
Abstract
Abstract
Textile effluents (TEs) without correct treatment cause high environmental impact because they display several problems mainly due to toxicity and recalcitrance of dyestuffs. The aim of this work was to examine the performance of Pseudomonas putida mt-2 in treating TE. P. putida was able to completely decolorize the studied effluent after 8 h of incubation under agitation in an aerobic bioreactor. Genotoxicity of effluent, before and after biodegradation with P. putida mt-2, was evaluated in vitro using the SOS chromotest and in vivo in mouse bone marrow, by assessing the percentage of cells bearing different chromosome aberrations compared with untreated mice. Results indicated that TE showed a significant ability to induce DNA damage, which was evaluated in both SOS (IF = 3.52) and chromosome aberration assay systems (23.5%). In addition, TE reduced strongly the acetylcholinesterse (60%) and butyrylcholinesterase (51%) activities and induces a remarkable lipid peroxidation effect (increased markedly by approximately fivefold). This toxicity was imputed to the presence of dye compounds of TE. However, toxicity in TE was significantly reduced after 8 h of aerobic incubation with P. putida mt-2 strain. The present study demonstrates that P. putida mt-2, incubated under aerobic conditions, has a metabolism that enables it to degrade TE and, especially, to detoxify the effluent mixtures.
Introduction
TEs are one of the most difficult-to-treat wastewaters on account of their considerable amount of suspended solids as well as the massive presence of toxic substances such as additives, detergents, surfactants, and particularly dyes (Skolar and Le Marechal, 1998). The presence of even trace concentrations of dyes in effluent is highly visible and undesirable. The dye-related industries, in particular, are known to produce consistently genotoxic effluents that have been shown by many scientists to be potent relative to other industrial discharges. A number of studies have demonstrated mutagenic activity in effluents from textile and dye-related industries (Sanchez et al., 1988; Wells et al., 1994). Azo dyes are extensively used in textile dyeing because of their superior fastness to the applied fabric, high photolytic stability, and their resistance to microbial degradation (Crini, 2008; Ferrero, 2007).
Their metabolism and toxicology have been the focus of considerable research because azo dyes are mainly metabolized at the intestinal wall and liver, producing free aromatic amines that are potentially carcinogenic and mutagenic (Chung et al., 1981; Manning et al., 1985; Ben Mansour et al., 2007). In fact, some studies reported mutagenicity of azo dyes (Chung et al., 1981; Umbuzeiro et al., 2005) and their potential to cause some types of cancer (Miller and Miller, 1961; Alves de Lima et al., 2007). According to Medvedev et al. (1988) and Percy et al. (1989), some azo dyes have been related to bladder cancer in humans; splenic sarcomas, hepatocarcinomas, and nuclear anomalies in laboratory animals; and chromosomal aberrations in mammalian cells. Myslak et al. (1991) have observed that German painters developed bladder cancer after long-time exposure to azo dyes.
It is well known that some azo dyes are both mutagenic and carcinogenic, for example, 6-dimethylaminophenylazobenzothiazole is a strong mutagen in the Salmonella typhimurium TA98 bacterial tester strain in the presence of an induced rat liver S9 mix (TA98+S9) and a potent liver carcinogen in rodents (Brown and Devito, 1993). In stark contrast, other azo dyes are neither mutagenic nor carcinogenic under quite similar conditions, for example, 40-phenyl-4-dimethylaminoazobenzene (Ashby et al., 1983). There are also some azo dyes that have been shown to be mutagenic but not carcinogenic, for example, 5-dimethylaminophenyl-azoindoline is a strong mutagen in TA98+S9; however, it does not appear to be a rodent carcinogen (Brown and Devito, 1993). Cytotoxicity of typical azo dyes may be relatively low, but the toxicity of related aromatic amine intermediates is very likely to be significantly high because of their carcinogenicity and/or mutagenicity (Ben Mansour et al., 2007, 2009b, 2009c).
Several mechanisms for the carcinogenicity of azo dyes have been identified in the literature (Brown and Devito, 1993). These mechanisms, which may be compound specific, generally incorporate some form of metabolic activation of the dye to reactive electrophilic intermediates that covalently bind to biological macromolecules.
Based on these observations, release of azo dyes into the environment from the effluents of dye-utilizing industries has become a major concern in wastewater treatment. Among the various types of biomass, Pseudomonas putida mt-2 was able to degrade some dyes, mostly found in textiles industries (Ben Mansour et al., 2007, 2009a, 2009b, 2009c, 2009d, 2010). In this study, we have opted to assess the ability of P. putida mt-2 to decolorize TE and to detoxify it.
In vitro and in vivo genotoxicity were evaluated for P. putida-treated and untreated TEs by using the SOS chromotest (a bacterial assay involving a set of functions known as the SOS responses induced by DNA-damaging agents) and by observing chromosome aberrations frequency. In addition, butyrylcholinesterase and acetylcholinesterase activities were also evaluated in mice bone marrow treated with TE before and after biodegradation in the presence of P. putida.
Materials and Methods
Chemicals
Butyrylthiocholine iodide, acetylcholine iodide, and 5,5′-dithiobis [2-nitrobenzoic acid] (DTNB) were purchased from Quimica Clinica Aplicada S.A. Vinblastin (Gedeon Richter Ltd.). Giemsa was obtained from Fluka (France). Methanol and acetic acid were obtained from Prolabo (France). The N-(1-naphtyl) ethlenediaminedihydrochloride, zearalenone o-Nitrophenyl-
Animals
The mice used for the experiments were female white BALB/C, which were of similar age and weight (20–25 g). Animals were kept for 1 week before the experiments for acclimatization and were maintained with food (conventional chow) and water ad libitum.
Characteristics of the textile wastewater
The original textile wastewater used in the present study was obtained in 2008 from an ennoblement industry located in Sousse, Tunisia. The effluent contained five azo dyes: Yellow GR (≈100 mg/L), Red FE3B (≈50 mg/L), Blue BRR (≈50 mg/L), Blue GLL (75 mg/L), and Brown BL (75 mg/L); dispersant (sarabide 10 g/L); two fixative agents (sodium sulphate 20 g/L and Rewin ACP 5 g/L); and potassium nitrate (1 g/L).
The effluent was sterilized by autoclaving at 120°C for 15 min prior to the biodegradation and toxicity studies.
Microorganisms and culture procedures
P. putida mt-2 (DSM 3931) used for the decolorization assays was obtained from Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH and was grown at 30°C in 250-mL flasks containing 50 mL of medium, under rotary shaking incubation at 200 rpm. The growth medium contained yeast extract (10 g/L), peptone (3 g/L), and glucose (5 g/L). Fresh textile wastewater (5 mL), sterilized with a 0.45-μm filter (Nalgene Labware), were then added to the culture medium to induce the bacterium metabolization system.
After cultivation of P. putida mt-2 on an enzymatic inductive medium (nutrient broth supplemented with 10% of textile wastewater), the exponential phase culture (∼1.5 g/L of dry cells) was centrifuged (1,700 g for 10 min) to harvest cells, redissolved in 50 mL textile wastewater, and transferred into a 5-L bioreactor containing 2.450 mL of fresh textile wastewater supplemented with 5 mL of glucose (final concentration: 1 g/L). Then, color removal of textile wastewater was performed. Biodegradation test were conducted in the bioreactor at 30°C under oxygenated conditions assured by agitation (200 rpm) and air injection.
Samples (5 mL) of culture decolorization medium were taken every 2 h from the specific exit situated in the bioreactor and centrifuged (1,700 g, 10 min), and the absorbance of the supernatant (filtered at 0.45 μm; NalgeneTM Labware Neerijse) was measured at 520 nm using Pharmacia Biotech Novaspec II spectrophotometer.
The percentage of decolorization was calculated as follows:
where DOi is the absorbance before incubation with P. putida and DOf is the absorbance after incubation with P. putida.
The cell dry weight was estimated by recording absorbance at 660 nm (A660) and according to the following relation: Cell dry weight = 0.61 × A660 (Ben Mansour et al., 2007).
All decolorization experiments were performed in three sets. Abiotic (without microorganism) controls were always included.
After incubation of P. putida mt-2 for 8 h under aerobic condition, the decolorization medium was centrifuged (3,500 rpm, 10 min, 4°C). The supernatants were filtered at 0.45 μm and were directly used for the toxicity assays.
Toxicity assessment
In vitro genotoxicity. Escherichia coli
PQ37 described by Quillardet and Hofnung (1985) was used to perform the SOS chromotest genotoxicity assay according to the method described in previous works (Quillardet and Hofnung, 1985; Ben Mansour et al., 2007). Only one dose of each treated (obtained after incubation during 8 h in the presence of P. putida) and untreated TE (20 mL/assay).
The induction of DNA damages was evaluated using an induction factor (IF) calculated as the ratio of Rc/R0, where Rc is equal to β-gal activity/AP activity determined at concentration c and R0 is equal to β-gal activity/AP activity in the absence of the effluent. Aflatoxin and nifuroxazide were used as positive controls, respectively, in the presence and absence of the S-9 mix metabolization system.
In vivo toxicity
Animal treatments
Animals were randomly divided into four groups:
Animals given a single dose (5 mL/kg bw) of culture medium run in the presence of P. putida mt-2 and without TE (negative control group). Animals given a single dose of Zearalenone (4 mg/kg) constitute the positive control group. Animals given a single dose of treated textile wastewater, administrated intraperitoneally (5 mL/kg bw). Animals given a single dose of untreated textile wastewater, administrated intraperitoneally (5 mL/kg bw).
It is of note that our results clearly showed that the selected dose of textile wastewater and its biodegradation derivatives administrated alone to animals did not have any toxic effect (mortality, body weight, feed intake, and size and shape of liver and kidney).
Twenty-four hours before sacrifice, animals were given 500 μL of yeast extract/glucose (at final concentrations of 100 and 200 mg/mL, respectively) mixture to accelerate mitosis of bone marrow cells. Vinblastin (200 μL; at a final concentration of 250 μg/mL) was injected into the animals at 45 min before sacrifice, to block dividing cells in metaphase. Finally, the animals were sacrificed by cervical dislocation.
In vivo chromosome aberration assay
Bone marrow cells were obtained according to the technique of Yosida and Amano (1965). Briefly, femur and tibia were removed immediately after animal sacrifice and bone marrow was flushed out with KCl solution (0.075 M, preheated to 37°C) using a syringe. The bone marrow cell suspension was incubated for 20 min at 37°C and centrifuged at 3,500 rpm for 10 min. The pellet was washed three times with acetic acid/methanol (1:3, v/v) and then resuspended in 1 mL of the same solution and used for chromosome preparation. Chromosomes were prepared as reported by Evans et al. (1999) and Ben Mansour et al. (2010) and stained with Giemsa solution freshly prepared (4%, v/v) in phosphate buffer (0.15 M, pH 7.2). The slides were examined under 100 × magnifications using an optical microscope (Carl Zeiss). Three hundred well-spread metaphases per group were analyzed for abnormalities. Metaphases with chromosome breaks, gaps, ring, and centric fusion (robertsonian translocation) were recorded and expressed as percentage of total metaphases per group.
Serum preparation for studying butyryl and acetylcholinesterase activity inhibition and lipid peroxidation effect
Blood was collected in heparin glass tubes (plastique Gosselin 59522); red blood cells were eliminated by centrifugation at 1,800 rpm for 10 min, and the plasma (supernatant) was then recovered. Mouse plasma was used immediately for studying acetylcholinesterase, butyrylcholinesterase (BuChE), and lipid peroxidation activities or conserved at 2°C–8°C (stable for 7 days).
Evaluation of acetylcholinesterase and butyrylcholinesterase activities
BuChE and AcChE inhibiting activities were measured by the spectrophotometric method previously reported by Ellman et al. (1961) and adapted according to our experimental conditions. Before use, plasma was diluted (1/200) with 100 mM of phosphate buffer (pH = 8). Acetylcholine and butyrylthiocholine were used as substrates to assay BuChE and AcChE activities, respectively. To calculate cholinesterase activities, the following procedure was employed: 1 mL of 100 mM phosphate buffer (pH = 8) containing 0.26 mM of DTNB and 100 μL of plasma was placed in a microcuvette, which was incubated for 15 min at 30°C. The reaction (at 30°C) was started by the addition of acetylcholine (50 μL) or butyrylcholine and monitored by the formation of yellow 5-thio-2-nitrobenzoate anions resulting from the reaction of DTNB with the thiocholine released by the enzymatic hydrolysis of substrate. Absorbance was measured using an M350 double Beam UV-VIS spectrophotometer Camespec (TovaTech) at 405 nm, and the reading was repeated for 10 min at intervals of 30 s to verify the linearity of the reaction.
The enzymatic activity was calculated according to the following relation:
The inhibition percentages (%) of AcChE or BuChE activities were calculated as follows: (E − S)/E × 100, where E is the activity of the enzyme without test compound and S is the activity of enzyme in the presence of the test compound.
Evaluation of the lipid peroxidation effect of TE, using the thiobarbituric acid reactive species assay
The well-known method termed thiobarbituric acid reactive species (TBARS) assay concerns the spectrophotometric measurement of the pink pigment produced through reaction of thiobarbituric acid with malondialdehyde (MDA) and other secondary lipid peroxidation products. TBARS were determined according to Ohkowa et al. (1979). In this method, 0.4 mL plasma from each mouse plasma was mixed with 1.5 mL of 20% acetic acid and 1.5 mL of 0.8% thiobarbituric acid. The mixture was brought to a final volume of 4.0 mL with distilled water and heated to 85°C for 120 min. After cooling on ice for 10 min, 5.0 mL of a mixture of n-butanol and pyridine (15:1, v/v) was added to each sample, and the mixture was shaken vigorously. After centrifugation (3,000 rpm, 20°C, 10 min), the supernatant fraction was isolated and the absorbance was measured at 532 nm. Induction of lipid peroxidation was expressed as equivalent of MDA. Data were reported as mean ± SD for triplicate determinations.
Statistical analysis
Data are expressed as mean ± standard deviation from three replicates. The statistical analysis was performed with STATISTICA edition 99 France. Duncan test was used to compare tested compounds versus control. Difference was considered significant when p < 0.05.
Results
Biodegradation
The potential of P. putida mt-2 to decolorize textile wastewater was evaluated in a bioreactor under oxygenated condition. The kinetics of biodegradation plotted as increasing percentage of decolorization of effluent versus time is shown in Fig. 1. It shows that effluent coloration was totally removed after 8 h. The rate of decolorization was significantly lower in the first 2 h, showing that the presence of glucose limited the azo dye biodegradation and allowed strain growth.
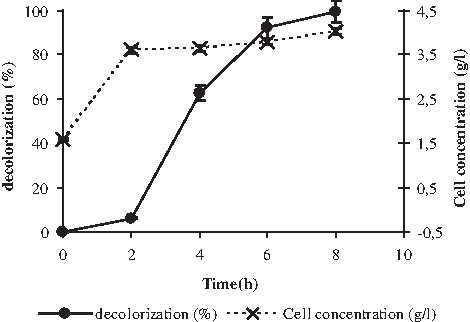
Decolorization of textile wastewater by Pseudomonas putida mt-2 in an aerobic bioreactor.
The curve biomass versus time (Fig. 1) reached a plateau after 2 h, with a final biomass of 4 g/L.
Toxicity
In vitro genotoxicity
In experiments, prior to the genotoxic study, it was ascertained that the amount (20 μL) of diluted TE added to the indicator bacterium (E. coli PQ37) does not influence its viability. No genotoxicity was shown by the TE before and after biodegradation without S-9 metabolization system because the induction factor (IF) determined by the SOS chromotest, in the presence of various concentrations of the compounds, ranged between 0.98 and 1.37. According to Kevekords et al. (1999), a compound is classified as “not genotoxic” if the IF is less than 1.5, as “marginally genotoxic” if it is between 1.5 and 2, and as “genotoxic” when it is more than 2.
When samples resulting from untreated TE were tested in the presence of S-9 (Table 1), IF exceeds 3.52, revealing a significant genotoxicity, whereas treated TE tested in the presence of S9 showed an IF of 1.73, indicating very weak genotoxic effect.
2-Aminofluorene and aflatoxine B1 are the positive controls used in the absence or presence of the metabolic activation system S-9, respectively.
p < 0.01.
TE, textile effluent.
In vivo toxicity
Chromosome aberrations
The volumes of TE, before and after biodegradation, injected into animals were chosen on purpose to obtain a compromise between chromosome damages and a sufficient number of scorable metaphases and limited lethality. Only structural aberrations induced by the different treatments were enumerated in the present study, with special emphasis on gaps, rings, breaks, and centric fusions. All of these types of structural abnormalities and their frequencies for both the control and treated groups are presented in Table 2. Numerical chromosome abnormalities such as polyploidy and aneuploidy were not evaluated.
Data are expressed as mean ± standard deviation.
p < 0.01;
p < 0.001.
Chromosome aberrations, whatever the type, significantly increased in mice treated with TE (23.5%) when compared with the negative control group (6%). Centric fusions represent the majority of chromosome abnormalities (about 10.5%), whereas gaps are the least frequent lesions, representing only 3.2%.
However, the ability to induce DNA damage decreased (6.5% of chromosome aberrations) when TE obtained after 8 h of aerobic bioreactor incubation with P. putida mt-2 was tested (Table 2).
Cholinesterase inhibition
Plasma AcChE and BuChE activities were 2,094 and 2,251 UI, respectively, in the bone marrow of male mouse treated with 100 μL of culture medium run in the presence of P. putida mt-2 and without TE (negative control). Figure 2 shows that the level of both AcChE and BuChE activities decreased significantly, thus indicating a toxic effect of the untreated TE when evaluated in the presence of intact TE. Inhibition percentages decreased, reaching 60% and 51%, respectively, when compared with the negative control.
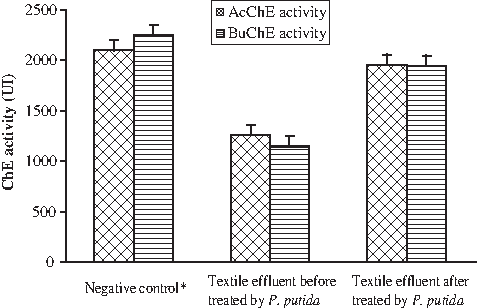
Plasma acetyl and butyrylcholinesterase activities in the bone marrow of mouse treated with textile effluent before and after biodegradation with P. putida. *Negative control: Animals given a single dose of culture medium run in the presence of P. putida mt-2 and without textile effluent.
However, a weak inhibition of these two enzyme activities was observed in the TE after treatment with P. putida. In fact, enzymatic activity values over 1,956 and 1,943 UI for AcChE and BuChE, respectively, correspond to percentage decreases of 6.5% and 13.61%, respectively.
Lipid peroxidation effect
After intraperitoneal administration of fresh TE to animals, the TBARS complex formation in serum increased markedly by approximately fivefold (corresponding to 40 nM) compared with the negative control (8 nM). However, less TABRS complex formation was observed, when animals were treated with the biodegradation extract of TE. In fact, the obtained MDA equivalent value (13 nM) was comparable with that of the control (8 nM).
Discussion
The textile industry plays an important role in Tunisia economy as well as in our daily life, but at the same time it consumes large quantities of water and generates huge amounts of polluted wastewaters (200 × 106 m3/year).
P. putida mt-2 exhibited a high ability to decolorize the TE collected from a Tunisian ennoblement industry. This result is very important as far as the conventional physical–chemical treatment of the tested effluent showed a weak efficiency, which could be ascribed to the considerable amount of suspended solids. These particles (such as dyes, sodium chloride, and potassium nitrate) are generally known to inhibit the growth and biodegradation power of bacteria (Yu et al., 2001; Tan et al., 2009). However, P. putida was found to be able to grow in this effluent and to utilize these particles as sole carbon source. The decolorization of the tested TE by P. putida could be explicated by the expression of the azoreduction system by this bacterium (Ben Mansour et al., 2007, 2009a, 2009b, 2009c, 2009d, 2009e), which reduces the contents of azo dyes in this effluent.
TE has been the subject of several studies that, however, essentially focused on the determination of chemical oxygen demand and biological demand oxygen to evaluate the cleanup degree. For our part, we choose to evaluate cleanup by assessing the potential toxicity of TE before and after biodegradation in the presence of P. putida mt-2. The SOS chromotest using prokaryotic cells indicated that TE exhibited no genotoxic response in the absence of S-9 metabolization system, indicating that the effluent constituents were not directly genotoxic, whereas TE exhibited a strong genotoxicity when tested in the presence of S9 system. This genotoxicity was confirmed by an in vivo toxicity test. To this end, we evaluated the effect of a single injection of TE on mouse bone marrow, by monitoring its effect on chromosome aberrations induction.
The results obtained with SOS chromotest were in accordance with those obtained with the chromosome aberration assay system. In fact, based on the statistical analysis, a significant number of some chromosome aberration types were induced by untreated TE. It has been established that centric fusions (robertsonian translocations) are detected relatively at high frequencies compared with other types of chromosome abnormalities; this may be due to the fact that almost all mouse chromosomes are acrocentric. These types of chromosomes have the exceptional ability to merge with each other. Only structural chromosome aberrations were investigated, as very little polyploidy or aneuploidy cases were detected. This specific aspect needs further investigation. In this study, the chromosome aberration test, recommended by regulatory authorities for the assessment of genotoxicity and mutagenicity of many chemicals and natural compounds, has provided positive data (Carita and Marin-Morales, 2008). These indirect genotoxic effects related to some of tested compounds can be explained by several mechanisms: generally, intact molecules undergo a metabolization by hepatic cytochrome P450, leading to the formation of epoxides and/or reactive oxygen species responsible of DNA damages (Ben Mansour et al., 2007). Thus, we evaluated the capacity of TE and its biodegradation metabolites to induce the oxidation of membrane lipids. In fact, lipid peroxidation can be initiated by the •OH radicals or by perferryl or ferryl derivatives generated in the reactions involving chelated iron, O2•−, and H2O2 (Natalia and Catalá, 2009). Curiously, the same compounds inducing the increase of chromosome aberration frequencies were revealed to be able to cause lipid peroxidation.
On the other hand, as cholinesterase has been commonly used as a biomarker for pollution evaluation and risk assessment (Roex et al., 2003; Tahara et al., 2005; Küçükkilinç and Özer, 2007) and a correlation has been established between mutagenicity/genotoxicity and cholinesterase inhibition activity (Williams et al., 1994; Eldeen et al., 2005; Ben Mansour et al., 2010), we have opted to assess the effects of the tested compounds on the AcChE and BuChE activities. Untreated TE revealed an effective inhibitory effect on both BuChE and AcChE activities. This was in accordance with the aforementioned genotoxic results obtained by in vitro and in vivo assay systems. However, the toxicity of TE decreased significantly after aerobic incubation in the presence of P. putida mt-2. Thus, we can attribute the observed toxicity in vitro as well as in vivo to the presence of colorant metabolites obtained in the presence of the metabolizing hepatic system, as this toxicity disappears in the P. putida-treated effluent. Biodegradation of colorant molecules by P. putida should give intermediates devoid of toxicity. Also, several studies have shown that the dye content is responsible for the TE genotoxicity and anticholinesterase and lipoperoxidation effects (Mathur et al., 2005; Ben Mansour et al., 2007, 2009a, 2009b, 2009c, 2009d, 2009e, 2010).
In a previous work, P. putida mt-2, incubated under good oxygenated conditions, was found to be able to oxidize totally azo dyes and their aromatic amine derivatives, which undergo metabolization pathways involving, amongst other activities, oxygenases (Ben Mansour et al., 2009c).
Footnotes
Author Disclosure Statement
No competing financial interests exist.