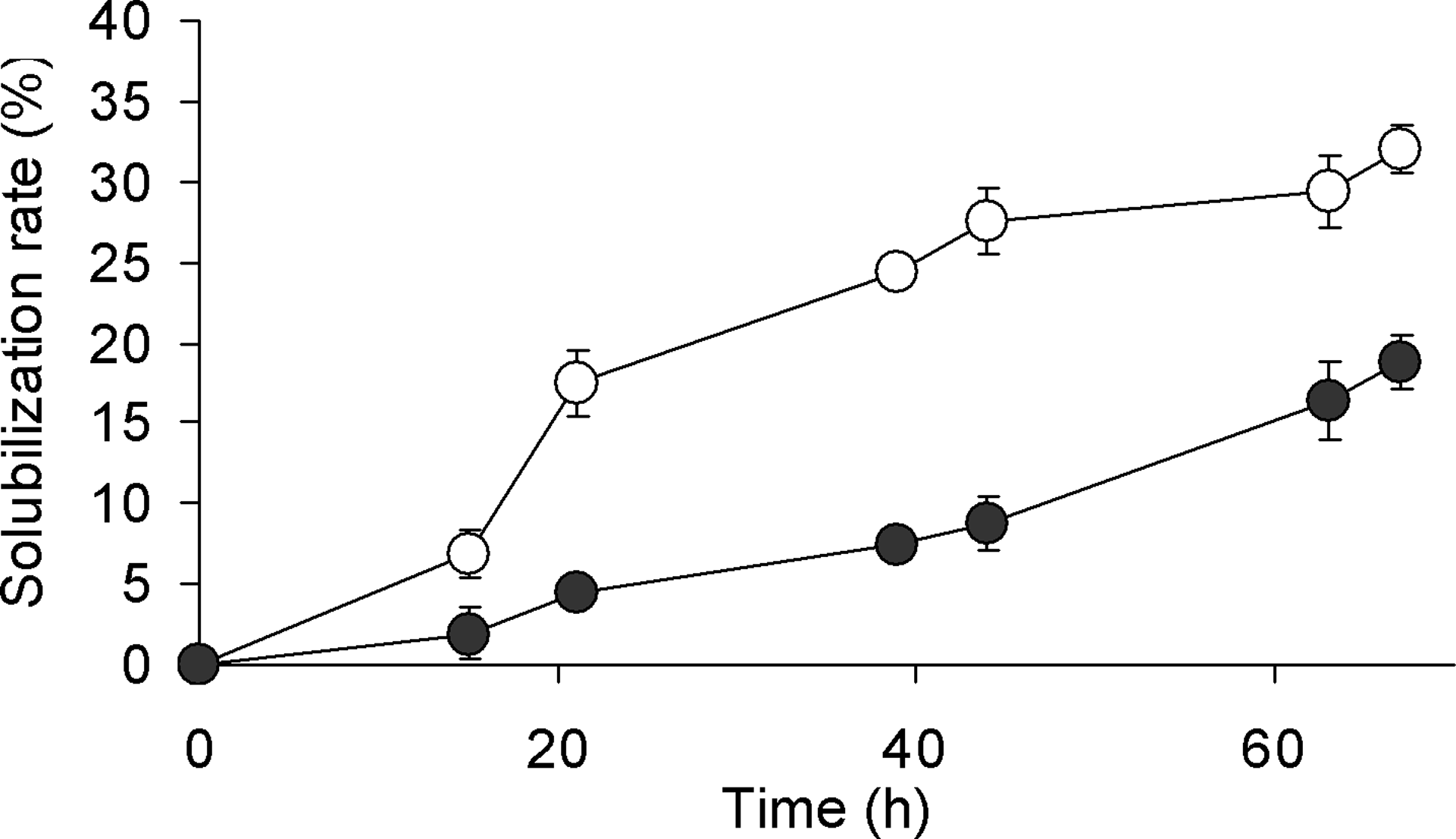
Solubilization rate of the heat-treated sewage sludge with (○) and without (•) strain MU3 culture. Experiments were performed in triplicate. Error bars represent standard deviation (SD).
SS, sum of squares; df, degree of freedom; MS, Mean-square.
Change in total bacterial number and solubilization rate in the sewage sludge receiving single and twofold doses of stain MU3 culture
The number of bacteria grown from the heat-treated sewage sludge was measured. As shown in Fig. 6, the CFU showed almost no change at 45 h in the sewage sludge subjected to single and twofold doses. Strain MU3 was observed as the predominant colony at the second dose (45 h) (Fig. 6A). However, the isolate rapidly decreased, and the colonies subsequently appearing at 48 h were morphologically different from strain MU3 (Fig. 6C). The colony number in the twofold dose was more than that in the single dose at 48 h (Fig. 6).
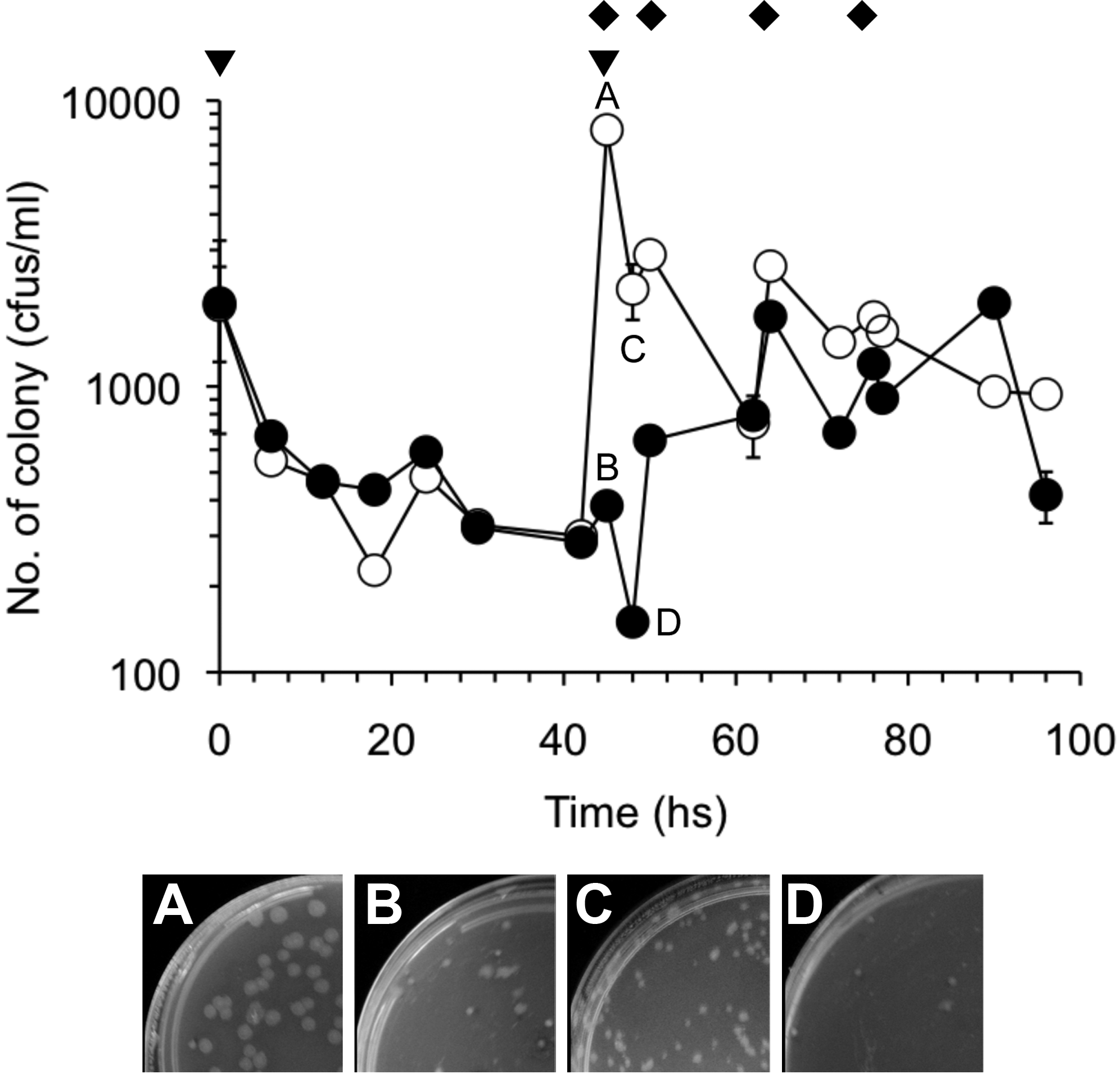
Fluctuation of number of total bacterial colony into heat-treated sewage sludge in single and twice doses of strain MU3 cultured broth. Colony forming unit in sewage sludge of single dose (•) and twice dose (○) of strain MU3 culture broth. Experiments were performed in duplication. Error bars represent SD.
The solubilization rates in CODcr analysis also showed almost no difference between the single and twofold dose at 45 h. Although the solubilization rates were temporarily reduced by the exchange of the sewage sludge, these values recovered in a time-dependent manner. The solubilization rates of twofold dose showed a significantly higher solubilization rate than those of single dose after 45 h (Fig. 7 and Table 3).
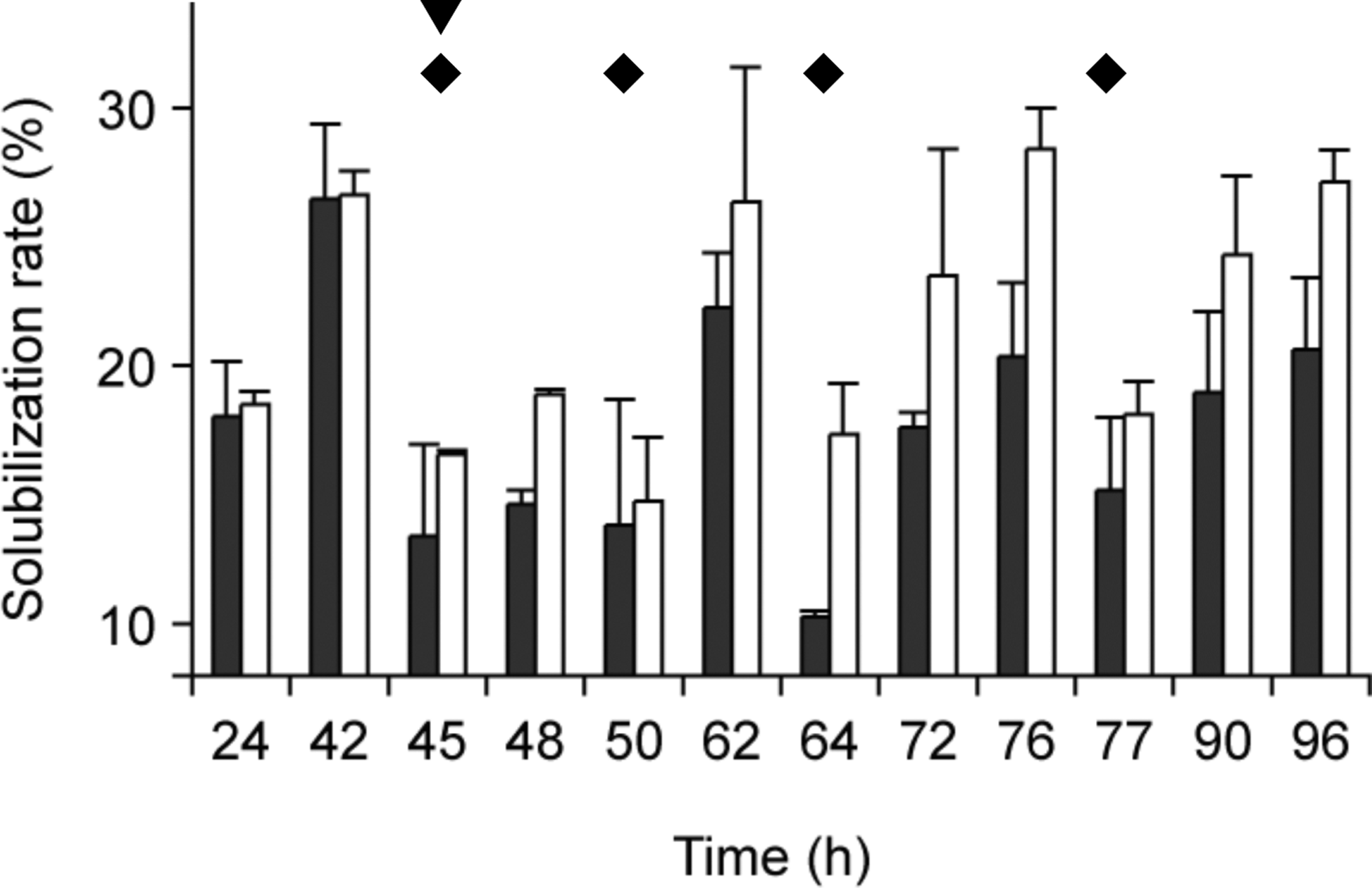
Solubilization rate of heat-treated sewage sludge in single dose (▪) and twice dose (□) of strain MU3 culture. ▾, addition of strain MU3 culture to sewage sludge. ♦, change of sewage sludge (125 mL). Experiments were performed in duplication. Error bars represent SD.
Discussion
Many authors have reported methods for the solubilization process (Häner et al., 1994; Juteau, 2006; Tsubota et al., 2007). Tsubota et al. reported that the hyper-thermophilic solubilization system (80°C) had several advantages such as higher degradation rate of the sewage sludge and exclusion of ammonia, which is an inhibitor of methane fermentation (Tsubota et al., 2007). In addition, the strategy of enzymatic hydrolysis of sewage sludge resulted in enhanced digestion and increased biogas production relative to other processes, as well as a better energy balance (Hasegawa et al., 2000; Biosolids Technology Fact Sheet, 2006). In fact, the efficacy of thermostable enzymes has been investigated previously in the thermophilic aerobic condition, resulting in a sludge solubilization rate of about 40% (Hasegawa et al., 2000). Therefore, the application of thermophilic bacteria that secrete extracellular enzymes to the sewage sludge solubilization can be fully exploited as a pretreatment strategy. This approach permits more complete digestion of sludge, with concomitant higher methane yields.
Strain MU3, which produces extracellular thermophilic protease, was isolated from a hot spring. The isolate belonged to the genus Anoxybacillus. The 16S rDNA sequence of strain MU3 showed a 99.6% similarity to A. rupiensis DSM 17127T (Derekova et al., 2007) and a 96.5% and 95.1% similarity to A. contaminans LMG 21881T (De Clerck et al., 2004) and A. voinovskiensis TH13T (Yumoto et al., 2004), respectively. Although these type strains were incapable of hydrolyzing casein (De Clerck et al., 2004; Yumoto et al., 2004; Derekova et al., 2007), the thermophilic protease produced by strain MU3 showed casein hydrolysis activity at 50°C–90°C, with an optimum temperature of 70°C and pH 5–10, with an optimum pH of 6 (acetate buffer) or 8 (glycine buffer) (Table 1). Therefore, it was considered that strain MU3 may be suitable for the application of the sludge solubilization process among genus Anoxybacillus. In addition, the characteristic property of protease was a molecular weight of about 57,000 and stability at 90°C for over 8 h (data not shown). Since the used sewage sludge showed pH 6–7, these properties of thermophilic protease were consistent with the solubilization conditions of sewage sludge.
The number of CFU differed in the two experiments. This was considered attributable to be the difference of the provided sewage sludge. When strain MU3 culture was added to the sewage sludge, the isolate unfortunately disappeared in a short period. Although bacterial colonies that subsequently appeared at 38 h in the additional experiment or at 48 h in the twofold dose experiment were morphologically different from strain MU3, the bacteria showed the solubilization of skim milk. These bacteria could not be observed in the heat-treated sewage sludge without strain MU3 culture or single dose. The bacterial flora in the heat-treated sewage sludge may be affected by the addition of strain MU3 culture. In addition, the solubilization of sewage sludge was enhanced by the addition of strain MU3 culture (Figs. 5 and 7). Therefore, it is likely that the heat-treated sewage sludge is solubilized by the extracellular thermophilic protease produced by stain MU3.
As shown in Fig. 8, we constructed a methane fermentation plant (5 m3) equipped with hyper-thermophilic solubilization phase (1 m3, 80°C) in Nagasaki Prefecture, Japan. We will employ the addition of strain MU3 culture (10 L) to the hyper-thermophilic solubilization phase (1 m3). Strain MU3 secretes extracellular protease that is heat-stable and very useful for solubilization of heat-treated sewage sludge.
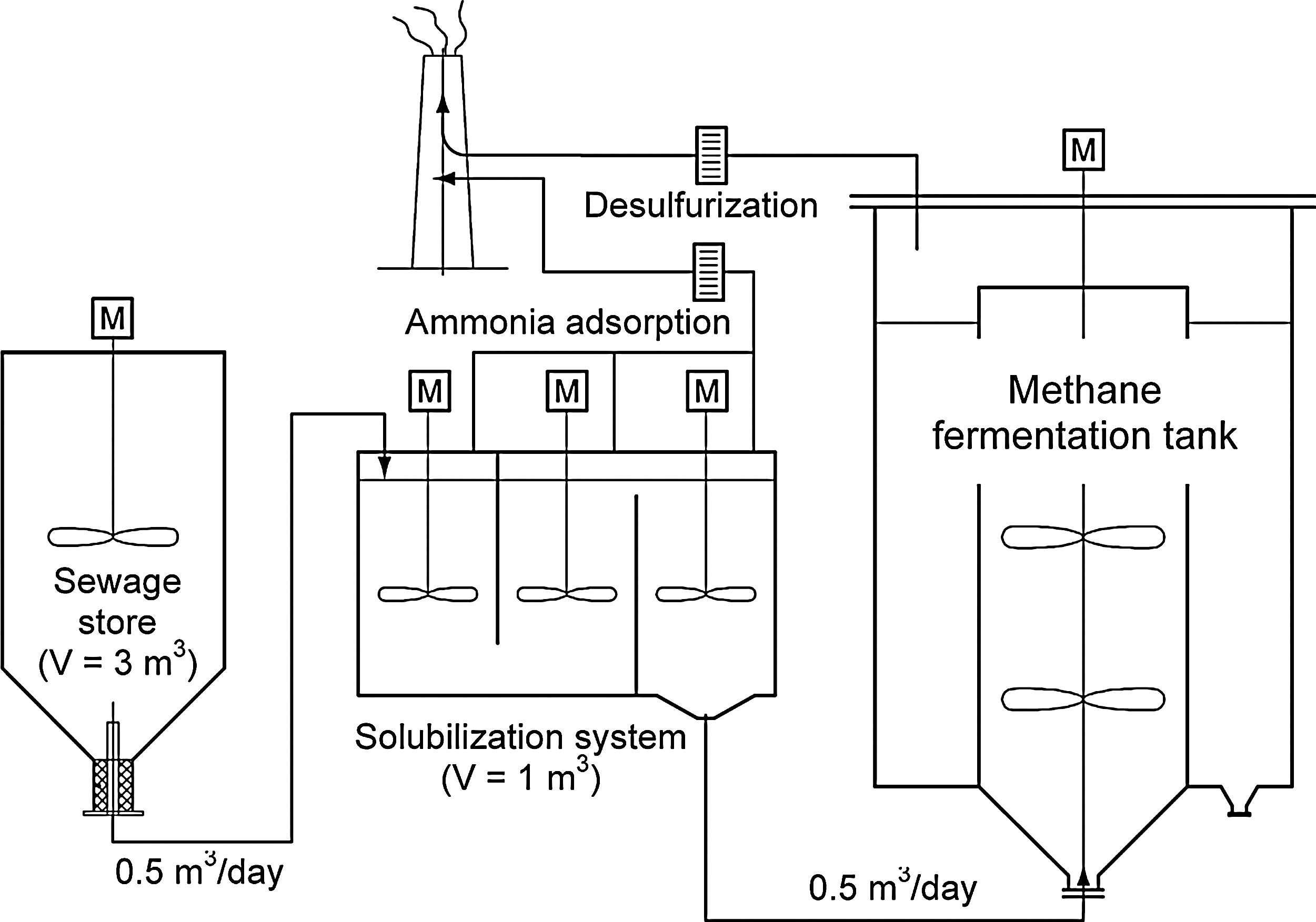
Illustration of methane fermentation plant equipped with hyper-thermophilic solubilization phase.
Footnotes
Acknowledgment
This study was supported in part by a research grant (2009–2010) from the Kyushu Bureau of Economy, Trade and Industry.
Author Disclosure Statement
No competing financial interests exist.