
Abstract
Abstract
Two thermophilic bacterial strains, Bacillus subtilis BUM (BUM) and Mycobacterium vanbaalenii BU42 (BU42), were tested for their potential in biodegradation of benzo[a]pyrene. Neither BUM nor BU42 utilized benzo[a]pyrene as the sole substrate under thermophilic condition. In the presence of 50, 250, and 500 mg L−1 phenanthrene, the biodegradation of benzo[a]pyrene by BUM occurred and removals in 30 days were 14.8%, 38.8%, and 63.6%, respectively. This is the first report on the biodegradation of benzo[a]pyrene by isolated thermophilic microorganism. The BUM strain was further tested for its ability in a soil composting system. Within a composting period of 42 days, removal of benzo[a]pyrene in the absence of BUM was 30.3%. Treatment with the inoculation of BUM significantly increased the removal of benzo[a]pyrene to 52.2%. Maximum zero-order degradation rates of benzo[a]pyrene by BUM in aqueous biodegradation experiment and soil composting system were 12.3 mg L−1 day−1 and 9.7 mg kg−1 day−1, respectively, which were significantly greater than most of the reported degradation rates by mesophiles. Experimental results affirmed that the strain BUM can effectively degrade benzo[a]pyrene under thermophilic condition.
Introduction
Degradation of benzo[a]pyrene by microorganisms is often thought to occur through cometabolic processes (Kanaly et al., 2000). To date, the reports on the roles of cosubstrates, for example, low-molecular-weight PAHs, in the biodegradation of benzo[a]pyrene are contradictory. The presence of a cosubstrate might impact the biodegradation of high-molecular-weight PAHs by competitive inhibition and by the toxicity of cosubstrate to microorganisms. The biodegradation of benzo[a]pyrene by induced cells of Mycobacterium sp. RGJII-135 was observed to be inhibited by PAHs with higher bioavailability, that is, benzo[a]anthracene, pyrene, and anthracene (McLellan et al., 2002). The competitive inhibition of naphthalene was also found during the degradation of phenanthrene by Pseudomonas sp. (Stringfellow and Aitken, 1995). Another mechanism for the inhibition of the biodegradation of PAHs by cosubstrate was reported by Shuttleworth and Cerniglia (1996), who found that the biodegradation of phenanthrene was inhibited by naphthalene because of the its toxicity to the PAH-degrading microbial strains. On the other hand, the presence of low-molecular-weight PAHs may facilitate the degradation of PAHs with high molecular weight. In the presence of 250 mg L−1 pyrene, a mixed culture of bacterium and fungus degraded 58.1% of 50 mg L−1 of benzo[a]pyrene in 56 days, whereas only 25.5% of benzo[a]pyrene was degraded when it was used as a sole carbon source (Boonchan et al., 2000). Furthermore, benzo[a]pyrene degradation by the resting cells of Mycobacterium sp. RGJII-135 can be induced by the presence of other PAHs, among which pyrene and phenanthrene were the most potent inducers (Cerniglia et al., 1992). The synergistic biodegradation of low-molecular-weight and high-molecular-weight PAHs has also been reported by other researchers (Kanaly et al., 1997; Bauer and Capone, 1998) and the responsible mechanisms may include increase in microbial growth by preferential utilization of the high-affinity substrates or induction of the PAH-degrading pathway (Cerniglia, 1993).
The degradation of benzo[a]pyrene under mesophilic condition was largely restricted by its low bioavailability due to its low solubility and mass transfer rate. The elevated temperature can increase the solubility and mass transfer rates of PAHs (Viamajala et al., 2007). Under thermophilic condition (60°C), high removal rates of PAH compounds, that is, phenanthrene, fluorine, and fluoranthene, were observed (Viamajala et al., 2007). To date, the feasibility of biodegradation of high-molecular-weight PAHs, for example, benzo[a]pyrene, by isolated strain under thermophilic condition has not been reported and this is the first report to our knowledge.
In the present study, we investigated the potential of two isolated thermophilic bacterial strains, B. subtilis BUM (BUM) and Mycobacterium vanbaalenii BU42 (BU42), on the biodegradation of benzo[a]pyrene under thermophilic conditions. Particular attention was paid to the effects of different cosubstrates on the benzo[a]pyrene biodegradation.
Experimental
Microorganisms
BUM and BU42 were originally isolated in our laboratory from a PAH-contaminated soil collected from Tsing Yi, Hong Kong (Wong et al., 2002; Zhao and Wong, 2009). Both strains can use phenanthrene as a sole carbon source. Under thermophilic condition, the maximum specific growth rates of BUM and BU42 in nutrient broth (Difco) were 0.21 and 0.19 h−1, which are significantly higher than that under 30°C of 0.12 and 0.13 h−1, respectively.
Biodegradation of individual PAHs by BUM and BU42
Series of experiments were conducted to assess the biodegradation of individual PAHs, that is, phenanthrene (96% purity; Sigma-Aldrich) and benzo[a]pyrene (98% purity; Sigma-Aldrich). All experiments in the following sections were preformed in Bushnell-Haas medium (Difco) at 55°C, unless indicated otherwise. Bacteria were cultured in nutrient broth (Difco) amended with phenanthrene. The cells of BUM and BU42 were centrifuged, washed, and resuspended in the Bushnell-Haas medium, to give a final density of 1 × 107 colony forming unit (CFU) mL−1. Phenanthrene or benzo[a]pyrene dissolved in dichloromethane (99.5% purity; Labscan) was carefully added to 20-mL vials, and dichloromethane was allowed to rapidly evaporate in a fume hood before 10 mL of Bushnell-Haas medium containing cells of BUM or BU42 was added to the vials. The initial concentration of phenanthrene or benzo[a]pyrene was 250 mg L−1. The vials were covered with aluminum foil, loosely capped with plastic caps, and then placed on a gyratory shaker conditioned at 150 rpm and 55°C. For every 12 h, the vials were uncapped for 30 min to avoid the development of anaerobic condition. Water loss due to evaporation during the experiment was compensated by adding sterilized distilled deionized water to the vials. Periodically, triplicate vials were removed to determine the growth of microorganisms and the amount of residual PAHs in the test vials. The growth of PAH-degrading microorganisms was evaluated by agar-spread technique. Serial dilutions of samples were made and 0.1 mL of each selected dilution was plated on the nutrient agar (BD Difco). The residual PAHs in the vials were extracted three times with n-hexane after the lysis of cells with addition of 0.1 mL of 10 N NaOH to each vial. The extracts were combined and concentrated to an appropriate volume to determine PAH concentrations using high-performance liquid chromatography (HPLC). To distinguish between abiotic losses and microbial degradation, abiotic control with 200 mg L−1 HgCl2 was prepared and incubated following the same procedures as described earlier.
Effects of cosubstrates on the biodegradation of benzo[a]pyrene
Phenanthrene and glucose were used individually as cosubstrates of benzo[a]pyrene. Different amounts of phenanthrene or glucose were added to the 20-mL vials containing 250 mg L−1 benzo[a]pyrene, and then 10 mL of Bushnell-Haas medium containing cells of BUM or BU42 at a concentration of 107 CFU mL−1 was added to the vials and the vials were incubated on a gyratory shaker conditioned at 150 rpm and 55°C. The initial concentrations of phenanthrene and glucose were 50, 250, and 500 mg L−1. Periodically, triplicate vials were removed to determine the residual PAH and microbial growth. The concentration of glucose in the cell-free media was assessed indirectly from the dissolved organic carbon (DOC) concentrations by Shimadzu ASI 5000A total organic carbon analyzer.
Effect of inoculation of BUM on the biodegradation of PAHs during thermophilic composting
Sandy-loamy soil (56.4% sand, 22.2% slit, and 21.4% clay) collected from abandoned shipyards at Northern Tsing Yi, Hong Kong, was air-dried at room temperature. The soil was sieved to <2 mm and then spiked with phenanthrene and benzo[a]pyrene dissolved in dichloromethane to a final concentration of 250 mg kg−1 for each of the two PAHs. The dichloromethane in soil was allowed to evaporate in a fume hood and the spiked soil was stored at room temperature for 6 months for aging. The bench-scale composting experiment was carried out in 1.0-L composting tanks and each treatment was performed in triplicate. To each tank, 300 g of air-dried soil spiked with PAHs was added and then mixed with pig manure at a ratio of 3:1 (soil:pig manure on dry weight basis). The basic properties of soil and pig manure used in this study are given in Table 1. The moisture content of composting material was adjusted to about 70% of its water holding capacity. BUM was inoculated into the composting material at various initial concentrations, that is, 0, 107, and 108 CFU g−1. The tanks were then aerated by pumping air at a flow rate of 200 mL min−1. A condenser was connected to the outlet of each flask to reduce moisture loss from the composting system. All tanks were incubated in an incubator maintained at 55°C for 42 days. To distinguish between abiotic losses and microbial degradation of PAHs, abiotic control with 200 mg kg−1 HgCl2 was prepared and incubated following the same procedures as described earlier. Periodically, samples were collected from each tank and PAHs in composting material were extracted with Soxhlet extraction (USEPA method 3540) and quantified with an HPLC equipped with a fluorescence detector.
The pH and EC were measured in a 1:5 (w/v) solid:water suspension.
Assay methods
All PAHs analysis was performed with 10 μL of samples separated using HPLC and fluorescence detector. Ultrasphere C18 column (5 μm × 4.6 mm × 25 cm; Beckman Coulter) with 100% acetonitrile as mobile phase and a flow rate of 1.5 mL min−1 (Wong et al., 2002) was employed.
Statistical analyses
Statistical analyses were performed on the triplicate data for means and standard error. Treatments were subjected to one-way analysis of variance and Duncan's multiple range test using SPSS ver.11.5 software.
Results and Discussion
Biodegradation of individual PAHs
Figure 1a shows the growth of BUM and BU42 in Bushnell-Haas medium amended with phenanthrene or benzo[a]pyrene. The growth of BUM on phenanthrene showed a growth curve with exponential phase followed by a stable phase from day 3 to day 30, with a peak population of 3.9 × 107 CFU mL−1 at day 18, whereas the stable phase of the growth curve of M. vanbaalenii BU42 was not apparent because the cell concentration increased in the first 6 days and then sharply decreased after the peak population was reached at day 6. When benzo[a]pyrene was used as a sole carbon source, no growth was observed and the population of the two bacteria decreased sharply, indicating that neither BUM nor BU42 could utilize benzo[a]pyrene as the sole carbon source for their growth.
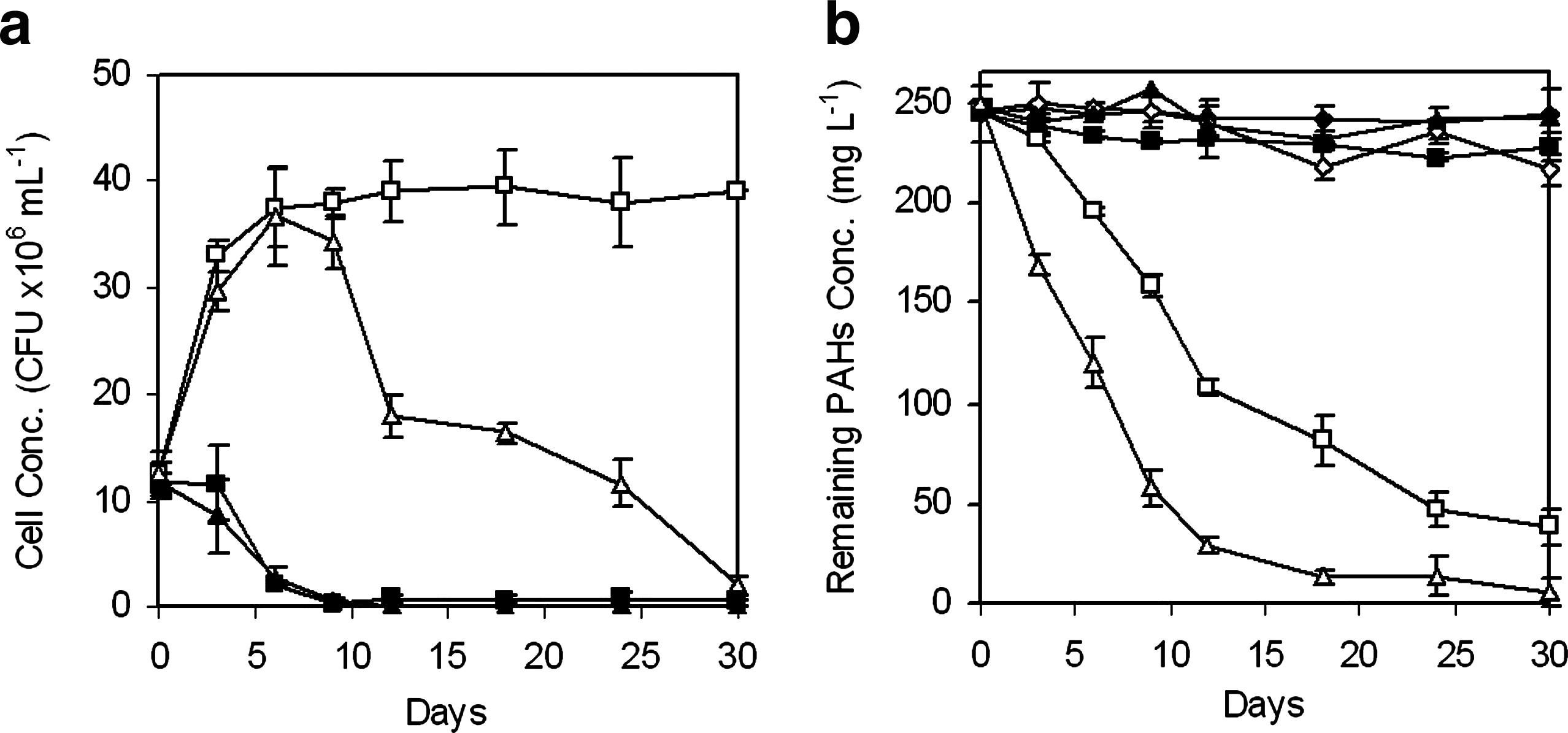
Biodegradation of individual PAHs.
Figure 1b shows the biodegradation pattern of phenanthrene and benzo[a]pyrene. There was about 11% and 3% loss of phenanthrene and benzo[a]pyrene, respectively, through volatilization and other abiotic processes during the experimental period in the abiotic controls. Hence, abiotic processes should play a minor role in the degradation of PAHs. Within the experimental period of 30 days, over 82% of phenanthrene was removed by BUM, whereas over 97% of phenanthrene was removed by BU42. However, no biodegradation of benzo[a]pyrene by both strains was noted when benzo[a]pyrene was used as the sole carbon source.
Effects of cosubstrates on the biodegradation of benzo[a]pyrene
As shown in Fig. 2, the growth of BUM and BU42 increased with increasing initial concentrations of phenanthrene. Compared with the treatments using 250 mg L−1 phenanthrene as the sole carbon source (Fig. 1), addition of benzo[a]pyrene did not significantly enhance the growth of both BUM and BU42. Therefore, we assume that the presence and consumption of benzo[a]pyrene did not contribute to the proliferation of these strains as PAHs with high molecular weight such as benzo[a]pyrene may not serve directly as carbon and energy sources for microbial populations during degradation (Juhasz and Naidu, 2000). Similar result has been reported by Rentz et al. (2005), who studied the cometabolism of benzo[a]pyrene by Sphingomonas yanoikuyae JAR02 in the presence of plant root extracts and exudates. Figure 3 shows the biodegradation of benzo[a]pyrene in the presence of phenanthrene as the cosubstrate for B-UM and BU42. There was no apparent biodegradation of benzo[a]pyrene by BU42 strain in the presence of phenanthrene. However, the presence of phenanthrene significantly (p < 0.05) increased the biodegradation of benzo[a]pyrene by BUM, and the removal rate of benzo[a]pyrene increased with the increasing initial concentrations of phenanthrene. It has to be highlighted that the degradation of benzo[a]pyrene occurred simultaneously with the biodegradation of phenanthrene. In the presence of 50 mg L−1 phenanthrene, the removal ratio of benzo[a]pyrene was 17.3% in the first 12 days. After the depletion of phenanthrene in the medium at day 12, the biodegradation of benzo[a]pyrene ceased.
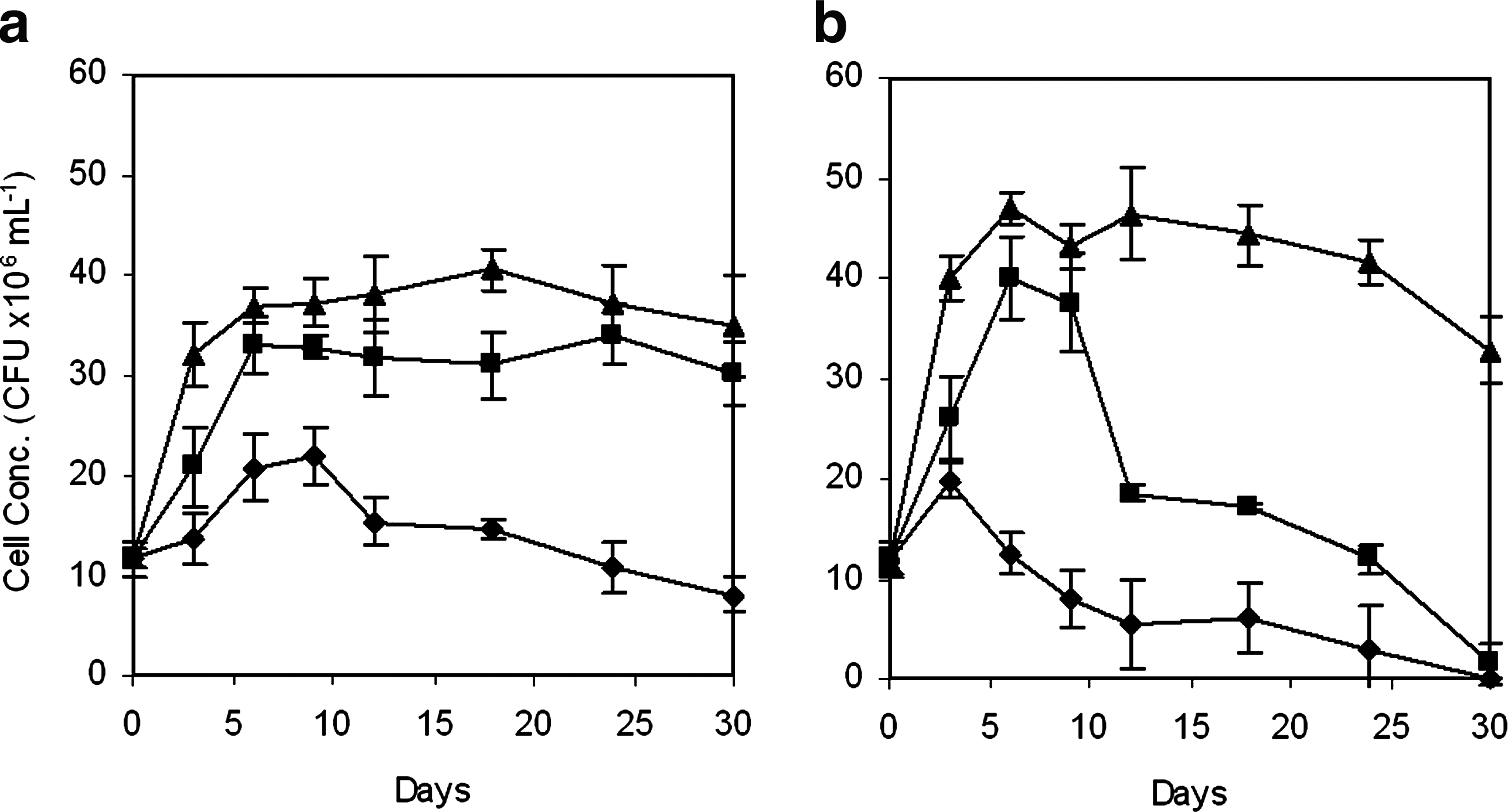
Growth of microorganisms on phenanthrene as a cosubstrate of 250 mg L−1 benzo[a]pyrene.

Biodegradation of 250 mg L−1 benzo[a]pyrene in the presence of phenanthrene at
The growth of the two bacterial strains in the presence of glucose as cosubstrate is shown in Fig. 4a. The concentrations of the bacteria were dramatically increased in the first 3 days of incubation, whereas almost all glucose had been depleted by microorganisms in the same period as indicated by the extremely low dissolved organic carbon concentration (<5 mg L−1) in the cell-free supernatants (data not shown). After the log growth observed in the initial 3 days, the cell concentrations decreased dramatically. However, the increase in the growth of microorganisms by glucose did not benefit the biodegradation of benzo[a]pyrene. As shown in Fig. 4b, no obvious biodegradation of benzo[a]pyrene was found in the inoculated treatments compared with the abiotic control.
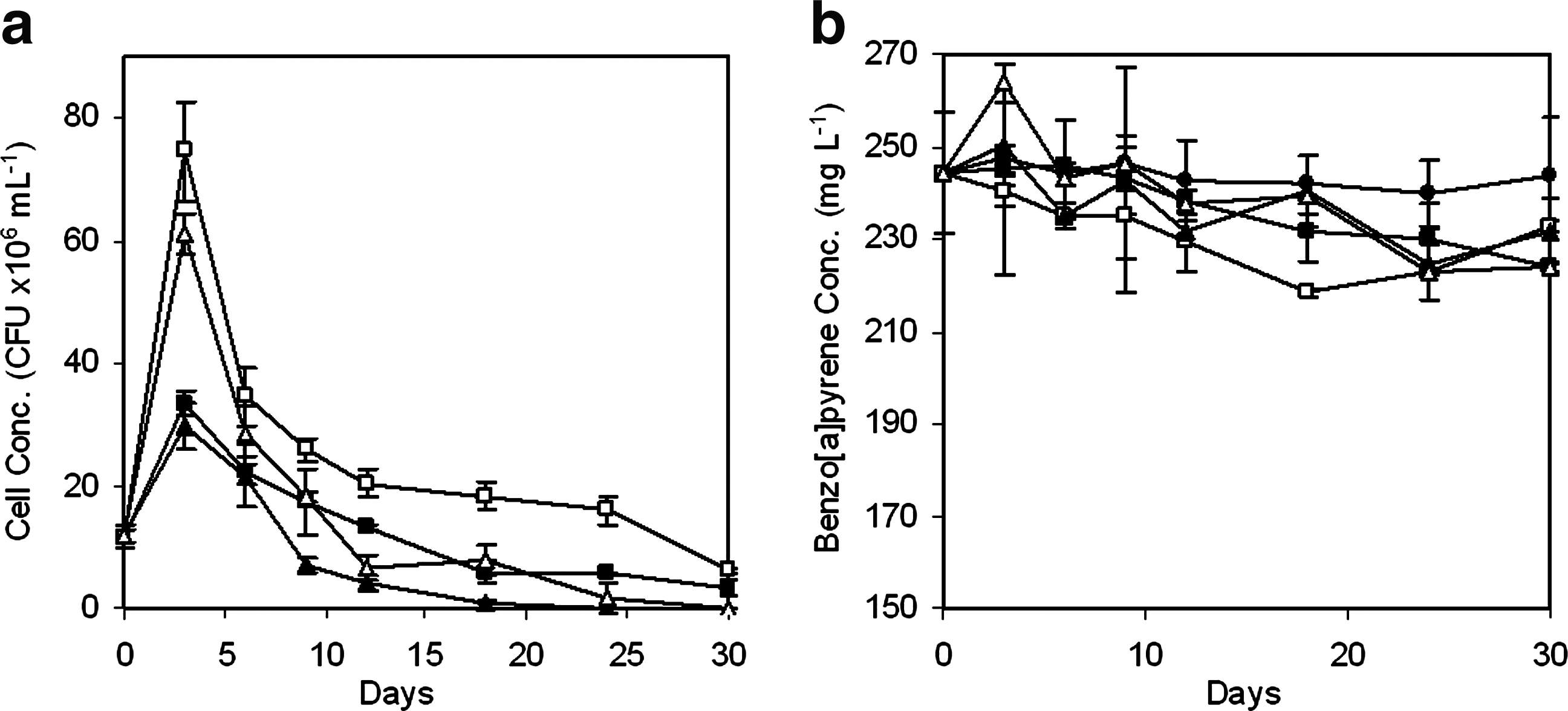
Biodegradation of benzo[a]pyrene in the presence of glucose as cosubstrate.
In this study, phenanthrene was easily degraded at a high degradation rate when it was used as a sole carbon source, whereas benzo[a]pyrene did not undergo significant biodegradation. The low uptake rates of PAHs with high molecular weight have been cited as one major reason responsible for their slow metabolism (Goswami et al., 1983). In the present study, however, the lack of biodegradation of benzo[a]pyrene as the sole carbon source may not result from the poor uptake, as our previous study showed that the highly lipophilic cell surface of BUM could facilitate the uptake of hydrophobic compounds (Wong et al., 2004). Therefore, it may be due to that benzo[a]pyrene may not be a suitable inducer of the catabolic pathway of PAHs or may have poor bioavailability to bring about adequate biomass for rapid biodegradation. When added as a cosubstrate, phenanthrene successfully initialized the biodegradation of benzo[a]pyrene by BUM.
The finding that BUM degraded and grew on benzo[a]pyrene in the presence of more bioavailable phenanthrene is of special interest, as PAHs of both high and low molecular weights coexist in soil in most of the contaminated sites. The experimental results revealed that the positive effect of phenanthrene on benzo[a]pyrene biodegradation may be possibly due to the induction of the catabolic pathway, as the increase in the growth of microorganisms by glucose did not benefit the biodegradation of benzo[a]pyrene (Cerniglia, 1993). The cometabolic degradation of benzo[a]pyrene requires an inducer to maintain or express PAH-degrading enzymes (Rentz et al., 2008). PAHs with low molecular weight, for example, fluoranthene, phenanthrene, and pyrene, are proved to be suitable inducers for cometabolism of benzo[a]pyrene by various microorganisms (Ye et al., 1996; Boonchan et al., 2000). The positive effect of phenanthrene on the biodegradation of benzo[a]pyrene was also previously reported by McLellan et al. (2002), who suggested that the four- to sixfold increases in the biodegradation of benzo[a]pyrene in the presence of other PAH compounds was due to the involvement of a downstream pathway that acts on the smaller structures and provides further metabolism of the products of partial degradation of benzo[a]pyrene.
Effect of inoculation of BUM on the biodegradation of PAHs during thermophilic composting
The loss of phenanthrene through volatilization and other abiotic processes was about 18%, whereas no significant loss was observed for benzo[a]pyrene (Fig. 5). Within a composting period of 42 days, the removal of benzo[a]pyrene without inoculation of BUM was 30.3%, whereas almost all of phenanthrene was removed. As expected, inoculation of BUM significantly enhanced the degradation of benzo[a]pyrene to 52.2%, but no significant difference was observed between the two treatments with different inoculation amounts. For all the treatments, biodegradation of benzo[a]pyrene mainly occurred in the first 28 days of composting, but no significant biodegradation was observed after the depletion of phenanthrene on day 28.
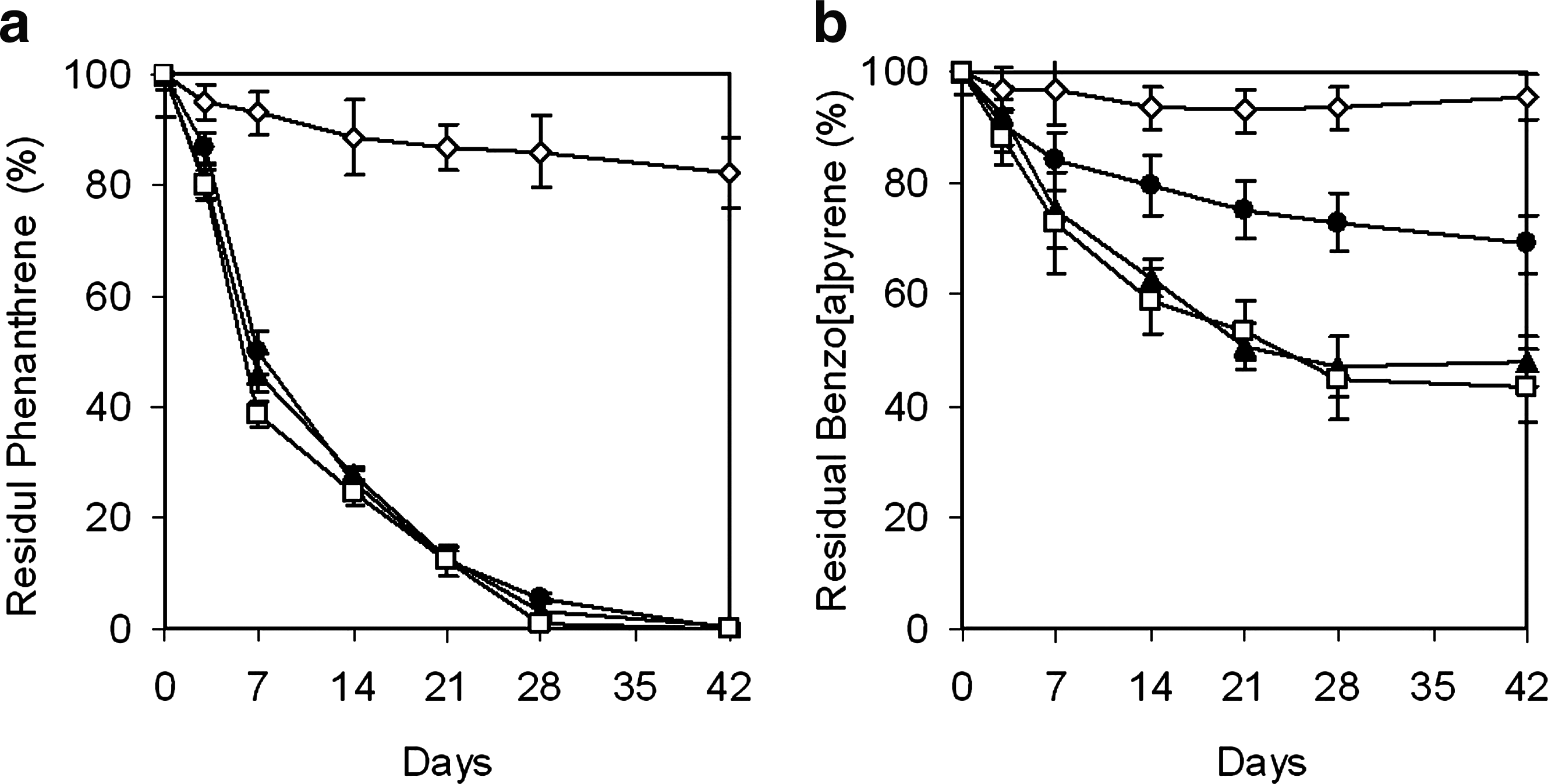
Removal of
Although numerous microorganisms have been found to degrade low-molecular-weight PAHs, only a few have been reported with the ability to degrade high-molecular-weight PAHs. Mycobacterium sp. and Sphingomonas sp. are the most popular high-molecular-weight PAH-degrading bacteria as reported (Schneider et al., 1996; Ye et al., 1996; Moody et al., 2001; McLellan et al., 2002; Su et al., 2008). However, compared with those PAHs with low molecular weight, the biodegradation of benzo[a]pyrene was extremely slow. The resting cells of Sphingomonas paucimobilis EPA505 was shown to degrade 33% of 10 mg L−1 benzo[a]pyrene in 16 h (Ye et al., 1996). A bacterial consortium including members of the genera Mycobacterium and Sphingobacterium mineralized benzo[a]pyrene in the presence of diesel fuel and the degradation rate was up to 1.08 mg L−1 day−1 (Kanaly et al., 2000). Recently, B. subtilis BMT4i was reported to remove about 74% of 50 mg L−1 benzo[a]pyrene within 21 days (Lily et al., 2009).
In the presence of phenanthrene, the rapid biodegradation of benzo[a]pyrene by BUM was observed in both aqueous and composting systems under thermophilic condition. The zero-order degradation rate was calculated as follows:
where Rde is the zero-order degradation rate, C0 is the initial concentration of PAHs, Ct is the concentration of PAHs at time t, and t is the time plot for the biodegradation of PAHs. The peak degradation rates observed in the first 3 days of biodegradation were 12.3 mg L−1 day−1 and 9.7 mg kg−1 day−1, respectively, in aqueous and soil composting systems. This is the first report of biodegradation of benzo[a]pyrene by an isolated thermophilic microorganism. Compared with mesophiles, the isolated thermophilic strain BUM removed benzo[a]pyrene more rapidly possibly because of (1) the elevated temperature that is expected to increase the solubility and mass transfer rates of PAHs (Viamajala et al., 2007) and (2) the higher substrate utilization rates of thermophilic bacteria, that is, 3–10 times greater than that of mesophilic bacteria (Lapara and Alleman, 1999). Under thermophilic condition (55°C), a high solubility and desorption of phenanthrene was achieved in our previous study (Cheng et al., 2004). Similarly, the biodegradation of phenanthrene by thermophilic Geobacilli strains was more rapid than mesophiles (Viamajala et al., 2007). However, the octanol–water partition coefficients of PAHs, termed as Kow, decreased at elevated temperatures (Viamajala et al., 2007), which may decrease the sorption of benzo[a]pyrene onto the hydrophobic cell surface of BUM. However, the sorption concentration of PAHs is dependent upon both the aqueous solubility and the Kow. The enhanced solubility under thermophilic condition could likely compensate the negative effect of decreased Kow as the increase of temperature from 20°C to 60°C increased the solubility of phenanthrene, fluorine, and fluoranthene by 6.03-, 4.87-, and 8.56-fold, while decrease the values of Kow by only 59.4%, 44.5%, and 62.3%, respectively (Viamajala et al., 2007).
The Pearson's correlation coefficients between the zero-order degradation rates of phenanthrene and benzo[a]pyrene were determined. A statistically significant correlation was found in both the aqueous biodegradation (r = 0.79, p < 0.01) and soil composting systems (r = 0.87, p < 0.01), strongly indicating that the degradation of benzo[a]pyrene was a cometabolic process with phenanthrene (Juhasz and Naidu, 2000).
Conclusions
The feasibility of biodegradation of benzo[a]pyrene by isolated thermophilic strains was investigated. In the presence of phenanthrene as cosubstrate, a thermophilic strain, that is, BUM, rapidly degraded benzo[a]pyrene and its zero-order degradation rate was up to 12.3 mg L−1 day−1 in aqueous biodegradation experiment and 9.7 mg kg−1 day−1 in soil composting system. Statistically significant correlations were found between the degradation rates of benzo[a]pyrene and that of phenanthrene, indicating that the presence of inducing compounds such as phenanthrene may be an important factor in increasing the capacity of microorganisms in degrading benzo[a]pyrene under thermophilic condition.
Footnotes
Acknowledgment
The authors thank the Research Grant Council of the Hong Kong Special Administrative Region, People's Republic of China (grant HKBU261307), for the financial support.
Author Disclosure Statement
The authors declare that no competing financial conflicts exist.