
Abstract
Abstract
Phytoremediation uses plants for environmental cleanup. However, various shortcomings have prevented routine adoption and use of this technology by engineers. For the purposes of heavy metal extraction, one such shortcoming has involved insufficient knowledge of plant metal trafficking biochemistry. This research focuses on the xylem sap concentrations of thiols such as phytochelatins (PCs), which are small peptides with the sequence (γ-Glu-Cys) n -Gly and are a major class of heavy metal-binding peptides in plants. Despite their importance in heavy metal binding and sequestration, their possible role in root-to-shoot transport is not well understood and is the source of some conflicting reports in the research literature. However, understanding root-to-shoot transport is critical for rational design and successful use of phytoremediation. In this study, we used a flowing culture system to grow Brassica napus and subsequently analyze its xylem sap for the presence of PCs and other thiols in response to low, but environmentally relevant, Cd concentrations ranging from 0.5 to 5.0 μM. Results indicated that PCs and cysteine were detectable in the B. napus xylem sap within 24 h of initial Cd exposure and responded to Cd in a time- and dose-dependent manner. This work is the first to focus on low Cd concentrations and demonstrate temporal and concentration-dependent PC responses in the xylem sap, which may ultimately help in developing better plants for heavy metal extraction.
Introduction
Phytoremediation involves using plants to clean up a polluted environment. When using plants to remove pollutants, such as heavy metals, from soil the term phytoextraction is used (Salt et al., 1998). The ideal plant for heavy metal phytoextraction should have the following properties: accumulation of the target metal to high concentrations, high translocation to above-ground plant biomass, fast growth, high biomass production, and easy harvestability (Karenlampi et al., 2000). However, incomplete knowledge of how to use plants effectively to extract heavy metals from environmental systems has hampered engineering efforts and implementation of this technology (Clemens et al., 2002).
One key area where our current knowledge is incomplete involves the mechanisms used by plants to traffic and transport heavy metals from roots to above-ground plant tissue. Some aspects of plant heavy metal trafficking are understood. Numerous classes of membrane transporters have been identified, which have important roles in metal uptake and transport to various organelles, and important components of in-planta metal trafficking networks have been now understood (Hall and Williams, 2003; Clemens, 2006b; Palmer and Guerinot, 2009). Further, the widespread use of tools from both genomics and molecular biology provides useful insights into the structural, mechanistic, and regulatory motifs present in numerous transport proteins, such as P-type ATPases (Kuhlbrandt, 2004). Despite these advances, however, important questions involving heavy metal transport from roots to shoot have remained unanswered. Basically, these questions involve the following: (1) how do heavy metals enter and exit the xylem, and (2) what metal-binding ligands are used in long-distance xylem transport.
Xylem transport pathways are at least partially understood. Several research reports have indicated that the plasma membrane transporters HMA2 and HMA4, members of the P1B class of P-type ATPases, are important for xylem loading of Zn and Cd (Hussain et al., 2004; Papoyan and Kochian, 2004; Verret et al., 2004; Mills et al., 2005; Talke et al., 2006; Courbot et al., 2007; Hanikenne et al., 2008; Wong and Cobbett, 2009). However, additional work will be needed before a complete picture of xylem transport proteins will emerge (Palmer and Guerinot, 2009).
Other research has focused on the roles of certain metal-binding ligands used by plants. As nearly all metal ions are expected to be ligand-bound rather than exist freely in the cytoplasm (Clemens, 2006b), understanding the role of these compounds is important for efforts directed toward engineering plants to accumulate high shoot metal levels. Phytochelatins (PCs) are a major type of heavy metal-binding peptide, which were first isolated in plants from cell suspension cultures and had the general structure (γ-glu-cys) n -gly (n = 2–11) (Grill et al., 1985). These peptides are nontranslationally synthesized by PC synthase in response to a variety of metals (Clemens et al., 1999; Ha et al., 1999; Vatamaniuk et al., 1999). They are also widely distributed in nature because of their universal presence in higher plants and have been found in representatives across all eukaryotic kingdoms (Clemens, 2006a). Generally, once synthesized and bound to heavy metal ions, PCs have been suggested to be primarily involved in shuttling the metal to the vacuole for storage and sequestration (Zenk, 1996; Cobbett, 2000; Clemens, 2001).
However, PCs may have an additional role in long-distance transport of heavy metals via binding during xylem transport, although some controversy exists in the research literature on this topic. In one study, X-ray absorption spectroscopy was used to examine binding ligands for Cd in Brassica juncea (Salt et al., 1995). In roots and shoots, Cd was coordinated to four S ligands, whereas the xylem sap data indicated Cd coordination with six oxygen or nitrogen ligands. In a review (Salt et al., 2002), it was suggested that translocated Cd resulted from two possibilities: either it never underwent sulfur coordination in roots and was therefore never stored, or it was moved from storage to transport forms through an active process. Possible mechanisms behind this potential ligand switching were not understood.
Later studies demonstrated that PCs were capable of long-distance transport. Using the cad1-3 mutant of Arabidopsis thaliana (a PC synthase deficient line), a PC synthase gene from wheat (TaPCS1) was transformed into this strain under the control of the alcohol dehydrogenase promoter, which limited gene expression to root tissue (Gong et al., 2003). Despite this, PCs were detected in the stem and leaf tissue of plants expressing Adh::TaPCS1, which provided evidence for long-distance root-to-shoot transport of PCs (Gong et al., 2003). Although overall PC levels were similar between wild-type controls and plants that expressed TaPCS1, transgenic lines contained more Cd in their stem and leaf tissue. The mechanism behind this phenomenon, however, was unclear. Later work found evidence for long-distance transport of PCs in the opposite direction (shoot-to-root) in Arabidopsis as well (Chen et al., 2006). Here, TaPCS1 was expressed in cad1-3 plants and was under control of the shoot-specific CAB1 promoter. Although the transgenic plants were unable to rescue the Cd sensitivity of cad1-3 roots, PC2 was clearly detected in root tissue. Further, grafting of a wild-type shoot onto a cad1-3 atpcs2-1 mutant resulted in detection of PC2, PC3, and PC4 in roots, which demonstrated that under their native promoter, PCs were also capable of shoot-to-root transport (Chen et al., 2006). Taken together, these papers provided strong, although indirect, evidence for long-distance PC transport in plants—or at least Arabidopsis—and therefore possible involvement of PCs in long-distance metal transport.
Another study directly analyzed the xylem sap to understand the potential role(s) of PCs. Using size-exclusion high-performance liquid chromatography (HPLC) coupled with a UV detector, fraction collector, and offline graphite furnace atomic absorption spectroscopy, PCs were detected in the B. juncea xylem sap under Cd stressed conditions, which consisted of a 50 μM Cd exposure for 10 h (Wei et al., 2007). A 10 μM Cd exposure for 10 h did not apparently induce significant PC transport in the xylem sap, although detection limits of their system were not reported. Graphite furnace atomic absorption spectroscopy measurements on the collected fractions suggested that 29% of the xylem sap Cd was present as PC-Cd, whereas the majority (68%) was bound to organic acids or inorganic ions (Wei et al., 2007). Conversely, other researchers used Brassica napus and found little evidence for a role involving PCs in the xylem sap transport of Cd, although higher levels of PCs were found in phloem samples (Mendoza-Cózatl et al., 2008). However, in metal speciation studies, difficulties frequently arise with finding a sampling method that does not disrupt the native chemical equilibrium of the target of interest (Callahan et al., 2006) and it cannot be ruled out that sampling and preparation methods may have altered the chemical equilibrium of the xylem sap in previous studies.
Therefore, the role of PCs in long-distance transport has not been satisfactorily resolved. Plant molecular biologists have shown the capability of PCs to undergo long-distance transport, but direct analysis of the xylem sap has led to conflicting results. Further, the concentration of Cd that was used to obtain a strong xylem sap response in the work of Wei et al. (2007) was likely higher than levels found at some Cd-contaminated sites such as brownfields or Cd-enriched agricultural soils (Klang-Westin and Eriksson, 2003; Murray et al., 2004). Thus, there is an open question regarding the role of PCs in long-distance xylem transport in response to Cd concentrations that are closer to what may be encountered at a diffusely contaminated site. Here, we present work that was focused on measurement of PC2 and other thiol-containing compounds in response to Cd exposure in the xylem sap of B. napus, a plant with phytoremediation potential.
Experimental Protocols
B. napus cv. Quantum seeds were kept refrigerated at 4°C until needed and were germinated in an aerated glass beaker placed in a controlled temperature water bath as previously described (Saathoff et al., 2009). B. napus plants were grown in a growth chamber in Kenneth Post Laboratory at Cornell University. Lighting was based on a 16-h light/8-h dark cycle, and chamber temperature was set at 23°C/20°C during the light and dark cycles, respectively. Plants were grown in a liquid culture-rearing system that was previously described (Saathoff et al., 2009), which involved placement and rearing of up to 32 plants in a 48-L nutrient tank (Harvester 2 × 4; CSL Plastics, Inc.). Nutrient solution was circulated through polypropylene filters using an impeller-driven pump (Model 1046; Eheim GmbH) at a rate of 606 L/h to assist in keeping the nutrient solution clean. However, use of the rearing system in an environmentally controlled growth chamber eliminated the need for a water-filled heat sink that was used in the original system. Plant growth was staggered such that eight plants were ready for use each week and plants were randomly placed within the rearing unit to eliminate any potential effects of location within the rearing unit on experimental results. Nutrient solution was a modified Johnson's solution (Johnson et al., 1957), and approximately 64% of the system solution was replaced weekly. Plants were grown in this rearing system starting with day 5 of the growing schedule and remained in this unit until days 28–30, when they were moved into the trough system, depending on the experiment that was being conducted.
Experimental trough system
The trough system used in these experiments was an adaptation and simplification of an Experimental Cadmium Uptake Detection System, which was previously described elsewhere (Saathoff et al., 2009). Briefly, the simplified system used two 1.82-m troughs (American Hydroponics, Inc.) with a nutrient reservoir for each trough. The solenoid valve system as well as the cadmium ion-selective electrode data acquisition system of the original design was not used in this configuration. A new stand was constructed to accommodate these troughs, allowing them to be placed side-by-side in the growth chamber. Fluid for each trough was pumped by a Masterflex LS digital drive peristaltic pump that was equipped with two pump heads (Masterflex Easyload 3; Cole-Parmer, Inc.). Flow rate was set at 200 mL/min and total system volume for each trough in these experiments was 10 L, which resulted in a mean volumetric retention time of 50 min. Typically, 4–5 plants from the rearing system were placed into the trough system for an experiment. Total root biomass averaged 1.0–1.5 g (dry weight) for each experiment. The system was generally operated in recirculation mode, where trough effluent reentered the storage reservoir via gravity flow and was then pumped from this reservoir back into the trough.
Xylem sap collection experiments
On days 28–30 (depending upon the desired Cd exposure time), plants were removed from the rearing unit and placed into the trough system. To minimize the effects of transport and handling shock, B. napus plants were always allowed to equilibrate in the trough system for 24 h with a nutrient solution that was lacking Fe-HEDTA and Cu prior to Cd treatment. After equilibration, system solution was changed to a new nutrient solution that still lacked Fe-HEDTA and Cu, but contained Cd. Plants were exposed to this Cd-containing nutrient solution for one of three time periods 24, 48, or 72 h, depending on the experiment. No additional Cd was added to the system in any of the experiments after the initial dosage. Once the desired time period had passed, plant stems were cut and xylem sap collection was initiated; collection experiments were designed so that they always occurred on day 32 of a growth cycle. Cadmium concentrations of 0.5, 1, and 5 μM were used. These concentrations are similar to those that may be encountered in Cd-contaminated agricultural soils or brownfield sites (Klang-Westin and Eriksson, 2003) and were used in an attempt to mimic the typically low bioavailability of Cd in soils (Zhuang et al., 2005).
Xylem sap was collected by detopping each plant just below the cotyledon node using a fresh razor blade (VWR International). New razor blades were used for each plant to ensure sharpness and to minimize sample contamination. Immediately after detopping, the stem was washed for 2 min under a constant stream of 18 MΩ cm deionized water. Then, depending upon stem size, a 1½-inch-long tubing ( or ¼ inch internal diameter; VWR International) was fitted to the tops of the stems. Silicone grease (Radiator Specialty Products, Inc.) was used to seal the base of the tubing on the stem to avoid potential leakage. Xylem sap was then allowed to exude under normal root pressure and was collected at hourly intervals for 3 h using a micropipette. Sap was placed into 1.5-mL plastic tubes, which were kept on ice during the collection period. Immediately after collection, a small amount (5–30 μL, depending upon sap volume) of 800 mM methanesulfonic acid (MSA; Sigma-Aldrich, Inc.) was added to each tube so that the final MSA concentration was approximately 10 mM to denature proteins and preserve thiols. Samples were then stored in a −20°C freezer until HPLC analysis, which was conducted within 72 h of collection.
In all experiments, Cd depletion in troughs was monitored to verify uptake by pipetting 10 mL samples of the trough effluent into 15-mL disposable plastic tubes (VWR International). Samples were then analyzed for Cd content using an inductively coupled plasma atomic emission spectrometer (ICP-AES) unit (Thermo Jarrel Ash ICAP-60) that had previously undergone extensive modifications (Rutzke, 2002). This instrument could accurately detect Cd concentrations down to about 0.015 ppm.
Xylem sap thiol measurements
Xylem sap thiols were analyzed by following a previously published method (Wei et al., 2003) with some slight alterations. Briefly, this method involved labeling thiols with monobromobimane (mBrB) followed by separation and quantification on an HPLC (Kosower et al., 1979; Newton et al., 1981; Ahner et al., 1995). Borate-DTPA buffer was added to samples, followed by the addition of DTT to reduce disulfide bonds. Samples were then reacted with mBrB according to the method of Newton et al. (1981). MSA was added at the end of the reaction to stabilize the fluorescently tagged thiols. Chromatography was carried out on a Shimadzu HPLC equipped with a fluorescence detector (RF-10AXL) set to an excitation wavelength of 390 nm and an emission wavelength of 478 nm. The column (Supelco Discovery RP Amide C16) was kept at 25°C in an oven (Prominence CTO-20AC) during all HPLC runs. The mobile phases consisted of HPLC-grade acetonitrile (Mallinckrodt Chemicals ChromAR) and 25 mM KH2PO4/H3PO4 buffer at pH 3.55. A lower pH buffer was used by Wei et al. (2003), but buffer at that pH did not sufficiently resolve all thiol groups of interest on our system. This method was capable of simultaneously resolving four distinct thiols: cysteine (J.T. Baker, Inc.), glutathione (GSH; Sigma-Aldrich, Inc.), γ-glutamyl cysteine (γ-EC; Sigma Aldrich, Inc.), and PC (n = 2; Cell Essentials, Inc.).
Data analysis
Thiol concentration results from the HPLC were averaged across replicates. Analysis of variance (ANOVA) was used to determine differences in mean thiol concentrations among treatment groups. The factor, Cd concentration, was considered a fixed effect in this model, and treatment groups consisted of plants that were exposed to Cd for the same period of time. MINITAB 15 (Minitab, Inc.) was used for all ANOVA computations and results were considered statistically significant at the α = 0.05 level. Further analysis was done among treatment means by using the Tukey–Kramer multiple comparison procedure with the family Type I error rate controlled at α = 0.05.
Results
B. napus was capable of rapidly depleting Cd from the flowing culture system to levels below the detection limit of the ICP within 24 h (data not shown). The thiol derivation protocol adapted from the work by Wei et al. (2003) provided a very sensitive method for detection of thiols labeled with mBrB. This method allowed for detection of cysteine, γ-EC, GSH, and PC (n = 2) in amounts down to 5 pmol in a 100 μL volume. Higher-order PCs, however, were not well resolved using this technique. A chromatogram of a typical xylem sap sample is shown in Fig. 1.
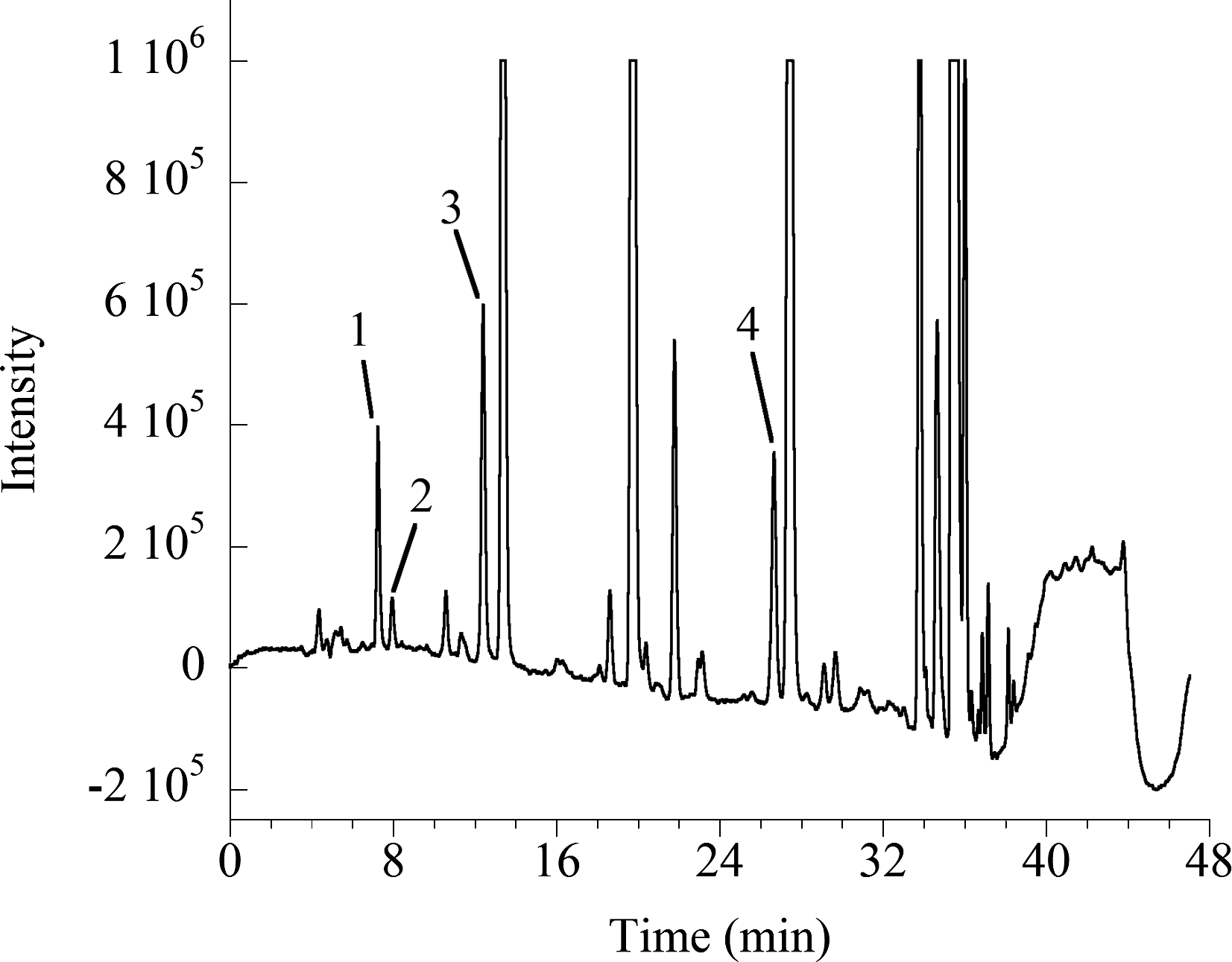
HPLC chromatogram of a xylem sap sample containing (1) cysteine, (2) γ-glutamyl cysteine, (3) glutathione-like, and (4) PC2. PC, phytochelatin.
The xylem sap cysteine results from the Cd exposure experiments across three time periods and three Cd concentrations are shown in Fig. 2A. Across the tested Cd concentrations, elevated cysteine levels were observed during the 48- and 72-h time periods. ANOVA revealed no meaningful differences among Cd treatments in the 24-h treatment group (p = 0.64), but did find differences among the 48- and 72-h exposure groups (p < 0.001). Within the 48-h group, significant differences were found between control versus 1 μM Cd (p = 0.0468), control versus 5 μM Cd (p = 0.001), 0.5 μM versus 5 μM Cd (p = 0.003), and 1 μM versus 5 μM Cd (p = 0.0172). For the 72-h Cd, cysteine data were log transformed to meet homoskedastic model assumptions, and Tukey's multiple comparison procedure showed that all 72-h Cd treatments were significantly different from the control with p-values of 0.0357, 0.002, and 0.001 for the 0.5, 1, and 5 μM Cd treatments, respectively. Additionally, significant differences were found between 0.5 μM versus 1 μM Cd (p = 0.0126) and 0.5 μM versus 5 μM Cd (p = 0.0041).

The results reported in Fig. 2A were replotted as a scatterplot (Fig. 2B) to better illustrate the temporal behavior of the xylem sap cysteine concentration in response to Cd exposure. From this plot, two observations were noted: (1) the presence of detectable cysteine in the xylem sap of unexposed control plants suggested that a basal level of cysteine was normally present, and (2) the rate of cysteine concentration change within a Cd exposure group appeared to increase over time. This observation suggested that the rate of increase may be proportional to the thiol concentration, which was integrated to the following expression:
To learn if Eq. (1) was an accurate representation of the temporal trends, it was fit to the cysteine data using KaleidaGraph 3.6 (Synergy Software). The results from the fit are presented in Table 1. These results show that the exponential model fits the cysteine concentration data reasonably well in the 1 and 5 μM Cd experiments. At the lowest Cd exposure level, 0.5 μM, this model was less adequate at explaining trends in the data (R2 = 0.77). A linear model did not result in a better fit (data not shown).
Model Fit to the Xylem Sap Cysteine Data
The results showing the xylem sap PC (n = 2; PC2) concentration are shown in Fig. 3A. In this case, PC was detected at all Cd exposure levels and across all time periods. No PC2 was detected in unexposed control plants, and thus data from control plants was not included in the PC2 ANOVA analysis. To satisfy ANOVA model assumptions for statistical analysis, data from the 24- and 72-h time periods were log-transformed. Statistically significant differences were found among the treatment means at all three time points (p = 0.005, p < 0.001, and p < 0.001 for the 24-, 48-, and 72-h treatment groups, respectively). Within the 24-h Cd group, Tukey's multiple comparison procedure found significant differences between 5 μM Cd versus 0.5 and 1 μM Cd treatments (p = 0.0064 and p = 0.0092). In the 48-h Cd group, the significant differences were 5 μM versus 0.5 μM Cd (p = 0.0006) and 5 μM versus 1 μM Cd (p = 0.0017). In the 72-h Cd group, statistically significant differences existed between the following: 0.5 μM versus 1 μM Cd (p = 0.0012), 0.5 μM versus 5 μM Cd (p < 0.0001), and 1 μM versus 5 μM Cd (p < 0.0001). When PC2 concentration data were replotted as a scatterplot (Fig. 3B), the data again suggested a rate of increase that may be proportional to thiol concentration. Eq. (1) was then fit to these data using the previously indicated method. The results from this fit are shown in Table 2, which indicates that the exponential model had a good fit to the data through 72 h. Taken together, PC2 was detected in the xylem sap of B. napus under different Cd concentrations and PC2 levels increased over time after initial Cd exposure.
Model Fit to the Xylem Sap PC2 Data
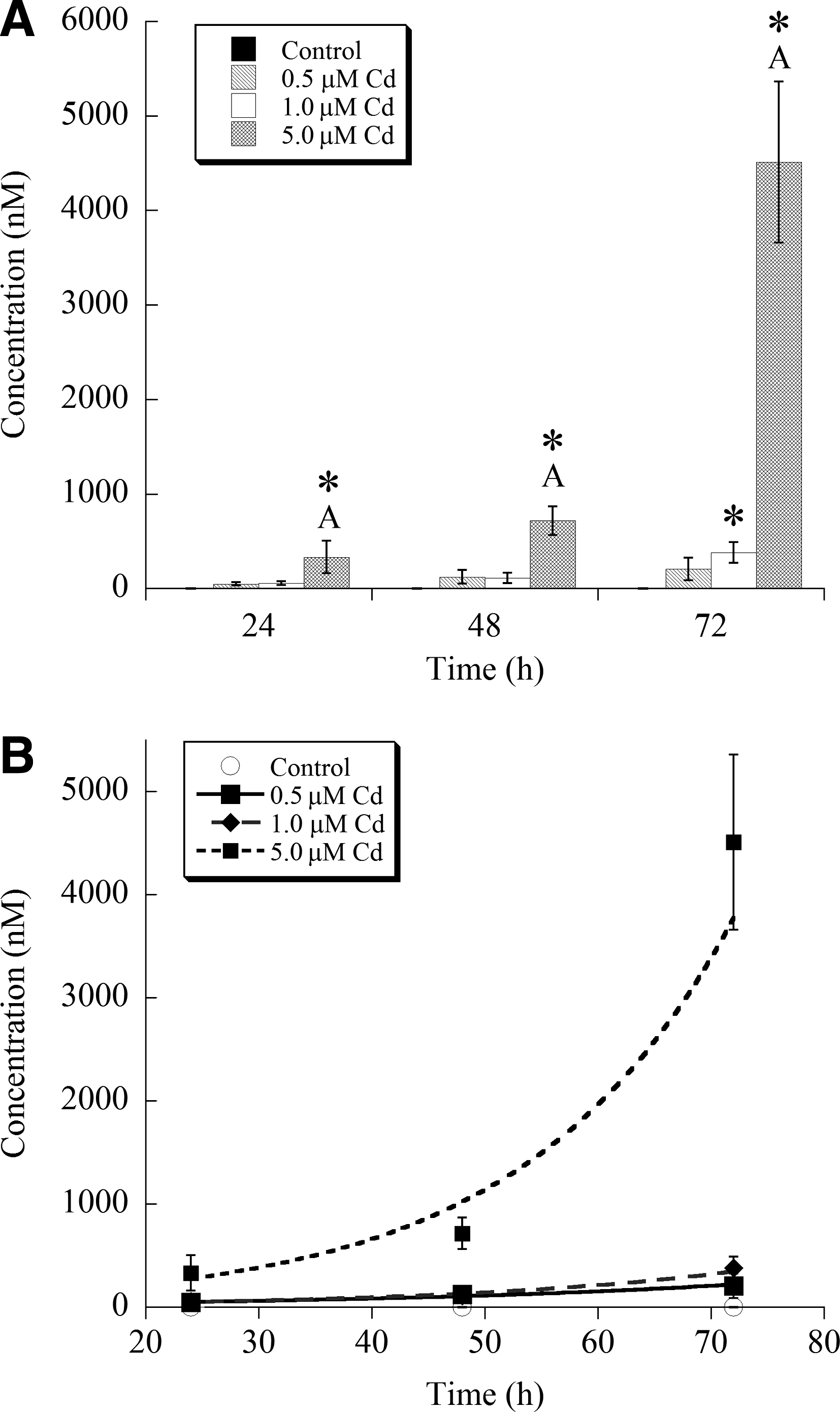
The remaining known thiols that were resolvable using this HPLC method, γ-EC and GSH, were not consistently detected in the xylem sap. In the case of γ-EC, it was only sporadically detected in the xylem sap, and when it was detected, it was at relatively low concentrations and did not appear to display a response to Cd (data not shown). Additionally, GSH, or at least GSH that was in the same form as our analytical standard, was not found in the xylem sap. However, there was clear evidence for an unknown thiol species that eluted off of the HPLC column at nearly the same time as our GSH standard. The differences in elution time between our GSH standard and this unknown thiol species were consistently between 0.2 and 0.3 min. Thus, this unknown thiol was termed “GSH-like.” Approximate concentration of this species was determined by using a GSH calibration curve, and the results are shown in Fig. 4. In the 24-h group, ANOVA found evidence for differences among the treatment means (p = 0.033), but Tukey's multiple comparison procedure found that no Cd treatments differed from controls. At 48 h, no statistically significant differences were found among the results in this treatment group (p = 0.16), although the large amount of variation within this group likely weakened the ANOVA results. At 72 h, data were log-transformed to again meet the ANOVA model assumptions, and the test indicated significant differences among treatment means (p < 0.001). Pairwise comparison procedures indicated significant differences between the control and all 72-h Cd treatments; further statistically significant differences were found between 0.5 μM versus 5 μM Cd as well as 1 μM and 5 μM Cd. In addition to this GSH-like thiol, another unknown thiol-containing species that eluted 0.7 min after cysteine was frequently present in our chromatogram. However, like γ-EC, this thiol was not always detected and approximate concentrations of this thiol always appeared to be about 500 nM or lower (estimated concentration; data not shown).

Approximate concentration of the GSH-like thiol species in the xylem sap of B. napus. An asterisk indicates a statistically significant difference from the control within a time period. Error bars represent the standard deviation of the mean. GSH, glutathione.
Discussion
In this study, we have used a flowing culture system to expose 29–31-day-old B. napus to three concentrations of Cd for three different exposure periods, and the xylem sap after these Cd exposures was examined for the presence of cysteine, GSH, γ-EC, and PC2. The primary purpose of this study was to investigate the effect of Cd on these thiol levels in the xylem sap. For thiol measurements, we used a well-established and sensitive HPLC technique that relied upon fluorescent labeling of thiols with mBrB; our system was capable of thiol measurements down to 5–10 pmol. Cd concentration was also monitored in the trough system during all Cd exposure experiments to ensure that the plants were absorbing Cd. We were unable to gather data to provide a detailed Cd profile of the xylem sap because of imprecise small-volume Cd measurements on our ICP, which resulted in the loss many of the most useful xylem sap samples. However, we were able to analyze two remaining xylem sap samples on a more precise instrument and found detectable levels of Cd in the low micromolar concentration range. These results were consistent with other research that has shown that B. napus accumulates Cd in above-ground tissue when grown on Cd-containing soil (Baryla et al., 2001; Rossi et al., 2002; Carrier et al., 2003), which strongly suggested that Cd was transported via the xylem because it was the only root-to-shoot pathway available. Also, direct measurements have shown the presence of Cd in the B. napus xylem sap (Mendoza-Cózatl et al., 2008), at least in response to higher Cd concentrations. Other work focused on the hyperaccumulator Arabidopsis halleri and demonstrated the presence of Cd in the xylem sap after plants were exposed to low micromolar Cd concentrations (Ueno et al., 2008). Thus, when taken together with our limited xylem sap Cd data, it is likely that Cd was present in all of the xylem sap samples from Cd-exposed B. napus. However, the lack of detailed xylem sap Cd data precluded drawing correlations between Cd concentration and thiol content.
The results of our thiol analysis indicated that thiols exist in measurable quantities in the B. napus xylem sap. This was particularly the case with cysteine, as control plants routinely exhibited concentrations of around 800 nM. Further, the xylem sap cysteine concentrations showed a strong response to Cd exposure, which was clearly evident by 72 h after initial introduction of Cd into the trough system. Temporal responses in concentration, at least for the 1 and 5 μM Cd treatments, appeared to follow the exponential model over the time period that was studied. However, this model would likely be unsatisfactory over longer time periods because cysteine concentration increases could not be maintained indefinitely. Additionally, xylem loading of cysteine or other thiols in response to Cd is mechanistically complex, involving upregulation of synthesis pathways and increased xylem loading through membrane-bound transport proteins; these types of complexities are not captured by our model. If cysteine concentrations were related to the xylem sap Cd concentration, cysteine levels would likely have shown a plateau over a longer time period, similar to the rapid rise followed by a plateau in the xylem sap Cd levels that has been previously shown in A. halleri (Ueno et al., 2008). For the 0.5 μM Cd experiments, though, additional time points may have been needed to get a better fit to the cysteine data.
PC2 was also detected in the xylem sap of B. napus. Unlike cysteine, PC2 results did show a response to Cd by 24 h at all of the tested Cd concentrations. In this case, PC2 was not found in any of the control plants across all time periods, which agrees with the current understanding of how PC synthase operates and is activated in nonhyperaccumulating plants (Clemens, 2006a). Although some have postulated a role for PCs in metal homeostasis (Rauser, 1995), our results gave no evidence for such a function as PC2 was not detected in the xylem sap of plants that were not exposed to Cd. Alternatively, it is possible that our detection method was not sensitive enough to detect if PC2 was involved in metal homeostasis or there may be homeostatic functions that do not involve PC2 presence in the xylem sap. Although all Cd treatments were effective in causing a detectable PC2 response in the xylem sap, a particularly sharp increase in PC2 levels was observed at the highest Cd concentration. The physiological significance, if any, of this PC2 concentration spike was unknown, although the higher Cd concentration represented a severe challenge to B. napus as evidenced by leaf chlorosis. PC2 temporal trends were well described by the exponential model at all of the tested Cd concentrations. However, the model again would likely be unsatisfactory over longer time periods for the same reasons that were previously discussed for cysteine. In this case, the model parameter C1 predicted PC2 concentrations between 19 and 75 nM at t = 0, which was close to our limit of detection of 50 nM. We were unable to detect PC2 in the xylem sap of control plants, which may have indicated that the model did not accurately describe PC2 data at t = 0. Alternatively, PC2 may have been present in control plants, but levels were below our detection limit. Instrumentation with higher sensitivity will be required to verify if PCs are present in the xylem sap of unexposed control plants.
Therefore, the PC2 results presented here provide direct evidence that PCs are capable of long-distance transport in B. napus via the xylem sap. However, the presence of PC2 does not mean that it was necessarily complexed to Cd. Although PCs have favorable thermodynamics for binding Cd (Chekmeneva et al., 2008), other ligands may also bind Cd (Callahan et al., 2006; Clemens, 2006b). The presence of PC2 in the xylem sap is in agreement with the A. thaliana PC transport studies that have shown PC transport in both the shoot-to-root and root-to-shoot directions (Gong et al., 2003; Chen et al., 2006). These results also further validate that direct measurement of PCs is possible in the xylem sap, as reported by Wei et al. (2007) on B. juncea, a relative of B. napus. However, some differences between our work and Wei et al. (2007) should be noted. First, we did not attempt to conduct a metal speciation study as it seemed unlikely that it was possible to avoid significant disturbance to the native xylem sap chemical equilibria using our sampling and detection methods. Further, our detection method relied upon labeling thiols with mBrB followed by separation on a C16 column and use of a fluorescence detector, rather than size-exclusion chromatography followed by a UV detector. Wei et al. (2007) found evidence for very few PCs in the xylem sap of B. juncea that was exposed to 10 μM Cd and concluded that 1% of the Cd was associated with PCs under this exposure. The work presented here detected PC2 at Cd exposures down to 0.5 μM. However, our Cd exposure times ranged from 24 to 72 h prior to collection, whereas a 10-h exposure period was used by Wei et al. (2007). As our work shows that the amount of thiols detected increased with time, it is possible that the shorter sampling time may have played a role in their results. Also, the highest Cd concentration that was used here was 5 μM, whereas Wei et al. (2007) used a maximum Cd exposure of 50 μM to induce a Cd-stressed condition that resulted in PC detection. Thus, our method, although not capable of resolving the speciation of Cd, may have been more sensitive for thiols. Despite these differences, however, both studies detected cysteine and PCs in the xylem sap. More work remains, though, in trying to determine the exact role of these compounds in long-distance Cd transport, which may be both time and concentration dependent.
More recently, it was shown that cysteine, PC2, PC3, GSH, and GSH-related peptides such as γ-glutamyl-cysteinyl-glutamine and hydroxymethyl-GSH were transported in the phloem of B. napus (Mendoza-Cózatl et al., 2008). In this case, 9-week-old B. napus plants were exposed to 75 μM Cd for up to 2 weeks, and it was shown that phloem levels of γ-EC, PCs, GSH-like, and PC-like thiols all increased in a time-dependent manner, whereas GSH and cysteine levels remained unchanged from controls. These authors also examined the xylem sap of B. napus and found measurable levels of GSH and cysteine, the latter of which appeared to show an increase in the xylem sap concentration over time in response to Cd. However, in contrast to our results, Mendoza-Cózatl et al. (2008) did not find significant quantities of PCs in the xylem sap of B. napus. The reason(s) for this difference is unclear. In both cases, a hydroponics system was used to grow and expose B. napus plants to Cd, although the Cd concentrations used in our study were much lower (0.5–5 μM versus 75 μM). Additionally, the plants used here were younger (4 weeks old rather than 9 weeks); therefore, age or Cd exposure levels may partly contribute to these differences. Also worth noting are the shorter Cd exposure times used in our study, which ranged between 24 and 72 h prior to plant detopping and xylem sap collection. Thus, our study focused on shorter-term responses to low-level Cd challenges, whereas Mendoza-Cózatl et al. (2008) focused on longer-term exposures to a much higher Cd concentration, which may have caused differences in plant responses. Methodological differences in the length of xylem sap collection times may have also contributed to these different findings. Our collection method was similar to the one used in a study of xylem transport of arsenite (Su et al., 2008), in that total collection periods were 1–2 h immediately after plant detopping and surface washing.
The xylem sap analysis also indicated the presence of an unknown thiol compound in chromatograms that showed an increasing response to Cd over time (Fig. 4). This compound always eluted off of the HPLC column at a time very close to that of the GSH standard and was thus termed GSH-like. However, because the difference in elution times was consistently 0.2–0.3 min, the unknown compound did not appear to be chemically identical to the GSH standard. Plants are known to contain GSH variants, called homo-GSHs, which have different C-terminal amino acids and result from different specificities of GSH synthetase or possibly postsynthesis modifications of GSH (Skipsey et al., 2005). These GSH variants subsequently give rise to homo-PCs (Grill et al., 1986), and it has been shown that B. napus does contain variants of GSH, which result in altered PCs (Mendoza-Cózatl et al., 2008). Although the xylem sap PC2 occasionally deviated slightly from the elution time of the PC2 standard, the deviation was not consistent across samples and was within the usual variation observed in elution times (<0.15 min.). However, an earlier work (Grill et al., 1986) reported that homo-GSH (and PCs derived from h-GSH) had only slightly altered retention times on HPLC systems, and our HPLC method may not have been able to distinguish between these variants. Statistical analysis of this GSH-like compound revealed meaningful differences only in 72-h plants when compared with controls. The fact that these differences were found only in the 72-h group was likely due to the higher variability seen in the GSH-like data, especially in the 48-h group, which subsequently weakened the power of the ANOVA model to detect meaningful differences in concentration among the various Cd treatments.
Taken together, the work presented here provides direct evidence for the presence of cysteine, PC2, and other thiol-containing compounds in the xylem sap of B. napus. Additionally, the xylem sap thiol responses to Cd generally took 48–72 h to reach levels that were significantly different from controls. The precise role of thiols in long-distance Cd transport remains unclear, although their presence in the xylem sap supports the hypothesis that, under the right conditions, thiols may be involved in long-distance heavy metal transport.
Footnotes
Acknowledgment
The authors thank Dr. D.W. Auld, University of Idaho, for the supply of B. napus seeds.
Author Disclosure Statement
No competing financial interests exist.